|
|
მთავარი - ბიბლიოთეკის შესახებ - ელ.რესურსები |



|
||||||||
|
|
|
მიკროევოლუციის საფუძვლები |
|
|
| საბიბლიოთეკო ჩანაწერი: |
| ავტორ(ებ)ი: შათირიშვილი ა., ჭუჭლაშვილი ი. |
| თემატური კატალოგი განათლება|სახელმძღვანელოები |
|
წყარო:
შათირიშვილი ა., ჭუჭულაშვილი ი. მიკროევოლუციის საფუძვლები : [სახელმძღვ. ბიოლ. სპეც. სტუდ. / რედ.: ი. ელიავა, თ. ლეჟავა] ; თსუ - თბ. : თბილ. უნ-ტის გამ-ბა, 2005 - 311გვ. : სურ. ; 20სმ. - - ბიბლიოგრ.: გვ. 298-300. - ISBN 99940-38-43-5 : სახელშეკრ. ფასი, 250ც.[MFN: 25912] UDC: 575.8(075.8) K 247.633/3 - საერთო ფონდი K 254.339/3 - საერთო ფონდი F 87.043/3 - ხელუხლებელი ფონდი P 339/2005 - ხელუხლებელი ფონდი |
|
|
1 წინასიტყვაობა |
▲ზევით დაბრუნება |
ბაკალავრიატში ფუნდამენტურ ბიოლოგიურ დისციპლინათა ათვისება ევოლუციური თეორიის შესწავლით სრულდება. იგი აჯამებს და განაზოგადებს ბიოლოგიის იმ კანონზომიერებებს, რომლებსაც სტუდენტები ცალკეული ბიოლოგიური დისციპლინის შესწავლის დროს დაეუფლნენ. ევოლუციურ თეორიაში ცენტრალური ადგილი მიკროევოლუციის კანონზომიერებათა განხილვას უკავია. მასში განიხილება ბიოლოგიის ისეთი საკვანძო საკითხები, როგორებიცაა სახეობის კატეგორია და მისი წარმოშობა, პოპულაციის სტრუქტურა, მიკროევოლუციის მამოძრავებელი ფაქტორები, ადაპტაციის ჩამოყალიბების მექანიზმები და ფორმები, პოპულაციური პოლიმორფიზმი.
ევოლუციურ გენეტიკასა და მოლეკულურ ბიოლოგიაში მოპოვებულმა წარმატებებმა მნიშვნელოვნად შეცვალა ჩვენი წარმოდგენები მიკროევოლუციის პროცესის შესახებ. გაღრმავდა და დაზუსტდა მიკროევოლუციის ზოგიერთი კარდინალური დებულება, ჩამოყალიბდა ევოლუციის ალტერნატიული თეორიები. სადღეისოდ მრავალი მიკროევოლუციური დებულება სრულიად განსხვავებულ ჭრილში განიხილება.
თბილისის სახელმწიფო უნივერსიტეტში ევოლუციის თეორიის სწავლებას დიდი ხნის ისტორია აქვს. 1936 წლიდან ფუნქციონირებდა გენეტიკისა და დარვინიზმის კათედრა, სადაც ერთ-ერთი მნიშვნელოვანი დისციპლინა ევოლუციური მოძღვრება იყო. ყოფილ საბჭოთა კავშირში 1948 წელს მომხდარი მოვლენები თბილისის სახელმწიფო უნივერსიტეტსაც შეეხო: დაიხურა გენეტიკისა და დარვინიზმის კათედრა, აიკრძალა გენეტიკისა და ევოლუციური თეორიის მეცნიერული კურსის სწავლება. მაგრამ პროფესორ-მასწავლებლები შენიღბულად მაინც ახერხებდნენ სტუდენტებისათვის ობიექტური ინფორმაციის მიწოდებას. 1936-67 წლებში ევოლუციური თეორიის კურსს უნივერსიტეტში ასწავლიდნენ პროფ. გ. პაპალაშვილი, დოც. ი. ახალაია, უფროსი მასწავლებლები ჯ. ბუაჩიძე და ქ. გოგინაშვილი 1967 წლიდან ევოლუციური თეორიის კურსის წაყვანა პროფ. ი. ჭუჭულაშვილს მიანდეს, რომელსაც იგი გარდაცვალებამდე (1992) უძღვებოდა. მან ძირეულად განაახლა სალექციო და სასემინარო კურსის თემატიკა. ი. ჭუჭულაშვილის თაოსნობით უნივერსიტეტში სალექციო კურსის წასაკითხად მოიწვიეს გამოჩენილი ევოლუციონისტები პროფ. ვორონცოვი, პროფ. ა. იუსუპოვი, აკადემიკოსები ი. ზახაროვი და ს. ინგე-ვეჩტომოვი. მანვე შეადგინა სახელმძღვანელო ევოლუციურ თეორიაში, რომლითაც სტუდენტები ეუფლებოდნენ ამ დისციპლინას. სამწუხაროდ, იმხანად ამ სახელმძღვანელოს დასტამბვა ვერ მოხერხდა.
წინამდებარე სახელმძღვანელოს საფუძვლად დაედო პროფ. ი. ჭუჭულაშვილის შრომა. იგი საფუძვლიანად გადამუშავდა უახლესი მონაცემების გათვალისწინებით და დაემატა ახალი თავებიც.
სახელმძღვანელოზე მუშაობის პერიოდში კონსულტაცია გაგვიწიეს პროფესორებმა: გ. ბედოშვილმა, რ. ჟორდანიამ, თ. ჯოხაძემ, დოცენტებმა: მ. შენგელიამ, ნ. ჯმუხაძემ, მ. კაჭარავამ, ნ. ჭელიძემ, მ. ჭურაძემ მ.კ. შ. ბარჯაძემ. ხელნაწერი ყურადღებით წაიკითხეს გენეტიკის კათედრის თანამშრომლებმა: თ. სადაგიშვილმა, მ. მენაბდემ, ასპირანტმა თ. ზარნაძემ. მათი მართებული შენიშვნები გათვალისწინებულ იქნა მუშაობის პროცესში. ინფორმაციის მოძიებაში დიდი დახმარება გაგვიწიეს პროფ. დ. ძიძიგურმა, პროფ. რ. გაგნიძემ და დოც. ე. მითაიშვილმა.
გვინდა აღვნიშნოთ სახელმძღვანელოს რედაქტორების - ე. წერეთლის, მეცნიერებათა აკადემიის წევრ-კორესპოდენტის პროფ. ი. ელიავას და გენეტიკის კათედრის გამგის, პროფ. თ. ლეჟავას, რეცენზენტების - ნეიროკიბერნეტიკის განყოფილების გამგის, პროფ. ს. ცაგარელის და დოც. ნ. ბარათაშვილის თანადგომა.
გულითად მადლობას ვუხდი ყველა მათგანს. მათ გარეშე ამ სახელმძღვანელოს არ ექნებოდა ის სახე, რომელიც ამჟამად აქვს. შენიშვნები, რომლებიც ხელს შეუწყობს სახელმძღვანელოს გაუმჯობესებას, მადლიერებით იქნება მიღებული.
პროფ. ა. შათირიშვილი
...დარწმუნებული ვარ, რომ ბუნებრივი
გადარჩევა იყო მეტად მნიშვნელოვანი,
მაგრამ არა ერთადერთი ფაქტორი ცვლილებისა.
ჩ. დარვინი
ევოლუციურ ბიოლოგიას სრულიად სამართლიანად
უწოდებენ უდიდესსა და ყოვლისმომცველ
ბიოლოგიურ თეორიას.
ე. მაირი
ჩ. დარვინის ევოლუციურმა თეორიამ
უდიდესი გადატრიალება მოახდინა
არა მხოლოდ ბიოლოგიაში, არამედ,
საზოგადოდ, საბუნებისმეტყველო მეცნიერებაში.
ი. შმალგაუზენი
|
|
2 შესავალი |
▲ზევით დაბრუნება |
ორგანული სამყაროს ისტორიულ განვითარებას იკვლევს მეცნიერება - ევოლუციური თეორია. იგი შეისწავლის ორგანული სამყაროს ევოლუციის მოვლენებს, მიზეზებს, გზებს, კანონზომიერებებსა და მართვის ხერხებს. ევოლუციური თეორია განაზოგადებს ბიოლოგიის ცალკეულ დარგში მიღებულ შედეგებს. იგი თანამედროვე ბიოლოგიის, სასოფლო-სამეურნეო და სამედიცინო პრაქტიკის უმნიშვნელოვანესი თეორიული საფუძველია.
ბიოლოგიური ევოლუცია არის ცოცხალი ბუნების შეუქცევადი და მიმართული ისტორიული განვითარება, რასაც თან ახლავს პოპულაციის გენეტიკური შემადგენლობის ცვლილება, ადაპტაციების ფორმირება, სახეობათა წარმოქმნა და ამოწყდომა, ეკოსისტემებისა და მთლიანად ბიოსფეროს გარდაქმნა.
ბიოლოგიური ევოლუცია ხასიათდება შემდეგი ძირითადი ნიშნებით:
1. ორგანული სამყაროს ევოლუცია* იგი, როგორც ყოველი განვითარება, უწყვეტ პროცესს წარმოადგენს. ყოველი სახეობა დროში წარმოდგენილია თაობათა უწყვეტი რიგით, თაობათა მონაცვლეობისას ხდება ახალი ელემენტების ჰარმონიული ჩაშენება ძველის წიაღში. განვითარება არაა ძველის განმეორება. ხდება თვისებრივად ახალი ფორმების წარმოქმნა - ამდენად იგი შემოქმედებით პროცესს წარმოადგენს.
2. ახალი ცვლილებების დაგროვება იწვევს ცოცხალ ფორმათა თვისებრივ გარდაქმნას. თაობათა უწყვეტ რიგში ერთ-ერთი თაობა იმდენად განსხვავებული შეიძლება აღმოჩნდეს დანარჩენებისაგან, რომ ამ უკანასკნელს ახალ სახეობად მიიჩნევენ. თაობათა უწყვეტ რიგში წარმოებს თვისებრივი ცვლილება. ახალი სახეობები ძველის არეალს იკავებენ. ხდება ფორმათა ცვლა დროში.
3. ევოლუციურ პროცესს შეგუებითი ხასიათი აქვს. სახეობების ევოლუცია მიმდინარეობს საარსებო პირობებთან შეგუების პროცესში. შეგუება (ადაპტაცია), პირველ რიგში, გულისხმობს ისტორიულ პროცესს, რომელიც იწვევს ისეთი ორგანული ფორმების განვითარებას, რომელთა აგებულება და ფუნქციები საარსებო პირობებთან სრულ შესაბამისობაშია, მეორე მხრივ, ადაპტაცია გულისხმობს ამ პროცესის შედეგსაც, ე.ი. ცოცხალი ფორმების მორფო-ფიზიოლოგიური ორგანიზაციის კონკრეტულ პირობებთან შესაბამისობის მოვლენასაც.
4. ევოლუციის საერთო მიმართულება იწვევს ცოცხალ ფორმათა ორგანიზაციის დონის ამაღლებას, რთული ორგანიზაციის ფორმები წარმოიშვნენ მარტივი ორგანიზაციის ფორმებიდან. იმავდროულად მიმდინარეობს მარტივი და რთული ორგანიზაციის დონის ფორმების თანაარსებობა. ორგანიზაციის დონის ამაღლება ევოლუციის საბოლოო შედეგია. ევოლუციის პროცესი ზოგიერთ შემთხვევაში ორგანიზაციის დონის ამაღლების გარეშეც ხორციელდება.
5. ევოლუციის პროცესი შეუქცევადია. ისტორიულად წარმოქმნილი ფორმები არ მეორდებიან. ევოლუცია უკუმიმართულებით არ მიმდინარეობს. მისი შედეგი ყოველთვის თვისებრივად ახალი ფორმების წარმოქმნაა. ამდენად, ევოლუციის შეუქცევადობა და განუმეორებლობა მისი ერთ-ერთი მნიშვნელოვანი თვისებაა.
6. სახეობათა მრავალფეროვნება ევოლუციის შედეგია. ცნობილია ცხოველთა 1,5 მილიონი, მცენარეთა 0,5 მილიონი, სოკოების 44 ათასი, ხოლო მიკრობების 4,5 ათასი სახეობა. ეს მრავალგვარობა სრულად კანონზომიერია და განისაზღვრება სახეობათა შორის ნათესაობის ხარისხითა და გარემოსთან შეგუების ფორმებით. ყველა ორგანიზმს საერთო ნიშნების მთელი წყება ახასიათებს. ასეთებია, მაგალითად, ცხოველური და მცენარეული უჯრედის ორგანიზაცია, გამრავლების პროცესები, მემკვიდრეობისა და ცვალებადობის მოვლენები, უჯრედის მიტოზური და მეიოზური გაყოფა, ქრომოსომათა ქცევა, ნივთიერებათა პლასტიკური და ენერგეტიკული ცვლის პროცესები და ა.შ. აღნიშნული ფაქტები ორგანული სამყაროს ერთიანი წარმოშობის მიმანიშნებელია. ამავე დროს ორგანული სამყაროს ერთიანი წარმოშობის ფაქტი მის მრავალფეროვნებას უნდა დაუკავშირდეს. მრავალგვარობა ორგანული ფორმების სხვადასხვა მიმართულებით ევოლუციის (დივერგენციის) შედეგია.
ზემოთ ჩამოთვლილ მოვლენებს მოეპოვებათ თავისი მიზეზები, რომელთაც ევოლუციის თეორია შეისწავლის. ევოლუციის მთავარი მიზეზი ბუნებრივი გადარჩევაა, რომელიც ჩ.დარვინმა აღმოაჩინა. ევოლუციის თეორია ორ მთავარ პრინციპს აქტუალიზმსა და ისტორიზმს ეფუძნება. ისტორიზმის პრინციპის თანახმად, თანამედროვე ორგანიზმები შეისწავლება იმ ცოდნის მომარჯვებით, რომელიც წარსულში მომხდარი ევოლუციის საფუძველზეა მიღებული. მის არსს გამოხატავენ თეზით - „წარსულის შესწავლა აწმყოს შემეცნების გასაღებია“. აქტუალიზმის პრინციპი კი მდგომარეობს აწმყოს ცოდნის ექსტრაპოლაციით წარსულში მომხდარი მოვლენების შესასწავლად. მის არსს გამოხატავენ თეზით: „აწმყოს შესწავლა წარსულის შემეცნების საფუძველია“.
|
|
3 §1. ევოლუციური თეორიის განვითარების მოკლე ისტორია |
▲ზევით დაბრუნება |
ევოლუციური თეორიის განვითარების საწყის ეტაპად მიჩნეულია პერიოდი, როდესაც გავრცელებული იყო სამყაროს ერთიანობისა და მისი განვითარების იდეა. ძველი სამყაროს (ჩინეთი, ინდოეთი, მესოპოტამია, ეგვიპტე, საბერძნეთი) მრავალი სწავლული და ფილოსოფოსი იზიარებდა ამ იდეას. ჯერ კიდევ ჩვ.წ.აღ-მდე I ათასწლეულის ბოლოს ჩინეთში გავრცელებული იყო მოძღვრება თანდათანობითი ცვლილების გზით ერთი ფორმის ორგანიზმთა მეორედ გარდაქმნის შესახებ. ასევე ჩვ.წ.აღ-მდე I ათასწლეულში ინდოეთში არსებობდა ფილოსოფიური სკოლები, რომლებიც იზიარებდნენ „პირველადი მატერიიდან“ მატერიალური (მათ შორის ორგანული) სამყაროს წარმოშობისა და განვითარების იდეას. აიურვედას უძველეს ტექსტებში გადმოცემულია შეხედულება მაიმუნიდან ადამიანის წარმოშობის შესახებ. სამყაროს ერთიანობისა და განვითარების იდეა სრულყოფილი სახით წარმოდგენილია ანტიკური ფილოსოფოსებისა და სწავლულთა (ჰერაკლიტე, ემპიდოკლე, დემოკრიტე, ლუკრეციუსი, არისტოტელე და სხვ.) შრომებში. მათ მიერ მოცემულია მახვილგონივრული მოსაზრებანი ორგანულ სამყაროზე. ამ ეტაპისათვის დამახასიათებელია ორგანული ბუნების შემეცნებისადმი ანალიტიკური მიდგომა. იგი ძირითადად წარმოადგენდა აზროვნების ნაყოფს და არ ემყარებოდა ცოცხალ ბუნებაზე უშუალო დაკვირვებას, მათ განზოგადებას. ამდენად ძველი სამყაროს სწავლულები არ შეიძლება მივიჩნიოთ თანამედროვე ევოლუციონისტების უშუალო წინამორბედებად.
ჟან ბატისტ ლამარკი
ფრანგი ბუნებისმეტყველი ჟან ბატისტ ლამარკი დაიბადა 1744 წლის 1 აგვისტოს ბაზანტენში (პიკარდიის პროვინცია), ღარიბი აზნაურის ოჯახში. მშობლებს უნდოდათ ის მღვდელი გამოსულიყო და კათოლიკურ სასწავლებელში მიაბარეს. 16 წლის ლამარკი ტოვებს სკოლას და მოხალისედ მოქმედ არმიაში მიდის (მიმდინარეობდა 7-წლიანი ომი). იქ მან დიდი სიმამაცე გამოიჩინა და ოფიცრის წოდება მიიღო. 1768 წლამდე ლამარკი სამხედრო სამსახურში იმყოფებოდა. მაგრამ ავადმყოფობის გამო ის იძულებული შეიქნა სამხედრო სამსახურისთვის თავი დაენებებინა. ლამარკი სწავლას აგრძელებს პარიზის უმაღლეს სამედიცინო სკოლაში. შემდეგ მუშაობას იწყებს სამეფო ბოტანიკურ ბაღში (1772-1776), რომელიც შემდგომში (1793) პარიზის საბუნებისმეტყველო მუზეუმად გადაკეთდა. შრომისმოყვარე და ნიჭიერმა ახალგაზრდა მეცნიერმა 1778 წელს დაწერა დიდი შრომა სამ ტომად - „საფრანგეთის ფლორა“. ამ წიგნმა ლამარკს დიდი სახელი მოუხვეჭა და პარიზის აკადემიის წევრად აირჩიეს. 1778 წლიდან იგი სათავეში უდგას ბოტანიკის კათედრას, 1793 წლიდან კი ზოოლოგიის კათედრას სთავაზობენ. 1801 წელს ის აქვეყნებს შრომას „უხერხემლო ცხოველთა სისტემა“, რომელშიც პირველადაა გამოთქმული ევოლუციური იდეები. 1809 წელს ლამარკმა დაბეჭდა ფუნდამენტური შრომა „ზოოლოგიის ფილოსოფია“, რომელშიც ჩამოყალიბებულია მისი ევოლუციური კონცეფცია. ლამარკმა შემოიღო ცხოველთა დაყოფა უხერხემლოებად და ხერხემლიანებად, რომელიც ამჟამადაც მყარადაა დამკვიდრებული ბიოლოგიაში. აღსანიშნავია, რომ ტერმინი „ბიოლოგია“, თანამედროვე გაგებით, პირველად ერთდროულად, მაგრამ ერთმანეთისაგან დამოუკიდებლად ლამარკმა და გერმანელმა ბუნებისმეტყველმა გ. ტრევირანუსმა (1802) იხმარეს. ლამარკს ეკუთვნის გეოლოგიური, ჰიდროლოგიური და მეტეოროლოგიური გამოკვლევებიც, რომლებშიც ის ეყრდნობა განვითარების იდეასა და ისტორიზმის პრინციპს. სიცოცხლის უკანასკნელ წლებში ლამარკი დაბრმავდა და უკიდურეს სიღარიბეში ჩავარდა. ყველასაგან მივიწყებული 1829 წლის 18 დეკემბერს გარდაიცვალა. |
XVII ს. დასაწყისიდან XIXს. შუა წლებამდე, სისტემატიკის, შედარებითი ანატომიის, ბიოგეოგრაფიის განვითარებამ ხელი შეუწყო ფაქტობრივი მასალის დაგროვებას. მომდევნო ეტაპზე ჩამოყალიბდა შეხედულებათა სისტემა - ტრანსფორმიზმი - მოძღვრება ორგანულ ფორმათა ცვალებადობისა და ერთი სახეობის მეორე სახეობად გარდაქმნის შესახებ. იგი შეიცავდა ევოლუციური იდეის ჩანასახს. ტრანსფორმიზმის იდეის მიმდევართა (რ.ჰუკი, ე.დარვინი, დ.დიდრო, ჟ.სენტ-ილერი, ი.გოეთე და სხვ.) შეხედულებები ზოგჯერ ანტიკური და შუასაუკუნეების სწავლულთა და ფილოსოფოსთა შეხედულებებს ეფუძნებოდა. ტრანსფორმისტები აღიარებდნენ ორგანიზმთა გარდაქმნას, მაგრამ სათანადო არგუმენტებით ვერ ასაბუთებდნენ. მართალია, მათ ვერ შექმნეს სრული ევოლუციური კონცეფცია, მაგრამ გამოიკვეთა ევოლუციური თეორიის ძირითად პრობლემათა სფერო. კერძოდ: ორგანიზმთა ევოლუციის არსი და მიზეზები, ორგანიზმთა მიზანშეწონილობისა და მრავალფეროვნების მიზეზები, ცალკეულ სახეობათა შორის მსგავსებისა და განსხვავების მიზეზები, პროგრესული ევოლუციის და განვითარების სხვადასხვა დონეზე მყოფ ორგანიზმთა ერთდროულად არსებობის მიზეზები, ცალკეულ სახეობათა ამოწყდომის მიზეზები. ამ პრობლემების არსში ჩაწვდომამ და მეცნიერული დონის შემდგომმა განვითარებამ მოამზადა ნიადაგი ახალი ეტაპისათვის. კერძოდ, ხელი შეუწყო ევოლუციური თეორიის პირველი კონცეფციების ჩამოყალიბებას.
პირველი სრული და თანმიმდევრული ევოლუციური თეორიის ფუძემდებელია ჟან ბატისტ ლამარკი (1744-1829), რომლის ძირითადი დებულებები დასრულებული სახით გადმოცემულია შრომაში „ზოოლოგიის ფილოსოფია“ (1809). ლამარკის ევოლუციური კონცეფცია მოიცავს ორგანული სამყაროს განვითარების მთელ პერიოდს სიცოცხლის წარმოშობიდან, ადამიანის წარმოშობის ჩათვლით. ლამარკის კონცეფციით ორგანიზმები ცვალებადია, სახეობა (ასევე სხვა სისტემატიკური კატეგორიები) პირობითია და თანდათანობით სხვა სახეობებად გარდაიქმნებიან. ლამარკის აზრით, უმარტივესი ორგანიზმები თვითჩასახვით არაცოცხალი მატერიიდან წარმოიქმნებიან. მისი თეორიის მიხედვით ორგანიზმები იცვლებიან და ვითარდებიან უმარტივესიდან ურთულესისაკენ, ნაკლებ სრულყოფილიდან უფრო სრულყოფილისაკენ ორგანიზაციის დონის ამაღლების - „გრადაციის“ გზით. ევოლუციის მამოძრავებელ ძალად ლამარკს მიაჩნდა თვით ორგანიზმებში არსებული სწრაფვა სრულყოფისაკენ“, განვითარების პროცესში მათი გარდაქმნის მიზეზად გარემოს პირობების უშუალო ზემოქმედებით გამოწვეულ ცვლილებებს თვლიდა. გარემო პირობები თავისი ზემოქმედებით ცვლის და არღვევს გრადაციის ბუნებრივ მსვლელობას. მცენარეებსა და დიფერენცირებულ ნერვულ სისტემას მოკლებულ ცხოველებზე გარემო უშუალოდ მოქმედებს და შეგუებით ცვლილებებს იწვევს. ნერვული სისტემის მქონე ცხოველებზე გარემოს ცვლილება ქცევებისა და ჩვევების შეცვლას იწვევს. ვარჯიშის შედეგად შესაბამისი ორგანო იცვლება საჭირო მიმართულებით (მაგ., ჟირაფის, გედის, ბატის გრძელი კისერი, წყლის ფრინველთა თითებს შორის საცურავი აპკი და სხვ.). უვარჯიშობით კი ორგანო სუსტდება, კნინდება და ქრება (მაგ., თხუნელას თვალი, სირაქლემას ფრთები, გველის კიდურების უქონლობა და სხვ.), (ე.წ. პირველი კანონი). ეს ცვლილებები მემკვიდრეობით გადაეცემა (ე.წ. მეორე კანონი). მოგვიანებით, გენეტიკური გამოკვლევებით დადგინდა, რომ გარემოს ზემოქმედებით გამოწვეული ორგანოთა ცვლილება მოდიფიკაციაა და შეძენილი ნიშან-თვისებები არ მემკვიდრეობს. ლამარკის კონცეფციით, ადამიანი რომელიღაც უმაღლესი მაიმუნიდან წარმოიშვა. იგი ანთროპოგენეზის ერთ-ერთ უმთავრეს ფაქტორად ზეამართულ სიარულზე გადასვლას და მეტყველებას მიიჩნევდა.
ამრიგად, ლამარკის თეორია სწორად ასახავს ევოლუციის პროცესის რეალურად არსებობას, მაგრამ ვერ იძლევა მისი მიზეზების მართებულ ახსნას. ვინაიდან ლამარკის ევოლუციური კონცეფცია სუსტად იყო არგუმენტირებული, თანამედროვეთა შორის აღიარება ვერ პოვა და იგი დავიწყებას მიეცა.
მეცნიერულად დასაბუთებული ევოლუციური კონცეფცია მხოლოდ XIXს. მეორე ნახევარში ჩამოყალიბდა. საკუთარი დაკვირვებების, თავის დროის მეცნიერული ცოდნისა და სელექციის მიღწევათა განზოგადების საფუძველზე ჩარლზ რობერტ დარვინმა (1809-1882) ჩამოაყალიბა ევოლუციური კონცეფცია. თეორია გადმოცემულია ტრილოგიაში: სახეობათა წარმოშობა ბუნებრივი გადარჩევის გზით, ანუ უკეთშეგუებული ჯიშების გადარჩენა სიცოცხლისათვის ბრძოლაში“ (1859); „შინაური ცხოველებისა და კულტურული მცენარეების შეცვლა“ (1868); „ადამიანის წარმოშობა და სქესობრივი გადარჩევა“ (1871).
ჩ. დარვინი „სახეობათა წარმოშობაზე“ დაახლოებით ოც წელს (1837-58) მუშაობდა. 1858 წ. 18 ივნისს ჩ. დარვინმა მიიღო ინგლისელი ზოოლოგის ალფრედ რასელ უოლესისაგან (1823-1913) სტატია, რომელშიც ევოლუციის იგივე კონცეფცია იყო გადმოცემული. ნაშრომში იდეა ნათლად იყო ჩამოყალიბებული, ხოლო შესრულებული იყო ნაჩქარევად. ავტორი სთხოვდა დარვინს სტატია გადაეცა გამოჩენილი გეოლოგისათვის, ჩ. ლაიელისათვის. აქვე უნდა აღინიშნოს, რომ ამ კონცეფციამდე ორივე ავტორი თომას რობერტ მალთუსის (1766-1834) ნაშრომმა „ხალხთმოსახლეობის ბუნებრივი კანონი“ (1822) მიიყვანა. დადგა პრიორიტეტის პირველაღმომჩენის საკითხი. დარვინს შეეძლო სტატიის გადაცემა საკუთარი ნაშრომის გამოქვეყნების შემდეგ, რაც მან არაკეთილსინდისიერად მიიჩნია და უოლესს თხოვნა შეუსრულა. ორივე ავტორმა თავიანთი თეორია 1858 წ. 1 ივნისს მოახსენეს ლონდონის ლინეს საზოგადოებას და დაიბეჭდა ამავე წელს საზოგადოების აგვისტოს შრომათა კრებულში. მოგვიანებით უოლესი წერდა, რომ ევოლუციის თეორიაზე დარვინი გაცილებით ადრე მუშაობდა და პრიორიტეტი მას ეკუთვნისო. მანვე შემოიტანა ევოლუციის თეორიის აღსანიშნავად ტერმინი „დარვინიზმი“. ხშირად ევოლუციის ამ კონცეფციას დარვინ-უოლესის თეორიას უწოდებენ.
ჩარლზ რობერტ დარვინი
ინგლისელი ბუნებისმეტყველი ჩარლზ რობერტ დარვინი დაიბადა 1809 წლის 12 თებერვალს, ქალაქ შრუზბერში. მამა იყო ცნობილი ექიმი, ხოლო პაპა - ერაზმ დარვინი - XVIII საუკუნის მოწინავე მეცნიერი, სახელგანთქმული პოლკოვნიკი, ექიმი, ნატურალისტი, ფილოსოფოსი და პოეტი.
დარვინმა დაწყებითი განათლება შრუზბერში, კერძო სკოლაში მიიღო. 1825 წელს მამის რჩევით, ის ედინბურგის უნივერსიტეტის სამედიცინო ფაკულტეტზე აგრძელებს სწავლას, სადაც მან ორი წელი დაჰყო. დარვინის მამა დარწმუნდა, რომ მისგან ექიმი არ გამოვიდოდა და ურჩია კემბრიჯში ღმრთისმეტყველებას დაუფლებოდა. დარვინმა წინადადება მიიღო. მას სურდა სოფლის მღვდელი გამოსულიყო, მაგრამ, ამასთანავე, ემუშავა ზოოლოგიაში როგორც მოყვარულ ნატურალისტს. 1831 წელს მან დაასრულა კემბრიჯის უნივერსიტეტის თეოლოგიის ფაკულტეტი და ბაკალავრის ხარისხი მიიღო.
უნივერსიტეტში ყოფნის დროს, დარვინი ძლიერ გაიტაცა ზოოლოგიამ და ბოტანიკამ. უნივერსიტეტის დამთავრებისთანავე დარვინმა გემ „ბიგლით“ დედამიწის გარშემო იმოგზაურა (1831-1836 წწ.). სამეცნიერო ექსპედიციისას ჩატარებული ბოტანიკური, ზოოლოგიური, პალეონტოლოგიური და გეოლოგიური დაკვირვებების საფუძველზე შეაგროვა მდიდარი საკოლექციო მასალა. 1842 წელს ის დასახლდა სოფელ დაუნში, სადაც სიცოცხლის ბოლომდე, ცხოვრობდა. 1842 წელს დარვინმა დაწერა 35 გვერდიანი (თავისთვისდა არა გამოსაქვეყნებლად) ევოლუციური კონცეფციის პირველი ნარკვევი. 1844 წელს ევოლუციური თეორია განავრცო და 230 გვერდიანი ნაშრომი შეადგინა. გავიდა შეუპოვარი შრომისა და ძიების კიდევ 14 წელი, ვიდრე დარვინი თავის იდეის გამოქვეყნებას გადაწყვეტდა. მეგობრების ჩ.ლაიელისა და ჯ.ჰუკერის მტკიცე მოთხოვნის შემდეგ, დარვინის შრომის მოკლე რეფერატი, ა.უოლესის სტატიასთან ერთად, 1858 წლის აგვისტოს შრომებში გამოქვეყნდა. 1859 წლის 24 ნოემბერს დაიბეჭდა დარვინის კაპიტალური ნაშრომი „სახეობათა წარმოშობა...“, რომელიც ბიოლოგიაში ახალი ერის დასაწყისს მოასწავებდა. წიგნის ყველა ეგზემპლარი იმავე დღეს გაიყიდა, რაც მოწმობდა პროგრესულად მოაზროვნე საზოგადოების დიდ ინტერესს ამ თხზულებისადმი.
გულმოდგინე მუშაობა, დიდი წინდახედულება, ფართო დიაპაზონი დარვინში შერწყმული იყო არაჩვეულებრივ თავმდაბლობასთან. მან მრავალრიცხოვანი შრომები დატოვა. ისინი ეხება ზოოლოგიის, ბოტანიკის, გეოლოგიისა და ნიადაგმცოდნეობის საკითხებს. დარვინის დიდი დამსახურებაა, რომ მან დაადგინა ევოლუციურის განვითარების კანონზომიერებები და სახეობათა ჩამოყალიბების პროცესი, აღმოაჩინა ევოლუციის მამოძრავებელი ძალები.
დარვინი გარდაიცვალა 1882 წლის 19 აპრილს. ის დიდი პატივით დაკრძალეს უესტმინსტერის სააბატოში - ინგლისის პანთეონში - ნიუტონის, ფარადეისა და სხვა დიდ მოღვაწეთა გვერდით. |
ალფრედ რასელ უოლესი
გამოჩენილი ბუნებისმეტყველი ალფრედ რასელ უოლესი დაიბადა 1823 წლის 8 იანვარს ასკიში (მონმუტშირის საგრაფო). დაწყებითი განათლება ჰერტფორდის დაწყებით სკოლაში მიიღო. 1844 წელს მსახურობს სკოლის მასწავლებლად ჯერ ლეიჩესტერში, ხოლო შემდეგ - უელსში. 1848-1852 წლებში გამოჩენილ ენტომოლოგ ბეიტსთან ერთად მდინარეების ამაზონისა და რიუ-ნეგრუს სანაპიროების ფლორა და ფაუნა გამოიკვლია.
1852 წელს ტროპიკული ციება შეეყარა და იძულებული გახდა ინგლისში დაბრუნებულიყო. შუა ზღვაში გემს ხანძარი გაუჩნდა და მრავალწლიანი შრომის შედეგად შეგროვებული ძვირფასი კოლექცია და ხელნაწერები მთლიანად განადგურდა.
1854-62 წლებში უოლესმა გამოიკვლია მალაის არქიპელაგის ფლორა და ფაუნა. შეაგროვა ზოოლოგიური და ბოტანიკური, ასევე გეოლოგიური კოლექციები. მან შეისწავლა არქიპელაგზე მცხოვრებ ხალხთა კრანიოლოგია. მოპოვებული მასალის საფუძველზე მან დაასაბუთა, რომ მალაის არქიპელაგზე გადის საზღვარი (ე.წ. „უოლესის ხაზი“), რომელიც ზოოგეოგრაფიულად ყოფს კუნძულ სულავესს არქიპელაგის სხვა კუნძულებისაგან.
უოლესმა დარვინისაგან დამოუკიდებლად ჩამოაყალიბა ევოლუციის კონცეფცია, კერძოდ, ბუნებრივი გადარჩევის კონცეპტი. 1858 წელს სტატია დაბეჭდა ლინეს საზოგადოების შრომებში, სადაც დარვინის რეფერატიც გამოქვეყნდა. მიუხედავად იდეათა მსგავსებისა, მან ბუნებრივი გადარჩევის თეორიის შექმნის პრიორიტეტი დარვინს მიანიჭა, ხოლო ევოლუციის შემსწავლელ ახალჩამოყალიბებულ ბიოლოგიის დარგს „დარვინიზმი“ უწოდა. მანვე ჩაუყარა საფუძველი ევოლუციურ ზოოგეოგრაფიას.
უოლესი გარდაიცვალა 1913 წლის 7 ნოემბერს ბროდსტოუნში (დორსეტშირის საგრაფო). |
ამ კონცეფციის თანახმად ევოლუციის ფაქტორებია მემკვიდრულობა, ცვალებადობა და ბუნებრივი გადარჩევა. მექანიზმი, რომელიც წინარე სახეობიდან ახალი სახეობების წარმოშობას აპირობებს, არის ბუნებრივი გადარჩევა. ბუნებრივი გადარჩევის პრინციპი სამ მთავარ დაკვირვებასა და ორ დასკვნაზეა დაფუძნებული.
დაკვირვება 1. დარვინმა შენიშნა, რომ ყოველ ინდივიდს დიდი რეპროდუქციული პოტენციალი გააჩნია. მათ გეომეტრიული პროგრესიით გამრავლების უნარი აქვთ.
დაკვირვება 2. ყოველ სახეობაში ინდივიდთა რაოდენობა თაობათა მანძილზე თითქმის უცვლელია. რეალურად არსებულ ინდივიდთა რაოდენობა არ შეესაბამება გამრავლების ინტენსიობას (ჩნდება მრავალი შთამომავალი, მაგრამ სქესობრივ მომწიფებას მხოლოდ ერთეულები აღწევენ).
დასკვნა 1. ვინაიდან მშობლები დიდი რაოდენობით იძლევიან შთამომავლობას, ხოლო ზრდასრულ ასაკს ერთეულები აღწევენ, მათ შორის წარმოებს კონკურენცია საარსებო რესურსებისათვის, ანუ მიმდინარეობს „არსებობისათვის ბრძოლა“.
დაკვირვება 3. დარვინმა შენიშნა, რომ ერთსა და იმავე სახეობაში ორი აბსოლუტურად იდენტური ინდივიდი არ გვხვდება. შეინიშნება ნაირგვარობა (პოლიმორფიზმი), სხვაგვარად, ცვალებადობა ცოცხალი ბუნების ერთ-ერთი თვისებაა. თუ განსხვავდებიან ზრდასრული ფორმები, იმავე მოვლენას უნდა ვხვდებოდეთ მოზარდებშიც.
დასკვნა 2. არსებობისათვის ბრძოლაში იმარჯვებენ ის ინდივიდები, რომელთაც მოეპოვებათ სასარგებლო ნიშან-თვისებები, რითაც უკეთ ეგუებიან გარემო პირობებს და ამიტომ, ისინი დიდი რაოდენობით ტოვებენ შთამომავლობას, ე.ი. მათ „რეპროდუქციული უპირატესობა“ აღენიშნებათ ნაკლებ შეგუებულებთან შედარებით. დროთა განმავლობაში ამ პროცესის მრავალჯერადი განმეორება საზიანო ვარიაციების ელიმინაციას, და სასარგებლო ვარიაციების გადარჩენას იწვევს. უკეთ შეგუებულთა გადარჩენა არსებობისათვის ბრძოლაში ბუნებრივი გადარჩევის საფუძველია.
დარვინმა დამაჯერებლად აჩვენა, რომ ბუნებრივი გადარჩევის მეშვეობით შესაძლებელია აიხსნას ორგანული სამყაროს ყველა მახასიათებელი, ტაქსონომიურ-იერარქიული სისტემიდან მოყოლებული, ფარშავანგის კაშკაშა შეფერილობის წარმოშობით დამთავრებული, როგორც რუდიმენტული ორგანოების არსებობა, ისე დედამიწაზე ორგანიზმთა განსახლების თავისებურებანი. დარვინის მეცნიერული მემკვიდრეობა მეტყველებს იმაზე, თუ როგორ ღრმად და მრავალმხრივ სწავლობდა იგი ბუნებას, რაოდენ დიდი იყო მისი სამეცნიერო ერუდიცია. ასევე განსაცვიფრებელია მისი თეორიული ნოვატორობა. დიდი წინააღმდეგობის მიუხედავად დარვინ-უოლესის კონცეფცია სწრაფად დამკვიდრდა მეცნიერებაში და საფუძვლად დაედო თანამედროვე ბიოლოგიას. დარვინის თეორია განუწყვეტლივ ვითარდება და მასში შეიძლება სამი პერიოდი გამოვყოთ: რომანტიკული, უარყოფისა და სინთეზური თეორიის (ნეოდარვინიზმი).
რომანტიკული პერიოდის (1859-1900) ბიოლოგიაში ჩამოყალიბდა ახალი მიმართულებები და დისციპლინები: ევოლუციური ემბრიოლოგია, ეკოლოგია, ევოლუციური პალეონტოლოგია, ბიოცენოლოგია, ისტორიული ბიოგეოგრაფია და სხვ. დადგენილ იქნა მნიშვნელოვანი კანონზომიერებები: ალოგენეზისა და აროგენეზის წესი, ბიოგენეზური კანონი, არასპეციალიზირებული წინაპრიდან წარმოშობისა და პროგრესული სპეციალიზაციის წესი, ევოლუციის შეუქცევადობის პრინციპი, ორგანოთა მულტიფუნქციონალობის და ფუნქციათა შეცვლით ორგანოთა ევოლუციის პრინციპი და სხვა. ამ პერიოდში დარვინის ევოლუციურმა თეორიამ კლასიკური სახე მიიღო.
შემდგომი პერიოდი (1901-20-იანი წლები) კლასიკური დარვინიზმის უარყოფის სახელწოდებით მოიხსენიება. დარვინის კონცეფციის ქვაკუთხედი ბუნებრივი გადარჩევის თეორიაა. ამ პერიოდში დაიწყო გადარჩევის როლის მკაცრი კრიტიკა, რასაც რამდენიმე მიზეზი ჰქონდა. რომელთაგან მთავარი ის იყო, რომ ცოცხალი ბუნების ორ ძირითად თვისებაზე - მემკვიდრულობასა და ცვალებადობის შესახებ მეცნიერული თეორია არ არსებობდა. გენეტიკასა და ევოლუციურ თეორიას შორის კონფლიქტი ძირითადად გენეტიკოსების მიერ ორი ძირითადი აღმოჩენის მემკვიდრეობის კორპუსკულური ბუნების დადგენისა და მუტაციური ცვალებადობის გამოვლენის არასწორმა ინტერპრეტაციამ გამოიწვია. საჭიროა აღინიშნოს, რომ ჯერ კიდევ დარვინის სიცოცხლეში შოტლანდიელმა მათემატიკოსმა ფ.ჯენკინმა გამოაქვეყნა სტატია (1867), რომლითაც უარყოფდა ბუნებრივი გადარჩევის თეორიას. მას თავის თანამედროვეთა მსგავსად მემკვიდრეობა განზავების უნარის მქონე სითხედ წარმოედგინა. იგი შემდეგნაირად მსჯელობდა: შემთხვევით წარმოქმნილი მემკვიდრული ცვლილება, რომელიც ბუნებრივ გადარჩევას უნდა დაექვემდებაროს, ერთეულია. ორი ერთნაირი მემკვიდრული ცვლილების მქონე ინდივიდის შეჯვარებით შთამომავლობის წარმოქმნის ალბათობა ძალზე მცირეა. თუ ერთ მშობელს ახალი A ნიშანი ექნა, შვილებს ამ ნიშნის A/2 (ნახევარი) აღმოაჩნდებათ, შვილიშვილებს A/4 (მეოთხედი), შვილთაშვილებს A/8 და ა.შ. ამრიგად, ყოველ მომდევნო თაობაში ძველთან შეჯვარების კვალობაზე ახალი ნიშანი თანდათანობით განზავდება ძველში და ბოლოს გაქრება. ჯენკინის ამ მოსაზრების უკუგდება მხოლოდ გენეტიკის მონაცემებით შეიძლებოდა.
არსებობდა სხვა შრომებიც, რომლებშიც აკრიტიკებდნენ დარვინიზმს. კერძოდ, გენეტიკის ფუძემდებლებმა (ჰ. დე ფრიზი, უ. ბეტსონი, ვ. იოჰანსენი) ვერ შეძლეს მართებულად შეეფასებინათ თავიანთი აღმოჩენის უმნიშვნელოვანესი როლი ევოლუციური თეორიის განვითარებაში. ვ.იოჰანსენმა ჩატარებული ცდებით (1903) დაასაბუთა გადარჩევის არაეფექტურობა „წმინდა ხაზებში“. ჰ. დე ფრიზმა „მუტაციური თეორია“ დაუპირისპირა ბუნებრივი გადარჩევის თეორიას. მისი აზრით, მუტაციები უშუალო დასაბამს აძლევენ ახალ გარემო პირობათა მიმართ შეგუებულ სახეობებს.
ჯ.ლოტსის „ჰიბრიდოგენეზის“ კონცეფციაში აღინიშნებოდა კომბინაციური ცვალებადობის ევოლუციური როლის აბსოლუტიზაცია. ამ პერიოდში გენეტიკოსები ევოლუციის მთავარ მამოძრავებელ ფაქტორად მემკვიდრულ ცვალებადობას თვლიდნენ. ბუნებრივი გადარჩევა, მათი აზრით, მეორეხარისხოვანი იყო. იგი მხოლოდ საზიანო მუტაციებისა და ნაკლებ შეგუებულ ჰიბრიდულ ვარიანტთა „საცერის“ როლს ასრულებდა.
ნეოდარვინიზმის (ევოლუციის სინთეზური თეორიის) ჩამოყალიბებაში (…იანი წლები) არსებითი მნიშვნელობა ჰქონდა გენეტიკის, ეკოლოგიისა და ევოლუციური თეორიის მონაცემთა სინთეზს. გენეტიკისა და ევოლუციური თეორიის სინთეზი ნელი ტემპით მიმდინარეობდა. დიდი მნიშვნელობა ჰქონდა ა.ვაისმანის მიერ მემკვიდრეობის ქრომოსომული ჰიპოთეზის ჩამოყალიბებას და შეძენილ ნიშან-თვისებათა მემკვიდრეობის შეუძლებლობის პრინციპს. მნიშვნელოვანი იყო ასევე ჯ. ჰარდისა და ვ. ვაინბერგის მიერ პოპულაციაში ალელთა სიხშირის წონასწორობის პრინციპის ფორმულირება (1908). სინთეზის პროცესზე გადამწყვეტი მნიშვნელობა შეასრულა ს.ჩეტვერიკოვის შრომამ „ევოლუციური პროცესის ზოგიერთი კანონზომიერებების შესახებ თანამედროვე გენეტიკის თვალთახედვით“ (1926). მან დაასაბუთა, რომ პოპულაციაში მიმდინარე მუტაციური პროცესი იძლევა ახალ მასალას და ამდიდრებს გენოფონდს, რაც აპირობებს გენეტიკურ ჰეტეროგენულობას. ბუნებრივი გადარჩევის მოქმედებით ამ მასალის „გადამუშავება“ მიმდინარეობს და ხორციელდება ევოლუციის პროცესი. 1928-1930 წწ. ჩატარებული გამოკვლევებით (ნ. დუბინინი და დ. რომაშოვი, ს. რაიტი და სხვ.) ნაჩვენები იქნა, რომ ევოლუციის პროცესში უდიდეს როლს ასრულებს არა მარტო მუტაციური პროცესი, არამედ გენური სიხშირეების არამიმართული ცვლილებებიც (გენთა დრეიფი). რ.ფიშერმა მათემატიკურად დაასაბუთა (1930) ევოლუციური ცვლილების მიმდინარეობა ბუნებრივი გადარჩევის მოქმედებით. 1937-1939 წწ. ფ. დობჟანსკიმ და ნ. ტიმოფეევ-რესოვსკიმ ჩამოაყალიბეს მოძღვრება მიკროევოლუციის შესახებ. ექსპერიმენტული გამოკვლევებით დაფორმულირდა შეხედულებები ელემენტარულ ევოლუციურ სტრუქტურაზე (პოპულაცია), მასალაზე და მოვლენაზე. ჯ.ჰაქსლის შრომამ „ევოლუცია: თანამედროვე სინთეზი“ (1942) დააჩქარა ევოლუციის სინთეზური თეორიის დაფუძნება.
ევოლუციის სინთეზური თეორიის ჩამოყალიბებაში არსებითი როლი შეასრულა დარვინიზმის ბაზაზე გენეტიკურ-პოპულაციურმა გამოკვლევებმა, არსებობისათვის ბრძოლის მათემატიკურმა და ექსპერიმენტულმა გამოკვლევებმა, სახეობის თეორიის დამუშავებამ, მაკროევოლუციის ზოგი სხვა პრობლემების შესწავლამ. ევოლუციის სინთეზური თეორიის ფუძემდებლებია ფ.დობჟანსკი, ე.მაირი, ჯ.სიმპსონი, ჯ.ჰაქსლი და ჯ.სტებინსი.
|
|
4 §2. პოპულაცია - ევოლუციური როცესის ელემენტარული ერთეული |
▲ზევით დაბრუნება |
ბიოლოგიის ნებისმიერ დარგში ზუსტი ანალიზის ჩატარება იმ შემთხვევაშია შესაძლებელი, როდესაც ხერხდება ელემენტარული ერთეულის გამოყოფა და შესწავლა, რომლის მონაწილეობით გარკვეული მოვლენა მიმდინარეობს. ბუნებრივია, ამგვარი ერთეული სიცოცხლის გარკვეული ორგანიზაციული დონისთვისაა ელემენტარული, ხოლო მისი შემადგენელი ნაწილები ამ თვისებას მოკლებულია. გენეტიკაში ასეთ ერთეულს წარმოადგენს გენი, სისტემატიკაში - სახეობა, ეკოსისტემაში - ეკოცენოზი და ა.შ. იგი სიცოცხლის დისკრეტულ ბუნებას გამოხატავს. გენეტიკური იდეისა და მეთოდოლოგიის გამოყენებით შეიძლება გამოიყოს ევოლუციური პროცესის ის ერთეული, რომლის მეშვეობითაც შეიძლება მიკროევოლუციის შესწავლა და ელემენტარული ევოლუციური ერთეულის დადგენა. მიკროევოლუცია ევოლუციის საწყისი ეტაპია, რომელიც სახეობის შიგნით მიმდინარეობს, იწვევს მის დიფერენცირებას და სახეობათა წარმოქმნით მთავრდება.
ვინაიდან ბიოლოგიური ევოლუცია წარმოადგენს ორგანიზმების ცვლილებისა და დივერგენციის პროცესს დროში, ამდენად საჭიროა განისაზღვროს ამ პროცესის ელემენტარული ერთეული, რომელიც დააკმაყოფილებს შემდეგ მოთხოვნებს:
1. აღარ უნდა ხდებოდეს მისი შემდგომი დაყოფა უფრო მარტივ ერთეულებად. ე.ი. უნდა წარმოადგენდეს ერთგვარ მთლიანობას დროსა და სივრცეში*
2. მას უნდა ჰქონდეს დროში ანუ ბიოლოგიური თაობების მანძილზე მემკვიდრული ცვლილების უნარი*
3. ბუნებრივ პირობებში იგი რეალურად და კონკრეტულად უნდა არსებობდეს. ასეთი ელემენტარული ერთეული არის პოპულაცია.
ყოველ სახეობას გარკვეული არეალი უჭირავს. ცხოველთა, მცენარეთა და მიკროორგანიზმთა ნებისმიერი სახეობის ინდივიდები მათ მიერ დაკავებულ არეალში სივრცობრივად არათანაბრად არიან განაწილებულნი. ისინი შეიძლება დასახლებული იყვნენ „კუნძულებრივად“ (ე.წ. იზოლატები) ან მჭიდროდ დასახლებული ტერიტორიები შედარებით მეჩხერად დასახლებულ ტერიტორიებს ცვლიდნენ (ე.წ. კონტინიუმი). ერთი სახეობის ინდივიდთა ერთობლიობები, რომლებიც ქმნიან ტერიტორიაზე (ხმელეთზე ან აკვატორიაში) მჭიდროდ დასახლებულ „ცენტრებს“, პოპულაციებს წარმოადგენენ. ერთი სახეობის ინდივიდთა ერთობლიობას, რომელიც ხანგრძლივად არსებობს გარკვეულ ტერიტორიაზე, შედარებით გამოცალკევებულია ამავე სახეობის სხვა ერთობლიობებისაგან, ხასიათდება მის ფარგლებში თავისუფალი შეჯვარებით და ერთიანი გენეტიკური სისტემით, აქვს თავისი სტრუქტურის შენარჩუნების უნარი, პოპულაცია ეწოდება.
ამ განმარტებიდან გამომდინარეობს, რომ პოპულაცია საკმაოდ მრავალრიცხოვანი ინდივიდებისაგან შემდგარი ისეთი ჯგუფია, რომელსაც გარკვეული ეკოლოგიური ნიში უკავია და არსებობს ხანგრძლივად - განუსაზღვრელ თაობათა მანძილზე. პოპულაციის წევრები სხვადასხვა სიხშირით - თავისუფლად ეჯვარებიან ერთმანეთს (მიმდინარეობს პანმიქსია) და სივრცობრივად მეტ-ნაკლებად იზოლირებული არიან, ამავე სახეობის სხვა პოპულაციებისაგან. აქვე უნდა შევნიშნოთ, რომ ასეთი განმარტება არის ზოგადი და არსებობს გამონაკლისი. მაგ., ასექსუალური პოპულაციები (ამება, ქლორელა და სხვ.). იგივე ითქმის არეალზე, მაგ., მიმომფრენი ფრინველები შორს მიგრირებენ და ა.შ.
ისმის კითხვა, რატომ ითვლება პოპულაცია ევოლუციის ელემენტარულ ერთეულად და არა სახეობა (როგორც ადრე თვლიდნენ) ან ცალკეული ინდივიდი (როგორც ზოგიერთ მეცნიერს მიაჩნდა). ცალკეული ინდივიდი ვერ აკმაყოფილებს იმ მოთხოვნებს, რომელიც ზემოთ იყო დასახელებული. პოპულაციაში ინდივიდი ევოლუციის მთავარი ფაქტორის - ბუნებრივი გადარჩევის მოქმედების ობიექტია, მაგრამ მას „საკუთარი ევოლუციური ბედი“ არ გააჩნია. ეს ნიშნავს, რომ ცალკეული ინდივიდი მცირე ხანს ცოცხლობს, მაშინ როდესაც ევოლუციური მასშტაბი გრანდიოზულია და ათასობით და მილიონობით წლებს ითვლის. ამა თუ იმ ინდივიდის სიცოცხლე, რაგინდ ხანგრძლივი უნდა იყოს ის, მაინც ერთი ბიოლოგიური თაობაა. მასში წარმოქმნილი ვარიაცია, მათ შორის მემკვიდრული, შესაძლოა ვერ გამოვლინდეს. ალელთა დომინანტურ-რეცესიული ურთიერთდამოკიდებულების გამო მისი გამოვლენა სათუოა (შეიძლებაგამოვლინდეს ან არა) მომდევნო თაობებში. ამდენად, ევოლუციის ელემენტარული ერთეული უნდა იყოს ინდივიდთა რომელიმე ჯგუფი (ევოლუციას ექვემდებარება არა ინდივიდი, არამედ ინდივიდთა ჯგუფი). ელემენტარული ერთეული არაა ისეთი ჯგუფი, როგორიცაა: ოჯახი, ჯოგი და სხვ., ვინაიდან მისი არსებობაც დროში ეფემერულია. სახეობა ჩაკეტილი, რთული, ჰეტეროგენული სისტემა აღმოჩნდა, მართალია მას აქვს ევოლუციური ბედი, მაგრამ მეტად მრავალრიცხოვანი წევრებისაგან შედგება, არათანაბრად არის განაწილებული სივრცეში და შედგება მცირე ერთეულებისაგან - პოპულაციებისაგან, რის გამოც იგი არაა ელემენტარული ერთეული. ამით სახეობის როლი ევოლუციის პროცესში როდი კნინდება, იგი ამ პროცესის მნიშვნელოვან ეტაპს წარმოადგენს. ამრიგად, პოპულაცია დამოუკიდებელი ევოლუციური სტრუქტურაა. იგი წარმოადგენს უმცირეს, ელემენტარულ ჯგუფს, რომელიც ორგანიზებული ერთიანი სახით ხანგრძლივად არსებობს განსაზღვრულ არეალში, ექვემდებარება ევოლუციური ფაქტორების მოქმედებას და აღენიშნება დამოუკიდებლად ევოლუციის უნარი. ამდენად, პოპულაცია ევოლუციის ელემენტარულ სტრუქტურულ ერთეულს წარმოადგენს.
|
|
5 §3. პოპულაციის ეკოლოგიური სტრუქტურა |
▲ზევით დაბრუნება |
მიკროევოლუციაში ზუსტი ანალიზის ჩატარება მას შემდეგ გახდა შესაძლებელი, რაც გამოყვეს და აღწერეს ევოლუციის ელემენტარული სტრუქტურული ერთეული - პოპულაცია. დადგენილია, რომ პოპულაციის კანონზომიერ ცვლილებას გარკვეული წვლილი შეაქვს სიცოცხლის შენარჩუნებისა და ისტორიული განვითარების (მიკროევოლუციის) პროცესში. ისტორიულად პოპულაცია ფორმირდება კონკრეტულ ეკოლოგიურ პირობებში, ამიტომაც მის დახასიათებას ახდენენ: დაკავებული ტერიტორიით (სახეობის არეალის მონაკვეთი), რიცხოვნობით, ასაკობრივი და სქესობრივი შემადგენლობით, ასევე პოპულაციის დინამიკით.
|
|
5.1 პოპულაციის არეალი |
▲ზევით დაბრუნება |
ყოველ პოპულაციას სახეობის არეალში გარკვეული ტერიტორია უკავია და გააჩნია სივრცობრივი სტრუქტურა. დაკავებული ტერიტორია (არეალი)პოპულაციის ერთ-ერთ მნიშვნელოვან კრიტერიუმს წარმოადგენს. იგი ასახავს პოპულაციის არეალში ცალკეული ინდივიდებისა და მათი დაჯგუფებების განაწილების ხასიათს. ერთი მხრივ, ეს განაწილება დამოკიდებულია პოპულაციის წევრებზე მოქმედ ნაირგვარ გარემო პირობებზე, მეორე მხრივ, პოპულაციის შემადგენელი ინდივიდების ბიოლოგიურ (გადაადგილებისა და აგრეგირების უნარი და მისთ.) თვისებებზე.
შესაძლებელია პოპულაციამ არეალი განავრცოს, მაგრამ მან ახალი ტერიტორია უნდა აითვისოს. ეს იმ შემთხვევაშია შესაძლებელი თუ ტერიტორია პოპულაციის აუცილებელი ნიშა გახდება. დაკავებული ტერიტორიით განსხვავდებიან, როგორც სხვადასხვა, ისე ერთი და იგივე სახეობის პოპულაციები. მაგალითად აღმოსავლური ნაძვი (Picea orientalis) საქართველოს დასავლეთ ნაწილში მასიურად გვხვდება და ზოგჯერ ერთ პოპულაციას ათეულობით ჰექტარი ფართობი უკავია, აღმოსავლეთ ნაწილში, კერძოდ არაგვის ხეობაში ნაძვი მცირე პოპულაციების სახითაა აღრიცხული და შესაბამისად მცირე ფართობი უკავია (გაგნიძე, 1996), მარდი ხვლიკის (Lacerta agilis) ერთმა პოპულაციამ შეიძლება 0,1-დან რამდენიმე ათეულ ჰექტარამდე ტერიტორია დაიკაოს. წყლის მემინდვრიამ (Arvicola terestris) - 1-დან რამდენიმე ჰექტრამდე.
ზოგიერთი პოპულაცია ამავე სახეობის სხვა პოპულაციებიდან მკაფიოდ არის გამიჯნული, რის გამოც ადვილია არეალის განსაზღვრა. ასეთია, მაგალითად, დიზუნქციური არეალის მქონე პოპულაციები. ზოგჯერ უწყვეტი არეალის მფლობელ სახეობაში გაერთიანებულ პოპულაციებს შორის საზღვარი გარდამავალი ფორმებით იმდენადაა წაშლილი, რომ საზღვრის დადგენა გაძნელებულია.
ნებისმიერ პოპულაციას ყოველთვის ახასიათებს წევრთა ინდივიდური გადაადგილების სპექტრი. ინდივიდთა ნაწილი მუდმივად რჩება გაჩენის ადგილზე, ნაწილი კი სხვადასხვა მანძილით განსახლდება. არეალის სიდიდე მნიშვნელოვნად არის დამოკიდებული ინდივიდთა გავრცელების ხარისხზე - რეპროდუქციული აქტიობის რადიუსზე (ამ სიდიდით გამოხატავენ გაჩენის ადგილსა და კონკრეტული თაობის 95%25 ინდივიდების გამრავლების ადგილს შორის მანძილს). თუ ასეთი რადიუსი მცირეა, მაშინ პოპულაციის არეალიც მცირეა და პირუკუ. მცენარეებში რეპროდუქციული აქტიობის რადიუსი განისაზღვრება იმ მანძილით, რომელზეც ვრცელდება მტვრის მარცვლები, თესლები, სპორები და გამრავლების უნარის მქონე ვეგეტატიური ნაწილები (იხ. ცხრ. 3.1).
ცხრილი 3.1
ზოგიერთი სახეობის რეპროდუქციული
აქტიობის რადიუსი (იაბლოკოვი, 1987)
სახეობა |
რეპროდუქციული |
დროზოფილა |
400 |
არყის ხის მზომელა |
6000 |
შუააზიისეული კუ |
700 |
რუხი ყანჩა |
600000 |
თხუნელა |
300 |
ჩვეულებრივი მელა |
100000 |
მგელი |
190000 |
შველიჩ |
10000 |
კურდღელი |
10000 |
სიასამური |
80000 |
კომბოსტო |
24 |
სიმინდი |
10-20 |
ფიჭვი |
100 |
ხახვი |
1,8 |
პომიდორი |
7.3-50.3 |
ცხოველთა არეალის განსაზღვრისას ითვალისწინებენ გადაადგილების ხარისხსა და გამრავლების თავისებურებებს.
ზოგიერთი ღამურა ზამთრის გასატარებლად საკმაოდ დიდი მანძილით (ასეულობით და ათასეულობით კმ.) მიგრირებს, მაგრამ წლობით რამდენიმე კილომეტრის რადიუსის ტერიტორიაზე მრავლდება. ამ შემთხვევაში ტროფული (ტერიტორია, რომელზედაც ინდივიდი იკვებება) და რეპროდუქციული (ტერიტორია, რომელზედაც ინდივიდი მრავლდება) ერთმანეთს არ ემთხვევა. მსგავსი მაგალითების დასახელება ხერხემლიან ცხოველთა სხვა კლასებიდანაც შეიძლება. მაგ., ორაგული ზღვაში ბინადრობს, ხოლო მდინარეში ტოფობს. დიდი ტროფული არეალი ნიშანდობლივია მიმომფრენი ფრინველებისათვის., მაგ. ყარყატი (Ciconia ciconia) ზაფხულს ევროპაში ატარებს და ბუდობს, ხოლო ზამთრობს აფრიკაში. იგი მონოგამური ფრინველია და თითოეული წყვილი ევროპაში ძველ ბუდეს უბრუნდება. მართალია ზამთრობისას მრავალი განსხვავებული პოპულაციის ინდივიდები ერთმანეთს ერევა, მაგრამ გამრავლებისას მცირე ტერიტორია უკავია. ამგვარ ცხოველებში გამოყოფენ ტროფულ და რეპროდუქციულ არეალს, ასევე მიგრაციის გზებს (იხ. სურ. 3.1). ევოლუციურ-გენეტიკური თვალთახედვით მნიშვნელოვანია რეპროდუქციული არეალი
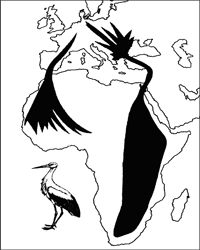
სურ. 3.1. თეთრი ყარყატის (Ciconia ciconia) ტროფული არეალი და მიგრაციის გზები (d. mak-farlendi, 1988).
|
|
5.2 ინდივიდთა რიცხოვნობა პოპულაციაში |
▲ზევით დაბრუნება |
ინდივიდთა ერთობლიობის - პოპულაციის შემადგენელი წევრები არეალში არათანაბრადაა განაწილებული. ეს გამოწვეულია ფიზიკურ-გეოგრაფიული და სხვა პირობების არაერთგვაროვნობით, რაც საკვები რესურსების, თავშესაფარის, ამა თუ იმ სქესის, ასაკის, მორფო-ფიზიოლოგიური ჯგუფებისთვის ხელსაყრელი ადგილების არაერთგვაროვან, შეზღუდულ განაწილებას განაპირობებს. ამდენად, არეალში მხოლოდ განსაზღვრული რაოდენობის ინდივიდებს შეუძლია არსებობა.
რიცხოვნობა (ინდივიდთა რაოდენობა კონკრეტულ ტერიტორიაზე) პოპულაციის მნიშვნელოვანი მახასიათებელია. არეალის ფართობის შესაბამისად პოპულაციის რიცხოვნობა მნიშვნელოვნად შეიძლება შეიცვალოს. ნორმალურ პირობებში კონკრეტულ პოპულაციას რიცხოვნობის სპეციფიკური სიდიდე გააჩნია. მაგალითად თბილისის მიდამოების ტერიტორიაზე მობინადრე ჩვეულებრივი ტრიტონის (Triturus vulgaris) რიცხოვნობა ასეულიდან - 1500-მდე ინდივიდს შეადგენს, ხოლო სავარცხლიანი ტრიტონისა (T. cristatus) - რამდენიმე ათეულს არ აღემატება (თარხნიშვილი, 2004). ღია სივრცეებზე გავრცელებული ზოგიერთი მწერებისა და ერთწლოვანი მცენარის პოპულაციებში ინდივიდთა რაოდენობა ზოგჯერ ათეულ ათასს და მილიონსაც აღწევს.
პოპუცლაციის რიცხოვნობას უკავშირდება მინიმალური რიცხოვნობის პრობლემა. მინიმალური რიცხოვნობა ეს არის ის ზღვრული რაოდენობა რომლის შემდეგაც ინდივიდთა რიცხვის შემცირება, ეკოლოგიურ-გენეტიკური მიზეზების გამო, პოპულაციის გადაშენებას იწვევს. ეს მაჩვენებელი სხვადასხვა სახეობისათვის განსხვავებული და სპეციფიკურია. შეუძლებლადაა მიჩნეული ისეთი პოპულაციების ევოლუციურად ხანგრძლივად არსებობა, რომელთა წევრები რამდენიმე ასეულზე ნაკლებია. მაგ., ამურის ვეფხვი ერთი პოპულაციისაგან შედგება, რომელშიაც 400-მდე ინდივიდია. მიუხედავად დაცვის მკაცრი ზომებისა, შემთხვევითმა (სტოქასტურმა) ფაქტორებმა (ხანძარი, წყალდიდობა და სხვ.) შეიძლება ამ რიცხვის შემცირება გამოიწვიოს, ერთეულმა ინდივიდებმა გამრავლების პერიოდში საჭირო სიხშირით ერთიმეორე ვეღარ იპოვონ. დასაშვებია, მათი რიცხვი იმდენად შემცირდეს, რომ შობადობამ დანაკლისი ვეღარ დაფაროს და რამდენიმე თაობაში ამოწყდნენ. ამ მდგომარეობაში შეიძლება აღმოჩნდეს მრავალი იშვიათი მცენარე და ცხოველი.
|
|
5.3 ხანმოკლე დაჯგუფებები |
▲ზევით დაბრუნება |
ხშირად ბუნებაში პოპულაციებისა და შიგაპოპულაციური დაჯგუფებების გარჩევა რთულია. ხანმოკლე დაჯგუფებები, თუნდაც მრავალრიცხოვანი, ბუნებრივ პოპულაციად არ შეიძლება ჩაითვალოს. მაგალითად, ბუნებრივი არეალიდან საკმაოდ დაცილებულ კუნძულზე (არქიპელაგი ტვარმინი, ფინეთის ყურე) ნახნავის მემინდვრია (Microtus agrestis) გამოჩენიდან, სამ წელზე მეტი ხნით არ აღინიშნებოდა. ზოგიერთ დიდ კუნძულზე მათი რიცხოვნობა ასეულობით ინდივიდს აღწევდა. მიუხედავად ამისა არც ერთ კუნძულზე მემინდვრიები პოპულაციად ვერ ჩამოყალიბდა (იხ. სურ. 3.2).

სურ. 3.2. ნახნავის მემინდვრიას (microtus agrestis) გავრცელება ტვარმინის არქიპელაგის სამ კუნძულზე (ჯ. პოკი, 1981).
შუააზიის უდაბნოებისა და ნახევრადუდაბნოების ბინადარი საჯას (Syrrhaptes paradoxus) გუნდები ზოგიერთ წელს არეალიდან რამდენიმე ათასი კილომეტრის დაშორებითაა აღრიცხული. მრავალი ფრინველი ჩრდილო-დასავლეთ ევროპამდე აღწევს, მაგრამ ერთი ან ორი ბუდობის შემდეგ უკვალოდ ქრება. საჯები ახალ ტერიტორიას ვერ ითვისებენ, ვინაიდან არ ხდება საკუთარი გენეტიკური სისტემის ფორმირება. ბუნებრივია ისინი დამოუკიდებელ პოპულაციას არ წარმოადგენენ.
პოპულაციად არ შეიძლება მივიჩნიოთ აგრეთვე, წითურმუცელა მემინდვრიას (achrogaster), ტბის (კრაბ-ორჩარდი, აშშ) 26 კუნძულზე განსახლებული მცირე ჯგუფები. მემინდვრიები ერთ ან ორ თაობას იძლევიან კუნძულებზე, შემდეგ კი ელიმინირდებიან. ძველის ადგილს მატერიკიდან მიგრირებული ინდივიდები იკავებენ. მიუხედავად იმისა, რომ კუნძულებზე გამუდმებით სახლობენ მემინდვრიები ისინი დამოუკიდებელ პოპულაციას არ წარმოადგენენ. მათ არ გააჩნიათ საკუთარი გენოფონდი და არეალი. კონკრეტულ შემთხვევაში პოპულაცია არის მატერიკისა და კუნძულებზე მოსახლე ინდივიდების ერთობლიობა. ასეთი საკმაოდ დიდი დაჯგუფება ხანგრძლივად არსებობს დამოუკიდებელი ევოლუციური გაერთიანების სახით.
|
|
5.4 პოპულაციის დინამიკა |
▲ზევით დაბრუნება |
„რიცხოვნობის ტალღების“ დიდი ევოლუციური მნიშვნელობა და ცოცხალ სამყაროში მათი საყოველთაო ხასიათი რუსი გენეტიკოსის ს. ჩეტვერიკოვის (1905) შრომებით გახდა ცნობილი. პოპულაციის საზღვრები, ასევე მისი ყველა სტრუქტურა, დინამიურია. მისი ყველა პარამეტრი რომელიმე საშუალო სიდიდის მახლობლად განუწყვეტლივ ვარირებს. პოპულაციის რიცხოვნობა დროსა და სივრცეში პერიოდულად მერყეობს. ფლუქტუაციას იწვევს ერთი მხრივ, მრავალი გარეგანი აბიოტური და ბიოტური ფაქტორი, მეორე მხრივ, თვით პოპულაციის მახასიათებლებს შორის ურთიერთდამოკიდებული შინაგანი ფაქტორები.
რიცხოვნობის ფლუქტუაციას შეიძლება აციკლური ან ციკლური ხასიათი ჰქონდეს. მაგალითად, ინგლისის ერთ-ერთ პატარა კუნძულზე ველურად ბინადრობს ბოცვერი (Oryctolagus cuniculus). გაზაფხულ-ზაფხულში უხვი საკვების მოპოვებით შემოდგომისთვის, ინდივიდებმა მაქსიმალურ რაოდენობას - 1000-ს მიაღწია. ერთხელ მკაცრმა ზამთარმა და საკვების ნაკლებობამ მათი რაოდენობა 100-მდე შეამცირა. რიცხოვნობის უფრო დიდი სეზონური მერყეობა ზოგიერთ წლებში აღენიშნებათ თაგვისებრ მღრღნელებს და მწერებს (კალიები, მაისის ხოჭო, კოლორადოს ხოჭო და სხვ.), სადაც ინდივიდთა რაოდენობა მილიონჯერ და მეტჯერ მატულობს.
სეზონური ცვლილებების გარდა მცენარეებსა და ცხოველებში ცნობილია რიცხოვნობის არასეზონური (პერიოდული და არაპერიოდული) ცვლილებები (იხ. სურ. 3.3). ევოლუციის პროცესში რიცხოვნობის მერყეობა დიდ როლს ასრულებს, იგი მკვეთრად და არამიმართულად ცვლის პოპულაციის გენეტიკურ სტრუქტურას (იხ. § 7.2; 7.3.).
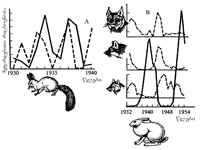
სურ. 3.3. ინდივიდთა რიცხოვნობის დინამიკა პოპულაციებში: A- ციყვის (Sciurus vulgaris) კოსტრომის მხარეში (პუნქტირით ნაჩვენებია ნაძვის თესლის მოსავალი); B - კურდღლის (Lepus timidus), ფოცხვერის (Felis lynx), მგლის (Canis lupus) და მელიის (Vulpes vulpes) რუსეთის ჩრდილოეთ ევროპული ნაწილის პოპულაციებში (ა. იაბლოკოვი, ა. იუსუპოვი, 2004).
|
|
5.5 პოპულაციის ასაკობრივი სტრუქტურა |
▲ზევით დაბრუნება |
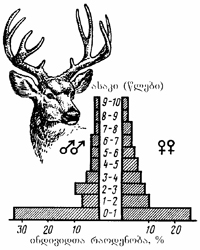
სურ. 3.4. კალიფორნიაში გავრცელებული შავკუდა ირმის (Odocoileus hemionus) პოპულაციაში ფურირემის და ხარირემის ასაკობრივი პირამიდა (რ. დაჟო, 1975).
პოპულაციას აქვს ასაკობრივი სტრუქტურა. ამრიგად პოპულაცია დიფერენცირებულია როგორც სივრცეში (არეალი), ისე დროში (შედგება სხვადასხვა ასაკის ინდივიდებისაგან). ხშირად სახეობის შემადგენელ ცალკეულ პოპულაციას მისთვის ნიშანდობლივი ასაკობრივი თანაფარდობა ახასიათებს. ამ თანაფარდობაზე მოქმედებს ორგანიზმთა სიცოცხლის ხანგრძლიობა, გამრავლების ინტენსიობა, თუ როდის აღწევენ სქესობრივ სიმწიფეს და სხვ. სწრაფად მზარდი პოპულაციები ძირითადად ახალგაზრდა ინდივიდებისაგან შედგებიან. სტაციონარულ პოპულაციებში ყველა ასაკის ინდივიდები გვხვდება და მათი თანაფარდობა თითქმის თანაბარია (იხ. სურ. 3.4). კლებადი რიცხოვნობის პოპულაციებში ხანდაზმული ჯგუფები ჭარბობენ. ნებისმიერი სტაციონარული პოპულაციისათვის ნიშანდობლივია სტაბილური ასაკობრივი სტრუქტურის შენარჩუნების ტენდენცია. მცენარეთა და ცხოველთა პოპულაციებში ძირითადად სამ ეკოლოგიურ ასაკობრივ ჯგუფს გამოყოფენ: პრერეპროდუქციულს, რეპროდუქციულს და პოსტრეპროდუქციულს. ადამიანში ასაკობრივი ჯგუფების თანაფარდობა თანაბარია და თითოეულის ხანგრძლიობა დაახლოებით 1/3-ს შეადგენს.
მეტად მარტივია ზოგიერთი პოპულაციის ასაკობრივი სტრუქტურა. მაგალითისათვის განვიხილოთ ეფემერული ძუძუმწოვრის ბიგას (Sorex) პოპულაცია. ისინი გაზაფხულზე ერთ ან ორ ნაყარს იძლევიან. ზრდასრული ინდივიდები მალე იღუპებიან. შემოდგომით პოპულაცია მხოლოდ სქესობრივად მოუმწიფებელი ინდივიდებისაგან შედგება. გაზაფხულზე ყველა გამოზამთრებული ინდივიდი სქესმწიფეა და ციკლი კვლავ მეორდება.
საკმაოდ რთული, მდგრადი ასაკობრივი სტრუქტურა გააჩნია ხანგრძლივად მცხოვრებ თანასაზოგადოებრივ ძუძუმწოვრებს. მაგ., ინდური სპილო (lephas maximus) ბინადრობს 10-20 ინდივიდისგან შემდგარ ოჯახურ ჯოგებში. სქესმწიფობას 8-12 წლის ასაკში აღწევს. ცოცხლობს 60-70 წელს. მდედრი ოთხ წელში ერთხელ შობს 1, იშვიათად 2 ნაშიერს. ჯოგში ზრდასრული სხვადასხვა ასაკის წევრები 80%25 შეადგენს, მოზარდები კი 20%25-ს. გამრავლების მაღალი ინტენსიობის ზოოსოციალურ ძუძუმწოვრებში ასაკობრივი სტრუქტურა გაცილებით რთულია. მაგ., პოლარულ ზღვებში ბინადარ დელფინ თეთრანას (elphinopterus leucas) გუნდი რამდენიმე ასაკობრივი ჯგუფისაგან შედგება. პირველ ასაკობრივ ჯგუფში წლამდე ასაკის ნაშიერებია გაერთიანებული, მეორეში - წინა წელს დაბადებულები, მესამეში - 2-3 წლის სქესმწიფე ინდივიდები, მაგრამ ისინი არ მრავლდებიან; მეოთხეში - 4-20 წლის ასაკის ინდივიდები, რომლებიც მრავლდებიან და სხვადასხვა ასაკის ინდივიდები ერთმანეთს ეჯვარებიან. ასაკობრივი სტრუქტურა მკაფიოდაა გამოხატული მრავალ ხემცენარეში. მაგ., მუხნარში ერთდროულად სხვადასხვა ასაკის მცენარეები გვხვდება. ახალგაზრდა 20 წლის მცენარემ შეიძლება დამტვეროს 1000 წლის (მშობელი) მცენარე.
|
|
5.6 სქესობრივი სტრუქტურა |
▲ზევით დაბრუნება |
ერთი შეხედვით სქესის მიხედვით თანაფარდობის შესწავლა მნიშვნელოვანია მხოლოდ ცალსქესიან ცხოველებსა და ორსახლიან მცენარეების პოპულაციებში. თითქოს მას მნიშვნელობა არ უნდა ჰქონდეს აგამურ, ჰერმაფროდიტ და პართენოგენეზურ პოპულაციებისათვის. ასეთი შეხედულება მცდარია, რადგან აგამურ ფორმებში აღმოჩენილია სქესობრივი პროცესის თავისებური ვარიანტი, ჰერმაფროდიტებში ობლიგატური თვითგანაყოფიერება მეტად იშვიათია, ხოლო ზოგიერთ პართენოგენეზურ პოპულაციებში აღმოჩენილია მამრები. ყოველივე ეს იმაზე მიგვანიშნებს, რომ სქესთა თანაფარდობა პოპულაციის მნიშვნელოვანი მახასიათებელია.
პოპულაციაში არჩევენ სქესის სამი სახის თანაფარდობას. პირველადი თანაფარდობა ზიგოტების წარმოქმნისას ყალიბდება. სქესის განსაზღვრას საფუძვლად უდევს ქრომოსომული მექანიზმი (ე.წ. ეპიგამური ანუ გენოტიპური ტიპი). მდედრი და მამრი ინდივიდები დაახლოებით თანაბარი რაოდენობით ჩაისახებიან (თანაფარდობაა დაახლოებით 1:1).
მამრობითი და მდედრობითი სქესის განსხვავებული სიცოცხლის უნარიანობის გამო, მეორეული თანაფარდობა (დაბადებულ ან გამოჩეკილ ინდივიდებს შორის) განსხვავდება პირველადისაგან. მესამეული თანაფარდობა სქესმწიფე ინდივიდებისათვის არის დამახასიათებელი. მაგ., ადამიანებში ყოველ ახალშობილ 100 ქალზე 106 ვაჟი მოდის (მეორეული თანაფარდობა), 16-18 წლის ასაკში ეს თანაფარდობა, ვაჟების მაღალი სიკვდილიანობის გამო, თანაბრდება. 50 წლის ასაკში ყოველ 100 ქალზე 85 მამაკაცი მოდის, ხოლო 80-წლის ასაკში ყოველ 100 ქალზე - 50 მამაკაცი.
ცხოველებისა და მცენარეების პოპულაციებში სქესის მეორეული და მესამეული თანაფარდობა ხშირად მნიშვნელოვან ფარგლებში ვარირებს. მაგ., დროზოფილაში სქესის თანაფარდობის ცვლილებას იწვევს მდედრის ნაყოფიერება, პოპულაციის სიმჭიდროვე, მამრების სიცოცხლისუნარიანობაზე მოქმედი სქესობრივი გადარჩევა. ზოგიერთ მწერებსა და რიგ კიბოსნაირებში სქესის მოდიფიკაციას ახდენენ ციტოპლაზმური სიმბიონტი ან პარაზიტი ბაქტერიები. ისინი იწვევენ ფემინიზაციას, პართონეგენეზს ან ანდროციდს რის გამოც შთამომავლობა მხოლოდ მდედრებისაგან შედგება. საზოგადოებრივ მწერებში (ფუტკრები, ჭიანჭველები და მისთ.) მდედრი ინდივიდების რაოდენობას სპეციფიკური საკვებით მუშა ინდივიდები არეგულირებენ. ზოგიერთი პართენოგენეზური პოპულაცია (მწერები, ხვლიკები და სხვ.) მხოლოდ მდედრებისგან შედგება, სხვადასხვა პოპულაციებში პართენოგენეზული მდედრების რაოდენობა მნიშვნელოვნად ვარირებს.
სქესთა თანაფარდობაში მეორე უკიდურესი ფორმაა ევოლუციურად ჩამოყალიბებული სქესის განსაზღვრის ლაბილური (ე.წ. ეპიგამერი ანუ მოდიფიკაციური) სისტემა. მაგ., ზღვის ბინადარი ჭია ბონელიას (Bonellia viridis) ზიგოტიდან განვითარებული, თავისუფლად მცხოვრები ბისექსუალური ლარვა მდედრის ხორთუმზე მოხვედრისას, ჩადის საშვილოსნოში და მამრად ყალიბდება, ზღვის ფსკერზე მოხვედრისას - მდედრად (იხ. სურ. 3.5). მსგავს მოვლენას ვხვდებით მცენარე არესემაში (Arisaema Japonica). სქესი დამოკიდებულია მშობლის მიერ წარმოქმნილი ტუბერის ზომაზე. დიდი ზომის, საკვებით მდიდარი ტუბერიდან მდედრობითი, ხოლო პატარა, საკვებით ღარიბი (ფშუტე) ტუბერიდან, მამრობითი სქესის მცენარეები ვითარდებიან.
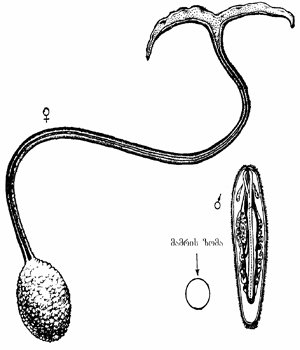
სურ. 3.5. მდედრი და მამრი ბონელია (Bonellia viridis). მდედრის სიგრძე 7 სმ-მდეა, გააჩნია 1მ-მდე სიგრძის ხორთუმი. მამრის სიგრძე 1-3 მმ-ია, რომელიც მდედრის საშვილოსნოში პარაზიტობს (ს. ინგე-ვეჩტომოვი, 1989).
|
|
5.7 პოპულაციის ეკოლოგიური ერთიანობა |
▲ზევით დაბრუნება |
პოპულაციისთვის ნიშანდობლივია მაღალდიფერენცირებული სტრუქტურა, რაც ვლინდება მორფო-ფიზიოლოგიურად განსხვავებული ჯგუფების არსებობით. პოპულაციის სტრუქტურის დიფერენცირება ემყარება გენეტიკურ საფუძველს, რომელიც ტერიტორიის ოპტიმალურად ათვისებისა და საარსებო რესურსების ეფექტურ გამოყენებისკენაა მიმართული. იგი ჩართულია პოპულაციის რეგულაციის ისეთ მნიშვნელოვან პარამეტრებში, როგორიცაა რიცხოვნობა, ასაკობრივი და სქესობრივი შემადგენლობა. პოპულაციის სტრუქტურა გარკვეულ სივრცეში დიფერენცირდება. ამდენად, მის ძირითად თავისებურებას საკუთარი ეკოლოგიური ნიშის დიფერენცირება წარმოადგენს. მაშასადამე, კონკრეტულ პოპულაციას მისთვის ნიშანდობლივი ეკოლოგიური სტრუქტურა გააჩნია. ამ სტრუქტურის ცოდნა აუცილებელია პოპულაციის, როგორც ევოლუციის ელემენტარული ერთეულის ღრმად შესასწავლად.
|
|
6 §4. პოპულაციის გენეტიკური სტრუქტურა |
▲ზევით დაბრუნება |
გენეტიკა შეისწავლის ორგანიზმთა გენეტიკურ კონსტიტუციას და შთამომავლობაში მემკვიდრული ინფორმაციის გადაცემის კანონებს. ევოლუციური გენეტიკა არის გენეტიკის განხრა, რომელიც იკვლევს სხვადასხვა სახეობების პოპულაციების გენეტიკურ სტრუქტურას და მის ცვლილებას თაობათა უწყვეტ რიგში. ევოლუციურ პროცესს საფუძვლად უდევს თაობებში მომხდარი მემკვიდრული ცვლილებები. ბუნებრივი პოპულაციები გენეტიკური ნაირგვარობით ხასიათდებიან, რაც მათში მიმდინარე მუტაციური და რეკომბინაციური ცვალებადობის შედეგია. ახალი სახეობების წარმოქმნა წინარე სახეობის წიაღში მის დივერგენტულ იზოლირებულ (შეუჯვარებად) ჯგუფებად დანაწევრებით იწყება. პოპულაცია წარმოადგენს იმ პირველად კერას სადაც ბუნებრივი გადარჩევისა და მიკროევოლუციის სხვა მამოძრავებელი ფაქტორების მოქმედებით ახალი ფორმები ყალიბდება. პოპულაციის ფორმირებისა და მისი დინამიკის პროცესები შეადგენენ მიკროევოლუციას. პოპულაციის გენეტიკური სტრუქტურის შესწავლას საფუძველი ჩაუყარა დანიელმა გენეტიკოსმა ვ.იოჰანსენმა, ნაშრომით „მემკვიდრეობის შესახებ პოპულაციებსა და წმინდა ხაზებში“ (1903).
|
|
6.1 ავტოგამური და ალოგამური პოპულაციები |
▲ზევით დაბრუნება |
გამრავლება ავტოგამურ პოპულაციებში თვითგანაყოფიერებით (მცენ. ინცუხტი, ცხოვ. ინბრიდინგი) ხორციელდება. ვ. იოჰანსენმა თვითმტვერია მცენარეებში (ლობიო, ქერი, ბარდა) შეისწავლა რაოდენობრივი ნიშნები, რომლებიც ოლიგოგენებით განისაზღვრებოდა. მან დაადგინა, რომ თვითმტვერია მცენარეთა პოპულაციები შედგებოდა გენეტიკურად განსხვავებული ხაზებისაგან (ჰომოზიგოტური ინდივიდიდან მიღებული გენოტიპურად ერთგვაროვანი შთამომავლობა), რომლებიც ერთმანეთს ვერ ეჯვარებიან. ყოველ ასეთ ხაზში თვითდამტვერვით მიიღწევა ჰომოზიგოტურობის მაღალი დონე, რის გამოც ცალკეული მათგანი გენეტიკურად წმინდაა. წმინდა ხაზებში ცალკეულ ნიშანსა და თვისებას შორის განსხვავება მოდიფიკაციური ანუ პარატიპური ცვალებადობის შედეგია. ამიტომ წმინდა ხაზებში, ნებისმიერი ნიშნის მიხედვით გადარჩევა არაეფექტურია.
სადღეისოდ მოგვეპოვება საკმაო ფაქტები, რომლებიც მეტყველებენ იმაზე, რომ აბსოლუტურად ავტოგამური ორგანიზმები ბუნებაში მეტად იშვიათია. ავტოგამური პოპულაციების ცალკეული ინდივიდები დაბალი სიხშირით მაინც ეჯვარებიან ერთმანეთს და შესაბამისად ცვლიან მემკვიდრულ ინფორმაციას. აგრეთვე თვითმტვერია მცენარეებს გააჩნიათ გენები, რომლებიც ზღუდავენ თვითდამტვერვას. ავტოგამურ ორგანიზმებში მაღალი სიხშირით ინდუცირდება სპონტანური მუტაციები, რაც მათ ჰომოზიგოტურობას არღვევს.
ბუნებაში გვხვდება აგამური ორგანიზმები (ზოგიერთი პროტოზოა, სოკო, წყალმცენარე და მისთ.). მათ ევოლუციის პროცესში დაკარგეს სქესობრივი გამრავლების უნარი. აგამურ ინდივიდებს შორის არ მიმდინარეობს ინფორმაციის გაცვლა. ამ შემთხვევაში გადარჩევის ობიექტი არის ცალკეული კლონი (გენეტიკურად იდენტური შთამომავლობა). ვინაიდან განსხვავებული კლონური ინდივიდები ერთმანეთს ვერ ეჯვარებიან, ამიტომ პოპულაციის შემადგენელ კლონებს შორის გენეტიკური ერთიანობა (ინტეგრაცია) მეტად დაბალია. ბუნებაში აგამური პოპულაციების არსებობას განსაზღვრავს განსხვავებული გენოტიპების სიმბიოტურ დამოკიდებულებაზე დამყარებული გადარჩევა.
ალოგამური (ჯვარედინმტვერია მცენარეები, ცალსქესიანი ცხოველები) პოპულაციები გენეტიკურად განსხვავებული ინდივიდების თავისუფალი შეჯვარებით - პანმიქსიით ფორმირდება. ნაირგვარი გამეტების შემთხვევითი შეხვედრა განსაზღვრავს პოპულაციაში ყოველი მომდევნო თაობის გენოტიპური და შესაბამისად ფენოტიპური კლასების თანაფარდობას. მაშასადამე პოპულაციის გენეტიკური სტრუქტურა განსხვავებული გამეტების შემთხვევითი შერწყმით ყალიბდება. პოპულაციაში კონკრეტული გენოტიპის მფლობელი ინდივიდების რიცხოვნობა განსხვავებულ ალელთა სიხშირეზეა დამოკიდებული. ერთსა და იმავე პოპულაციის საარსებო პირობებთან უკეთ შეგუებულ ინდივიდებში დიდი რაოდენობით წარმოიქმნება გამეტები, ვიდრე ნაკლებ შეგუებულებში. ამდენად პოპულაციაში ამა თუ იმ გენის შეხვედრის სიხშირეს ბუნებრივი გადარჩევა განსაზღვრავს. ალოგამურ პოპულაციებში მემკვიდრეობა დისკრეტული მემკვიდრეობის კანონზომიერების შესაბამისად ხორციელდება. ხშირად მათ „მენდელურ პოპულაციებს“ უწოდებენ. მართებულია ალოგამურ ერთობლიობებს ეწოდოს პანმიქტური პოპულაციები, ვინაიდან მათ არსებობას როგორც მენდელის კანონზომიერებები, ისე მიკროევოლუციის ყველა ფაქტორის ურთიერთქმედება განსაზღვრავს.
|
|
6.2 გენოტიპებისა და ალელების სიხშირე |
▲ზევით დაბრუნება |
ინდივიდთა თავისუფალი შეჯვარების (პანმიქსიის) შედეგად პოპულაციაში იქმნება ერთიანი გენეტიკური სტრუქტურა - გენოფონდი. ამრიგად, პოპულაციის გენთა ერთობლიობას გენოფონდი ეწოდება. როდესაც პოპულაცია N რაოდენობის დიპლოიდი ინდივიდებისაგან შედგება, მაშინ მას 2N რაოდენობის გენები გააჩნია, ასევე ცალკეული ალელების რაოდენობა 2N-ის ტოლია. გამონაკლისია სასქესო ქრომოსომაში ლოკალიზებული გენები.
პოპულაციის უმნიშვნელოვანეს მახასიათებლად ალელებისა და გენოტიპების სიხშირე ითვლება. პოპულაციის გენეტიკურ შესწავლას გენების სიხშირისა და გენოტიპების თანაფარდობის განსაზღვრის გზით ახდენენ. პოპულაცია ღია გენეტიკური სისტემაა, რაც იმას ნიშნავს, რომ პოპულაციათა შორის ხდება მიგრაცია. შესაბამისად მათ შორის მიმდინარეობს გენეტიკური ინფორმაციის გაცვლა (ე.წ. გენთა ნაკადი). მაშასადამე ბუნებრივი პოპულაციები განსხვავდებიან ალელთა შეხვედრის სიხშირით და ამა თუ იმ გენოტიპის რაოდენობრივი თანაფარდობით. პოპულაციას აანალიზებენ არა უშუალოდ გენებისა და გენოტიპების, არამედ ფენოტიპების დისკრეტული ერთეულის - ფენის მიხედვით. ამ სახის ფენს ზოგჯერ მარკერულ ნიშანს უწოდებენ. პოპულაციათა შესწავლისა და ურთიერთშედარებისას მოსახერხებელი და საიმედოა მრავალი განსხვავებული ფენის მიხედვით მათი ანალიზი. ფენთა სიხშირის მიხედვით ადგენენ მათი განმსაზღვრელი გენების შეხვედრის სიხშირეს და წარმოქმნილ გენოტიპთა თანაფარდობას პოპულაციაში. ასევე ადგენენ პოპულაციის საზღვრებს, სხვადასხვა პოპულაციათა შორის მსგავსება-განსხვავების დონეს. ასევე შესაძლებელი ხდება ცალკეული პოპულაციის ისტორიული განვითარების შესახებ დასაბუთებული ჰიპოთეზის ჩამოყალიბება, სხვადასხვა ელემენტარული ფაქტორის მოქმედების დადგენა.
მაგალითისათვის განვიხილოთ ადამიანში სისხლის MN ჯგუფი. როგორც გენეტიკიდან ვიცით, არსებობს სამი ჯგუფი M,N და MN, მათ ერთი გენის ორი LM და LN ალელი განსაზღვრავს. თბილისში პოპულაციის 500 ინდივიდზე ჩატარებული ანალიზისას, 198-ს აღმოაჩნდა M ჯგუფის სისხლი, 230-ს - MN, ხოლო 72-ს - N. სისხლის ჯგუფის შესაბამისად გენოტიპის სიხშირეს ადგენენ თითოეული მაჩვენებლის საერთო რიცხვთან თანაფარდობით. M ჯგუფის სიხშირე ტოლია 198/500=0,396, შესაბამისად MN-ის - 0,460, N-ის - 0,144. ვინაიდან 500 ადამიანი შემთხვევით შერჩეული ერთობლიობაა, მიღებული შედეგი შეიძლება განვიხილოთ, როგორც მთელი პოპულაციისათვის დამახასიათებელი ნიშანი. ალელთა სიხშირის განსაზღვრა გენოტიპური კლასების ანალიზით ხდება: LMLM გენოტიპის ინდივიდს ორი LM ალელი გააჩნია, LMLN - თითო LM და LN ალელი, ხოლო LNLN გენოტიპისას ორი LN ალელი. ერთობლიობაში LM ალელის რაოდენობა ტოლია (198X2)+230=626; LN ალელის რაოდენობა კი (72X2)+230=374. ალელთა რაოდენობა ორჯერ აღემატება შესასწავლ პირთა რაოდენობას, ვინაიდან თითოეულს ორი ალელი აქვს 500X2=1000, ამრიგად, LM ალელის სიხშირე შეადგენს 626/1000=0,626; LN ალელის სიხშირე კი 374/1000=0,374.
როდესაც ჰომო- და ჰეტეროზიგოტური კლასები ერთიმეორისაგან ფენოტიპურად განსხვავდებიან (კოდომინანტობა, არასრული დომინანტობა და სხვ.) ფენის მიხედვით, ადვილია შესაბამისი გენოტიპის დადგენა. ვინაიდან სრული დომინანტობისას დომინანტ ჰომოზიგოტს (AA) და ჰეტეროზიგოტს (A) ერთიანი ფენი გააჩნიათ, ცალკეული ალელის სიხშირისა და ცალკეული გენოტიპური კლასების თანაფარდობის განსაზღვრა ფენოტიპური კლასების მიხედვით შეუძლებელია. ამ შემთხვევაში რეცესიული და დომინანტური ალელის სიხშირესა და შესაბამისი გენოტიპური კლასების თანაფარდობას ჰარდი-ვაინბერგის ფორმულით საზღვრავენ.
|
|
6.3 ჰარდი-ვაინბერგის კანონი |
▲ზევით დაბრუნება |
1908 წელს ერთიმეორისგან დამოუკიდებლად ინგლისელმა მათემატიკოსმა ჯ. ჰარდიმ და გერმანელმა ექიმმა ვ. ვაინბერგმა დაადგინეს კანონზომიერება, რომელსაც შემდგომ ჰარდი-ვაინბერგის კანონი უწოდეს (შევნიშნავთ, რომ 1903 წ. ამერიკელმა გენეტიკოსმა ვ. კასტლმა და ინგლისელმა ბიომეტრისტმა კ. პირსონმა მსგავსი პრინციპი უკვე ჩამოაყალიბეს, მაგრამ ეს შრომები შეუმჩნეველი დარჩა). ამ კანონით მტკიცდება, რომ მემკვიდრეობა პოპულაციაში არ ცვლის ალელთა სიხშირეს. ამრიგად, პოპულაციებში (თუ მასზე გარემო ფაქტორები არ მოქმედებს) დომინანტური ჰომოზიგოტების (AA), დომინანტური ჰეტეროზიგოტებისა (A) და რეცესეული ჰომოზიგოტების (AA) სიხშირე თაობათა მანძილზე უცვლელია, ე.ი. დინამიურ წონასწორობას ინარჩუნებს. თუ დომინანტი A ალელის სიხშირეს, აღვნიშნავთ P-თი, რეცესიული A ალელისას კი Q-თი, მაშინ პოპულაციაში მოცემული ლოკუსის მხოლოდ ორი ალელის არსებობის შემთხვევაში წარმოიქმნება ტოლობა: PA+QA=1. ამ შემთხვევაში გენოტიპთა თანაფარდობა იქნება: (PA+QA)2=P2AA+2QAA+Q2AA, რაშიც ადვილად დავრწმუნდებით თუ პენეტის ცხრილს გამოვიყენებთ:
Q/ W |
PA |
QA |
PA |
P2AA |
PQAA |
QA |
PQAA |
Q2AA |
დავუშვათ, რომ პოპულაციაში ერთი გენის ორი განსხვავებული ალელის მქონე ჰომოზიგოტი ინდივიდები AA და აა თანაბარი რაოდენობითაა და თავისუფლად შეეჯვარებიან ერთმანეთს (პანმიქსია). მაშასადამე A და ა ალელები გვხვდება თანაბარი - 0,5 სიხშირით. ამ შემთხვევაში F1-ში გენოტიპურ კლასთა სიხშირე იქნება:
Q/ W |
0,5 |
0,5A |
0,5A |
0,25AA |
0,25AA |
0,5A |
0,25AA |
0,25AA |
ამრიგად, 0,25AA+0,50AA+0,25AA=1. ეს კი გვიჩვენებს, რომ პოპულაციის არსებულ თაობაში ჰომოზიგოტები AA და აა წარმოიქმნება შესაბამისად 0,25 სიხშირით, ხოლო ჰეტეროზიგოტები AA-0,50 სიხშირით. შემდგომ თაობაშიც ალელებისა და გენოტიპების იგივე სიხშირე შენარჩუნდება. დომინანტი A ალელის მქონე გამეტების სიხშირეა 0,5 (0,25 AA ჰომოზიგოტებიდან +0,25 AA, ჰეტეროზიგოტებიდან), ასევე რეცესიული A ალელის მქონე გამეტების სიხშირე 0,5-ია (0,25 AA ჰომოზიგოტებიდან + 0,25 AA ჰეტეროზიგოტებიდან). თუ ეს წონასწორობა პოპულაციაზე რაიმე ფაქტორის მოქმედებით არ იქნება დარღვეული, იგივე თანაფარდობა შენარჩუნდება მომდევნო თაობებში.
ბუნებრივ პოპულაციებში ჰომოზიგოტი AA და AA კლასები განსხვავებული სიხშირით გვხვდება. ამის შესაბამისად დომინანტური (A) და რეცესიული (ა) ალელების სიხშირეც განსხვავებულია. დავუშვათ, რომ პოპულაციაში დომინანტური A ალელის სიხშირე 0,3-ია, ხოლო რეცესიული A ალელის კი 0,7. მაშინ გენოტიპური კლასების ასეთ სიხშირეს მივიღებთ:
Q/ W |
0,3A |
0,7ა |
0,3A |
0,09AA |
0,21AA |
0,7A |
0,21AA |
0,49AA |
მაშასადამე 0,09AA+0,42A+0,49AA. ამრიგად, შთამომავლობაში ყოველი 100 ზიგოტიდან 9 ჰომოზიგოტი AA, 49 ჰომოზიგოტი AA და 42 ჰეტეროზიგოტი AA იქნება. მომდევნო თაობაში გენოტიპური კლასები იმავე სიხშირით წარმოიქმნება. ვინაიდან დომინანტი A ალელის მქონე გამეტები წარმოიქმნება 0,3 სიხშირით (0,09 AA ჰომოზიგოტებიდან + 0,21AA ჰეტეროზიგოტებიდან), ხოლო A ალელის მქონე გამეტები კი 0,7(0,49AAჰომოზიგოტებიდან + 0,21A ჰეტეროზიგოტებიდან).
თუ მოცემული გენის ერთი ალელის სიხშირეს აღვნიშნავთ ქ სიმბოლოთი, მაშინ ამავე გენის ალტერნატიული ალელის სიხშირე შეიძლება გამოისახოს როგორც 1-ქ. თუ ალელების აღმნიშვნელ სიმბოლოებს პენეტის ცხრილში ჩავსვამთ, მაშინ იგი ასეთ სახეს მიიღებს:
Q / W |
Q |
(1-Q) |
Q |
Q2 |
ქ(1-Q) |
(1-Q) |
Q(1-Q) |
(1-Q)(1-Q) |
იგი შეჯამებული სახით შეიძლება ასე წარმოვიდგინოთ Q2+2QX(1-Q)+(1-Q)2 ან [Q+(1-Q)]2; ჰარდი-ვაინბერგის ფორმულით შესაძლებელი ხდება პოპულაციაში ალელებისა და გენოტიპური კლასების თანაფარდობის განსაზღვრა იმ შემთხვევაშიც კი, როდესაც ყველა გენოტიპი არაა იდენტიფიცირებული. მაგ., ადამიანში ალბინიზმს იწვევს რეცესიული გენი. იგი პოპულაციაში დაბალი სიხშირით გვხვდება. თუ ნორმალური პიგმენტაციის განმსაზღვრელი გენის დომინანტურ ალელს აღვნიშნავთ A-თი, ხოლო ალბინიზმის გამომწვევ რეცესიულ ალელს კი A-თი, მაშინ ნორმალური პიგმენტაციის მქონე ადამიანების გენოტიპები იქნება AA ან A, ხოლო ალბინოსის - AA. თუ რომელიმე პოპულაციაში ალბინიზმის სიხშირეა 1:10000, მაშინ ჰომოზიტური (AA) კლასის სიხშირე იქნება Q2=0,0001; Q=0,01, A ალელის სიხშირე კი 0,01-ის ტოლია. A ალელის სიხშირე კი იქნება 1-Q=1-0,01=0,99; ნორმალური დომინანტური ჰომოზიგოტი (AA) კლასის სიხშირე იქნება P2=0,992=0,98; ჰეტეროზიგოტული AA კლასების სიხშირე 2PQ=2X0,99X0,01=0,0198, ე.ი. დაახლოებით 0,02 ტოლია. მაშასადამე, რეცესიული ალელი ჰეტეროზიგოტულ მდგომარეობაში 100-ჯერ მეტია, ვიდრე ჰომოზიგოტურში. ამრიგად, თუ ვიცით პოპულაციაში მხოლოდ რეცესიული ჰომოზიგოტების (AA) სიხშირე, საკმარისია ამ სიდიდიდან კვადრატული ფესვის ამოღება, რომ დავადგინოთ რეცესიული ალელის სიხშირე (ქ). დომინანტი ალელის სიხშირე კი შეადგენს P=1-Q-ს. ამ ხერხით პოპულაციაში A და ა ალელების სიხშირის განსაზღვრის შემდეგ ადვილია პოპულაციის ყველა გენოტიპური კლასების სიხშირის დადგენა.
ამრიგად, ჰარდი-ვაინბერგის კანონის თანახმად, უსასრულოდ დიდი პანმიქტიური პოპულაციები, რომლებზეც არ მოქმედებს გარემო ფაქტორები, დინამიურ წონასწორობაშია, ანუ ალელთა და გენოტიპთა სიხშირე მასში თაობათა მანძილზე უცვლელია. ეს კანონი მოქმედებს იმ შემთხვევაში, თუ: 1. პოპულაცია არის მრავალრიცხოვანი; 2. შეჯვარება შემთხვევითი ხასიათისაა; 3. არ წარმოიქმნება ახალი მუტაციები; 4. ყველა გენოტიპი ერთნაირი სიცოცხლისუნარიანია და, მაშასადამე, გადარჩევა არ მიმდინარეობს; 5. თაობები ერთმანეთს არ ფარავენ; 6. არ მიმდინარეობს ემიგრაცია და იმიგრაცია (გენთა ნაკადი), ე.ი. პოპულაციათა შორის გენეტიკური მასალის მიმოცვლა არ ხდება.
თუ დაცულია ზემოთ ჩამოთვლილი პირობები, მაშინ სხვადასხვა ალელების სიხშირე თაობათა მანძილზე (ე.ი. ხანგრძლივად) უცვლელი რჩება, პოპულაციაში გენეტიკური წონასწორობა მყარდება და მასში არავითარი ევოლუციური ცვლილება არ მიმდინარეობს. ბუნებაში ასეთი პოპულაციები მეტად იშვიათია. მეტწილად პოპულაციის გენეტიკური სტრუქტურის დინამიკაზე მოქმედ ფაქტორთა გავლენით თანაფარდობა ირღვევა.
|
|
6.4 გენეტიკური ჰეტეროგენულობა |
▲ზევით დაბრუნება |
ბუნებრივი პოპულაციების გენეტიკური ჰეტეროგენულობის მნიშვნელობას ყურადღება ს.ჩეტვერიკოვმა (1926) მიაქცია. მან ჰარდი-ვაინბერგის კანონზე დაყრდნობით, თეორიულად შეისწავლა რეალური სიტუაცია და გამოთქვა მოსაზრება, რომ პოპულაციაში მეტწილად რეცესიული მუტაციები წარმოიქმნება. ისინი პოპულაციაში ჰეტეროზიგოტულ მდგომარეობაში ინახება და არ ცვლის მის გარეგან სახეს (ფენოტიპს). ამის გამო პოპულაცია როგორც „ღრუბელი“, ისეა გაჟღენთილი მუტაციებით. ნებისმიერი პოპულაცია გენეტიკურად ჰეტეროგენულია.
XXს. 50-იან წლებში პოპულაციის გენეტიკური სტრუქტურის შესახებ ორი ალტერნატიული ჰიპოთეზა წარმოადგინეს (კლასიკური და ბალანსური მოდელი). კლასიკური მოდელის ავტორია ნობელის პრემიის ლაურეატი ჰ. მელერი, ამ შეხედულების მიხედვით, პოპულაციის გენეტიკური ცვალებადობის დონე მეტად უმნიშვნელოა. ბუნებრივი პოპულაცია ძირითადად დომინანტი ჰომოზიგოტი ალელებისგან შედგება. დომინანტი ალელების სიხშირე თითქმის ერთს უტოლდება. ამასთანავე, პოპულაციას გენოფონდში მცირე რაოდენობით მოეპოვება სპონტანურად ინდუცირებული საზიანო რეცესიული მუტაციები. ბუნებრივი გადარჩევით მათი სიხშირე უმნიშვნელო დონეზეა შენარჩუნებული. ამ მოდელის მიხედვით ტიპური ინდივიდის თითქმის ყველა ლოკუსი ფლობს ველური ტიპის დომინანტ ალელებს. დასაშვებია, რომ ლოკუსთა მცირე რაოდენობას მუტანტური რეცესიული ალელი გააჩნდეს ჰეტეროზიგოტულ მდგომარეობაში. „ნორმალური“ ანუ იდეალური გენოტიპის მფლობელ ინდივიდში ყველა ალელი ველური ტიპისაა და ჰომოზიგოტურია. პოპულაციის ევოლუციური მიმდინარეობა იშვიათად ინდუცირებულ სასარგებლო ალელების გადარჩევას ეფუძნება. ბუნებრივი გადარჩევის მოქმედებით მათი სიხშირე პოპულაციაში იზრდება.

სურ. 4.1. პოპულაციის გენეტიკური სტრუქტურის ორი: კლასიკური და ბალანსური მოდელი. მოწოდებულია სამი ტიპური წევრის ჰიპოთეტური გენოტიპი. ლოკუსები აღნიშნულია ლათინური ანბანით. განსხვავებული ალელები მოცემულია ციფრებით (ინდექსები). კლასიკურ მოდელში ველური ტიპის გენი „+“ ნიშნითაა აღნიშნული და მხოლოდ ზოგიერთი ლოკუსებია ჰეტეროზიგოტულ მდგომარეობაში. ბალანსურ მოდელში მრავალი ლოკუსი ჰეტეროზიგოტულ მდგომარეობაშია (ფ. აიალა, ჯ. კაიგერი, 1988).
ბალანსური მოდელი შემოთავაზებულია გამოჩენილი ამერიკელი ევოლუციონისტისა და გენეტიკოსის ფ. დობჟანსკის მიერ, რომლის მიხედვით, ბუნებრივ პოპულაციებში ნიშანდობლივია ცვალებადობის მაღალი დონე. მათი გენეტიკური სტრუქტურა პოლიმორფულია. ცალკეული გენისათვის არ არსებობს ველური ტიპის კონკრეტული სახის ალელი. გენთა უმეტესი ნაწილი ალელთა სერიისგან შედგება, რომლებიც პოპულაციებში განსხვავებული სიხშირით გვხვდება. პოპულაცია განსხვავებულ გენთა ერთობლიობას წარმოადგენს. ბალანსური მოდელის თანახმად, „ნორმალური“ ანუ იდეალური გენოტიპი არ არსებობს. ევოლუციური ცვლილება ბალანსურ დამოკიდებულებაში მყოფი გენებისა და ალელების გადარჩევაზეა დაფუძნებული. ამის შედეგად ალელთა ტიპები და სიხშირე თანდათანობით იცვლება. ორივე მოდელის მიხედვით ახლად წარმოქმნილი სპონტანური მუტაციების უმეტესი ნაწილი ორგანიზმისთვის საზიანოა. ბუნებრივი გადარჩევით საზიანო მუტაციები ელიმირდებიან ან შენარჩუნებულია მეტად დაბალი სიხშირით. ევოლუციის პროცესში ისინი მეორეხარისხოვან ნეგატიურ როლს ასრულებენ (იხ. სურ. 4.1).
ბუნებრივი პოპულაციებისათვის ნიშანდობლივია გენეტიკური ცვალებადობა. პოპულაციის ცვალებადობის დონეს ჰიბრიდული შთამომავლობის ანალიზით საზღვრავენ. ინბრიდინგით (იხ. §7.4) იზრდება შთამომავლობაში ჰომოზიგოტების (მათ შორის რეცესიული ჰომოზიგოტების) წარმოქმნის ალბათობა. ინბრიდინგის მეშვეობით შეძლეს ფარულ მდგომარეობაში მყოფი რეცესიული მორფოლოგიური მუტაციების გამოვლენა და მათი სიხშირის განსაზღვრა. კერძოდ, დადგენილია, რომ გენეტიკის კლასიკური ობიექტის - დროზოფილას ნებისმიერი პოპულაციის თითქმის ყველა ინდივიდი ფარულად ფლობს რეცესიულ ალელებს, რომლებიც ჰომოზიგოტურ მდგომრეობაში იწვევენ ნორმალური ფენოტიპიდან გადახრას. ასევე გამოირკვა, რომ მრავალ მცენარეს ჰეტეროზიგოტულ მდგომარეობაში გააჩნია რეცესიული ალელები, რომლებიც ჰომოზიგოტურ მდგომარეობაში არღვევენ ქლოროფილის ნორმალურ სინთეზს ან ამ პროცესის სრულ ბლოკირებას იწვევენ.
გენეტიკური ცვალებადობის შესახებ სარწმუნო არგუმენტებია მოპოვებული ხელოვნური გადარჩევის ექსპერიმენტებით. გამოვლენილია ბიოლოგიური მასალის მაღალი ჰეტეროზიგოტულობა სამეურნეო მნიშვნელობის ნიშნების მიხედვით ხელოვნური გადარჩევისას. მაგ., თეთრ ლენჰორნში გადარჩევით (1933-1965 წწ.) გაზრდილია წლიური კვერცხმდებლობა 125,6-დან 249,6 ცალამდე. სიმინდის ორ განსხვავებულ ხაზში ჩატარებული ხელოვნური გადარჩევით შეცვლილია მარცვალში ცილის შემცველობა. პირველ ხაზში ცილის შემცველობა გაზრდილია 10,9%25-დან 19,4%25-მდე, მეორეში კი შემცირებულია 10,9%25-დან 4,9%25-მდე. ხელოვნური გადარჩევით ადამიანისათვის საინტერესო სამეურნეო ნიშანი შეცვლილია მრავალ კულტურულ მცენარესა და შინაურ ცხოველში, ასევეა საწარმოო მნიშვნელობის მიკროორგანიზმებში. ხელოვნური გადარჩევა წარმატებული აღმოჩნდა პრაქტიკულად ყველა შემთხვევაში, რაც მიუთითებს, რომ ხელოვნური პოპულაციებისათვის (ჯიშები) ნიშანდობლივია გენეტიკური ცვალებადობის მაღალი დონე. ზემოთ ჩამოთვლილი არგუმენტებით დასტურდება ბალანსური ჰიპოთეზის მართებულობა.
მხოლოდ XXს. 60-იანი წლებში გახდა შესაძლებელი პოპულაციაში ჰეტეროზიგოტულობისა და პოლიმორფულობის უშუალო დონის განსაზღვრა და სარწმუნო არგუმენტების მოპოვება. მოლეკულურ ბიოლოგიაში შემუშავებული მეთოდებით ბუნებრივ პოპულაციებში შესწავლილია გენეტიკური ჰეტეროგენულობა. ჩაატარეს სხვადასხვა პოპულაციებში მრავალი გენისა და მისი პროდუქტის - ფერმენტის მოლეკულური ანალიზი. გამოვლენილია ბუნებრივ პოპულაციებში გენეტიკური ჰეტეროგენულობის მაღალი დონე. ცვალებადობის დონის შეფასებას ორი სიდიდით ახდენენ: გენეტიკური პოლიმორფიზმით (P) და ჰეტეროგენულობით (H). პოპულაციის პოლიმორფულობას პოლიმორფული ლოკუსების საერთო რაოდენობაში მათი წილით გამოხატავენ. მაგალითად, კალიფორნიულ ზღვის ჭიაში (Pჰორონოპსის ვირიდის) 30 შესწავლილი ლოკუსიდან 18 ლოკუსს აღმოაჩნდა ვარიანტები, 12 ლოკუსში ვარიანტები არ აღინიშნებოდა. ამ შემთხვევაში P=18:30=0,60. ამავე სახეობის ოთხ განსხვავებულ პოპულაციაში ეს სიდიდე აღმოჩნდა, 0,60; 0,50; 0,53; და 0,47. სახეობის საშუალო პოლიმორფულობა ტოლია (0,60+0,50+0,53+0,47)/4=0,525.
გენეტიკური ცვალებადობის ზუსტ მახასიათებელს პოპულაციის ჰეტეროზიგოტულობის განსაზღვრა წარმოადგენს. კონკრეტული ჰეტეროზიგოტულობა გამოისახება მაჩვენებლით: ჰეტეროზიგოტებისა და პოპულაციაში შესწავლილ ინდივიდთა საერთო რაოდენობასთან თანაფარდობით. თავდაპირველად საზღვრავენ კონკრეტული ლოკუსის ჰეტეროზიგოტულობას, ხოლო შემდეგ ყველა გაანალიზებული ლოკუსების მაჩვენებელს ასაშუალოებენ. დავუშვათ პოპულაციაში ოთხი ლოკუსის კონკრეტული ლოკუსების ჰეტეროზიგოტულობის მაჩვენებელია: 0,25; 0,42; 0,09 და 0,01. მოცემული ლოკუსების საშუალო ჰეტეროზიგოტულობა ტოლია (0,25+0,42+0,09+0,01)/4=0,19. ამრიგად პოპულაციის ჰეტეროზიგოტულობა 19%25-ის ტოლია. თუ ერთი სახეობის რამდენიმე პოპულაციაა გაანალიზებული ერთი და იმავე ლოკუსის მიხედვით, მაშინ შესაძლებელია სახეობის საშუალო ჰეტეროზიგოტულობის დადგენა. დავუშავათ ოთხი პოპულაციის ჰეტეროზიგოტულობა შეადგენს: 0,19; 0,15;0,13 და 0,17, მაშინ სახეობის საშუალო ჰეტეროზიგოტულობა ტოლია (0,19+0,15+0,13+0,17)/4=0,16.
მრავალი სახეობისათვის განსაზღვრულია საშუალო პოლიმორფიზმი (P და საშუალო ჰეტეროგენულობა (H. გამოირკვა, რომ უხერხემლო ცხოველებს (H 13,4%25) გენეტიკური ცვალებადობის გაცილებით მაღალი დონე ახასიათებს ვიდრე ხერხემლიანებს (H=6,0%25). ალოგამური მცენარეები (H=19%25) გაცილებით ცვალებადია ვიდრე ავტოგამური (H=6%25). ცილების გელ-ელექტროფორეზის შედეგად მიღებული მონაცემების საფუძველზე დადგენილია, რომ ადამიანის პოპულაციების საშუალო ჰეტეროზიგოტულობა 6,7%25-ს შეადგენს. საინტერესოა განვსაზღროთ თუ რაზე მიგვანიშნებს ეს სიდიდე.
თუ დავუშვებთ, რომ ადამიანს დაახლოებით 30000 სტრუქტურული გენი აქვს (რეალურთან შედარებით ეს მაჩვენებელი რამდენადმე შემცირებულია), მაშინ საშუალოდ თითოეულ ადამიანს ჰეტეროზიგოტულ მდგომარეობაში 30000x0,067=2010 გენი ექნება. პოტენციურად ასეთ პოლიჰეტეროზიგოტაში 22010“1065 განსხვავებული ტიპის გამეტები უნდა წარმოიქმნას. ბუნებრივია გამეტთა რაოდენობის ასეთი პოტენციური კომბინაცია არა მარტო ცალკეულ ადამიანში, არამედ კაცობრიობის მთელი არსებობის მანძილზე არასოდეს არ რეალიზირდება. შედარებისათვის აღვნიშნავთ, ფიზიკოსების შეფასებით სამყარო 1076 რაოდენობის პროტონებისა და ნეიტრონებისგან შედგება. აქედან ცხადია, რომ ბუნებრივ ჰეტეროზიგოტულობაზე დამყარებული კომბინაციური ცვალებადობის პოტენციური შესაძლებლობებით, ბუნებრივი გადარჩევისათვის კოლოსალური რეზერვი იქმნება. ამრიგად, მუტაციური პროცესითა და შეჯვარებით შენარჩუნებული გენეტიკური ჰეტეროგენულობა პოპულაციას საშუალებას აძლევს საარსებო პირობებთან შეგუებისათვის გამოიყენოს როგორც ახლად ინდუცირებული, ისე ადრე წარმოქმნილი, პოპულაციაში ჰეტეროზიგოტულ მდგომარეობაში არსებული მუტაციები. ამის მიხედვით შეიძლება ითქვას, რომ პოპულაციის ჰეტეროგენულობა უზრუნველყოფს მემკვიდრული ცვალებადობის ე.წ. „სამობილიზაციო რეზერვის“ არსებობას (შმალგაუზენი, 1969).
|
|
6.5 პოპულაციის გენეტიკური ერთიანობა |
▲ზევით დაბრუნება |
ჰეტეროგენულობის გამო ნებისმიერი პოპულაცია რთულ გენეტიკურ სისტემას წარმოადგენს, იგი დინამიურ წონასწორობაში იმყოფება. პოპულაცია ინდივიდთა ის უმცირესი ერთობლიობაა, რომელიც ხანგრძლივი დროის განმავლობაში (თაობათა მანძილზე) არსებობის უნარს ინარჩუნებს. თავისუფალი შეჯვარების შედეგად მრავალი მუტაცია (მათ შორის ისეთიც, რომლებიც ორგანიზმის ცხოველქმედებას აქვეითებს) ჰომოზიგოტურ მდგომარეობაში გადადის და ვლინდება კიდეც. მხოლოდ გარკვეული რიცხოვნობის ბუნებრივ პოპულაციებს აქვს უნარი საჭირო დონეზე შეინარჩუნოს გენეტიკური სისტემის ნაირგვარობა და მთლიანობა.
პოპულაციის გენეტიკაში მოპოვებული მონაცემების განზოგადების საფუძველზე გამოტანილია მნიშვნელოვანი დასკვნა, კერძოდ დებულება პოპულაციის გენეტიკური ერთიანობის შესახებ: ნებისმიერი პოპულაცია მისი შემადგენელი ინდივიდების ჰეტეროგენულობის მიუხედავად დინამიურ წონასწორობაში მყოფ რთულ გენეტიურ სისტემას წარმოადგენს.
ამრიგად, პოპულაციის გენეტიკური მახასიათებელია: მუდმივი გენეტიკური ჰეტეროგენულობა, შიდაგენეტიკური ერთიანობა, ცალკეული ალელებისა და გენოტიპების დინამიკური წონასწორობა. ეს თავისებურებანი განსაზღვრავენ პოპულაციის, როგორც ევოლუციის ელემენტარული ერთეულის, ორგანიზაციას.
|
|
7 §5. ევოლუციის გენეტიკური საფუძვლები |
▲ზევით დაბრუნება |
ბიოლოგიური ევოლუცია არის ბიოლოგიური ფორმების გენეტიკური სტრუქტურის ცვლილება და დიფერენცირება დროში. გენეტიკური ცვალებადობის პრობლემას ცენტრალური ადგილი უკავია ევოლუციურ თეორიაში. ორგანიზმის ნიშნებისა და თვისებების ევოლუციურ გარდაქმნას განაპირობებს გენოტიპში მომხდარი ცვლილებები. მიკროევოლუციის მექანიზმების შესასწავლად საჭიროა პოპულაციებში გენეტიკური ცვლილებების გამომწვევი წყაროების გამოვლენა და მათი ანალიზი. კლასიკური ფორმების გარდა თანამედროვე ეტაპზე გამოვლენილია არაკანონიკური ცვალებადობის ფორმები. დგინდება თითოეული მათგანის მნიშვნელობა ევოლუციის პროცესის მიმდინარეობაში.
ცვალებადობა არის ცოცხალი სისტემის ზოგადი თვისება, რის შედეგადაც პოპულაციის შემადგენელი ორგანიზმები ნაირგვარი ფორმით არსებობს. ცვალებადობა გამოხატავს ორგანულ ფორმათა უწყვეტობას, რომლის დროსაც შთამომავლობაში განსხვავებული სტრუქტურისა და ფუნქციის ვარიანტები ჩნდება. ცვალებადობას მოლეკულურ დონეზე საფუძვლად უდევს დნმ-ის კონვარიანტული რედუპლიკაცია. გენეტიკური ცვალებადობის ზოგადი მიზეზები მეტად ნაირგვარია. ჯერ კიდევ ჩ. დარვინი აღნიშნავდა, რომ ერთი წყვილი მშობლის შთამომავლები საკმაოდ განსხვავდებიან. „გამოცდილ ნატურალისტსაც კი განაცვიფრებდა ორგანიზმთა ყველაზე მნიშვნელოვანი ნაწილის ცვალებადობის მრავალრიცხოვანი შემთხვევა“. შემდგომში გენეტიკის განვითარებამ გამოავლინა ცვალებადობის მიზეზები.
გამოყოფენ ცვალებადობის ორ ძირითად ფორმას: ფენოტიპურს (არამემკვიდრულს) და გენოტიპურს (მემკვიდრულს). ჩ. დარვინი ნაშრომში „სახეობათა წარმოშობა...“ (1859) განასხვავებდა ცვალებადობის ორ ძირითად ფორმას - განსაზღვრულს და განუსაზღვრელს. ეს კლასიფიკაცია შეესაბამება თანამედროვე დაყოფას - არამემკვიდრულ და მემკვიდრულ ცვალებადობას.
მემკვიდრული ცვალებადობა განისაზღვრება გენოტიპით და გადაეცემა შთამომავლობას, ხოლო არამემკვიდრული ცვალებადობა განისაზღვრება გენოტიპისა და გარემოს ურთიერთქმედებით. იგი სქესობრივი გამრავლებისას შთამომავლობას არ გადაეცემა. ამრიგად მემკვიდრული ცვალებადობა გენოტიპის ცვლილებაა, ხოლო არამემკვიდრული - მხოლოდ ფენოტიპის. ცვალებადობა მემკვიდრეობასთან ერთად უნივერსალური თვისებაა, რომელსაც ფლობს ყველა ცოცხალი ორგანიზმი და იძლევა მასალას ევოლუციისთვის.
|
|
7.1 5.1. ფენოტიპური ცვალებადობა |
▲ზევით დაბრუნება |
არამემკვიდრული ცვალებადობის დროს გენოტიპი უცვლელია, იცვლება მხოლოდ ფენოტიპი. ორგანიზმების ინდივიდური განვითარების პროცესში ვხვდებით მორფოლოგიურ, ფიზიოლოგიურ, ბიოქიმიურ და ორგანიზმის სხვა ნიშან-თვისებათა ცვლილებებს. ამდაგვარი ცვლილებების დრო და თანამიმდევრობა ონტოგენეზში გენოტიპით მკაცრადაა განსაზღვრული. ცვალებადობის ამ ფორმას ასაკობრივს ანუ ონტოგენეზურს უწოდებენ. ცვალებადობის ეს ფორმა თვალნათლივ ვლინდება მეტამორფოზული განვითარების ცხოველთა (მწერები, თასმისებრი ჭიები, უკუდოსნაირი ამფიბიები და მისთ.) სასიცოცხლო ციკლში. ასაკობრივი ცვლილების მიუხედავად ინდივიდს ყოველთვის ერთნაირი გენოტიპი აქვს.
ლარვულ განვითარებას ადაპტური მნიშვნელობა გააჩნია, რომელიც ევოლუციის პროცესში ჩამოყალიბდა. მოძრავ ფორმებში (მაგ., მწერები, უკუდოსნაირი ამფიბიები) ლარვები და ზრდასრული ფორმები განსხვავებულ გარემოში ცხოვრობენ და სხვადასხვაგვარ საკვებს იყენებენ, რის გამოც ერთმანეთს კონკურენციას არ უწევენ. მცირედ მოძრავ ან მჯდომარე ცხოველებში (მარჯნები, ორსაგდულიანები და მისთ.) ლარვული განვითარება ხელს უწყობს მათ განსახლებას და ახალ საბინადრო გარემოს ათვისებას. ზოგიერთ ორგანიზმთა (პარაზიტი ჭიები, ციბრუტელები და მისთ.) ონტოგენეზში შეინიშნება გენეტიკური მასალის (დიმინუცია, დნმ-ის მნიშვნელოვანი ნაწილის დაკარგვა) ცვლილება, რაც მას მემკვიდრულ ცვალებადობასთან აახლოებს.
ერთნაირი გენოტიპის ინდივიდებზე ან ინდივიდთა ჯგუფზე განსხვავებული გარემო პირობების მოქმედებით გამოწვეულ ცვალებადობას მოდიფიკაციურს ანუ პარატიპურს უწოდებენ. მოდიფიკაციური ცვალებადობა მუტაციურის ალტერნატიული ფორმაა. კერძოდ, მუტაცია - გენების რეპროდუქციის პროცესის დარღვევის შედეგია, ხოლო მოდიფიკაცია - გენის ექსპრესიის ცვლილების შედეგი.
ორგანიზმის ნებისმიერი ნიშან-თვისება მემკვიდრულადაა დეტერმინირებული. არამემკვიდრული ნიშან-თვისება არ არსებობს. მემკვიდრეობით გადაეცემა არა ნიშან-თვისება, არამედ მემკვიდრული ინფორმაცია, რომელიც განსაზღვრავს კონკრეტული ნიშნისა და თვისების შესაძლო განვითარებას. ამრიგად გენოტიპით პროგრამირდება მხოლოდ ნიშან-თვისების ჩამოყალიბების ზოგადი მიმართულება, ხოლო გარემო პირობები მნიშვნელოვან გავლენას ახდენს მათ კონკრეტულ რეალიზაციაზე. გენოტიპის ფენოტიპური რეალიზაციისათვის აუცილებელია კონკრეტული გარემო პირობები. ნიშნის გამოვლენის ხარისხი შეიძლება გარემო პირობებზე დამოკიდებულების მიხედვით განსხვავებული იყოს. იგი გამოწვეულია იმით, რომ ყოველ გენოტიპს აქვს ცალკეული ნიშნის რეაქციის ნორმის განსაზღვრის უნარი. რეაქციის ნორმად მიჩნეულია ის ფარგლები, რომელშიაც შესაძლებელია მოხდეს ფენოტიპის ფლუქტუაცია გენოტიპის ცვლილების გარეშე. მიუხედავად იმისა, რომ ფენოტიპი მემკვიდრეობითაა განსაზღვრული, ის არ მემკვიდრეობს. მემკვიდრულია მხოლოდ ნებისმიერი კონკრეტული გენოტიპის გარემოს ზემოქმედებით გამოწვეული რეაქციის ნორმა.
გამოყოფენ მოდიფიკაციური ცვალებადობის რამდენიმე ფორმას, რომელთაგანაც ევოლუციის პროცესში მნიშვნელოვანია ადაპტური მოდიფიკაცია. იგი ბუნებაში ფართოდაა გავრცელებული. მას დიდი შეგუებითი მნიშვნელობა აქვს. ორგანიზმები ადაპტური მოდიფიკაციით ადვილად ეგუებიან შეცვლილ პირობებს და დიდი რაოდენობით ტოვებენ შთამომავლობას. მაგალითად, წყლის მცენარე ისარას (შაგიტტარა საგიტტარიაფოლია) ფოთლის ფორმა მკვეთრად ვარირებს განვითარების პირობების მიხედვით: წყალში განვითარებული ფოთლის ფირფიტა ლენტისებურია, წყლის ზედაპირზე - ოვალური, წყლის გარეშე - ისრის ბუნიკის ფორმისა. ამდაგვარი ვარიაცია სინათლის მოქმედების შედეგია (იხ. სურ. 5.1). ჩრდილის ამტან მცენარეებს გააჩნიათ: დიდი ფოთლის ფირფიტა, მესრისებურ პარენქიმაში რამდენიმე შრედ განლაგებული უჯრედები, ქლოროპლასტების დიდი რაოდენობა, რითაც აქტივიზირებულია ფოტოსინთეზი. ბეწვიანი ნადირის ბალანი დაბალი ტემპერატურისას (შემოდგომა-ზამთარი) ხშირი და გრძელია. ზოგიერთ მწერს, თევზს, ამფიბიას, ქვეწარმავალს და რიგ სხვა ცხოველს გარემო ფონის შესაბამისად ეცვლება სხეულის შეფერილობა. ადაპტური მოდიფიკაციის შედეგია ზოგიერთ ცხოველში შეფერილობის სეზონური ცვლა ე.წ. სეზონური დიმორფიზმი: მაგ., თეთრი გნოლი, თეთრი კურდღელი, ყარსაღი, ზამთარში თეთრია, ზაფხულში კი არა. ნამზეურობა ადამიანში ადაპტური ცვლილებაა, რაც მას იცავს მზის რადიაციის მავნე ზემოქმედებისაგან.
სურ. 5.1. მცენარე ისარა. გააჩნია ფენოტიპურად განსხვავებული სამი სახის ფოთლები: წყალქვეშა, ზედაპირული და წყლის ზედა (ს. ინგე-ვეჩტომივი, 1989).
მობილური გენეტიკური ელემენტების ანალიზით დადგენილია, რომ გარემოს ზემოქმედებაზე უჯრედს, როგორც მთლიან სისტემას შეუძლია ადაპტურად გარდაქმნას გენომი. გარემოს ფლუქტუაციაზე უჯრედი აქტიურად რეაგირებს გენეტიკური სტრუქტურის ცვლილებით. ამდენად, იგი პასიურად არ ელოდება ადაპტური მუტაციის შემთხვევით ინდუქციას. საილუსტრაციოდ შეიძლება რამდენიმე მაგალითი მოვიყვანოთ.
მედიცინაში, კერძოდ ონკოლოგიაში ავთვისებიანი უჯრედების ზრდის შესაჩერებლად ფართოდ იყენებენ უჯრედის გაყოფის ინჰიბიტორს - მეტატრექსატს. ეს შხამი უჯრედში იწვევს ფერმენტ დიჰიდროფოლატრედუქტაზას ინაქტივაციას, რომლის სინთეზს კონკრეტული გენი განსაზღვრავს. უჯრედი სელექციური აგენტის ზემოქმედებას შხამისადმი მდგრადობის განმსაზღვრელი ლოკუსის რაოდენობის თანდათანობითი გაზრდით - ტრანსპოზიციით რეაგირებს.
ძუძუმწოვრების ქსოვილთა კულტურის უჯრედებში ხშირად ფორმირდება ტიპური ქრომოსომები, რომლებიც მეტატრექსატისადმი მდგრადობის განმსაზღვრელი გენის (30-40 ათასი ნ.წ.) მრავალი ასლისაგან შედგება. რის გამოც ამ ნივთიერებისადმი მდგრადობა რამდენიმე ათასით არის გაზრდილი.
კანის წყლულის გამომწვევ შოლტოსან ლეიშმანიას (Leischmania tropica) მეტატრექსატისადმი მდგრადობის მხოლოდ ერთი გენი გააჩნია. ინჰიბიტორის ზემოქმედებით იგი ამპლუცირებს და შხამისადმი მდგრადობა თანდათანობით მატულობს. სელექციური ფაქტორის ზემოქმედებით ხორციელდება გენომის გარდაქმნა და ერთი ობლიგატური გენიდან, ფაკულტატური გენების ჩამოყალიბება. ნივთიერების ხანგრძლივი ზემოქმედებისას გენის შემცველი დნმ-ს ფრაგმენტები ქრომოსომებში ინტეგრირებს. ჩამოყალიბებული გენეტიკური კონსტრუქცია არასტაბილურია. როდესაც გარემოში მეტატრექსატი არ აღინიშნება გენთა რაოდენობა თანდათანობით მცირდება და საწყის დონეს უბრუნდება. ამდაგვარი რეაქცია ნიშანდობლივია ხანგრძლივი მოდიფიკაციისათვის. კერძოდ, გამოწვეული ვარიაცია მასობრივი (ფლობს ყველა ინდივიდი), მემკვიდრეობითი და შექცევადია. გარკვეული დროის შემდეგ მეტატრექსატის განმეორებით ზემოქმედებისას, საწყის ფორმებთან შედარებით გაცილებით სწრაფად ფორმირდება მდგრადი ინდივიდები. ლეიშმანიებს გადატანილ სტრესზე უყალიბდებათ თავისებური ამპლიფიკაციური უჯრედული მეხსიერება. ამრიგად, ადაპტური მოდიფიკაცია არის ორგანიზმის საპასუხო რეაქცია შეცვლილ გარემოზე, რათა წარმატებულად აითვისოს და შეეგუოს საარსებო პირობებს.
გენეტიკური პროცესი, როგორიცაა ტრანსკრიპცია (დნმ-ის მატრიცაზე რნმ-ის სინთეზი) და ტრანსლაცია (ინფორმაციული რნმ-ის მატრიცაზე ცილის სინთეზი) საფუძვლად უდევს ონტოგენეზში გენეტიკური ინფორმაციის რეალიზაციას. ამ პროცესების გარემო ფაქტორების ზემოქმედებით დარღვევა გარკვეული დონით საფუძვლად უდევს მოდიფიკაციურ ცვალებადობას.
|
|
7.2 5.2. მუტაციები - ევოლუციის ელემენტარული მასალა |
▲ზევით დაბრუნება |
ევოლუციის პროცესის შესწავლისას აუცილებელია ევოლუციის ელემენტარული მასალის გამოვლენა და მისთვის ნიშანდობლივი ნიშნების მკაფიოდ ფორმულირება. მემკვიდრული ცვალებადობის ელემენტარული ერთეული უნდა იყოს დაუყოფადი. ე.ი. არ უნდა შეიცავდეს ამავე თვისების მფლობელ სუბერთეულებს. მემკვიდრული ცვალებადობის დისკრეტული ერთეული, რომელიც წარმოადგენს ელემენტარული ევოლუციური პროცესისათვის მასალას უნდა აკმაყოფილებდეს შემდეგ მოთხოვნებს:
1. მემკვიდრული ცვალებადობის მატერიალური ერთეულები მუდმივად და საკმაო რაოდენობით, სპონტანურად უნდა წარმოიქმნან ყველა ცოცხალ ორგანიზმში.
2. ცვალებადობა უნდა შეეხოს ინდივიდის ნებისმიერ, მათ შორის „სასიცოცხლო მნიშვნელობის“ ნიშან-თვისებას. ცვლილება უნდა იწვევდეს საწყისი ფორმიდან სხვადასხვა მიმართულებით გადახრას.
3. წარმოქმნილი ცვლილებები განსხვავებული სიხშირით უნდა გვხვდებოდეს ნებისმიერი ცოცხალი ორგანიზმის ბუნებრივ პოპულაციებში.
4. ცვლილებების ნაწილი უნდა „გავიდეს“ ევოლუციის ისტორიულ არენაზე. მათი მონაწილეობით უნდა ჩამოყალიბდეს უმდაბლესი ტაქსონები.
5. ბუნებრივი პანმიქტური პოპულაციები მემკვიდრული ცვალებადობის ელემენტარული ერთეულების ნაკრებით და კომბინაციით ერთმანეთისგან არ უნდა განსხვავდებოდნენ.
ექსპერიმენტულმა გენეტიკამ გამოავლინა, მთელი სისრულით აღწერა და გააანალიზა ელემენტარული დისკრეტული მემკვიდრული ცვლილებები - მუტაციები. მხოლოდ მუტაციები აკმაყოფილებენ იმ პირობებს, რომელიც წაეყენება ევოლუციის ელემენტარულ მასალას.
მუტაციური ცვალებადობა ცოცხალი სისტემის უნივერსალური თვისებაა. იგი მიმდინარეობს ყველა ცოცხალ სისტემაში ვირუსებიდან მოყოლებული უმაღლეს მცენარეებსა და ცხოველებში, თვით ადამიანის ჩათვლით. მუტაციებს მოქმედების ფართო სპექტრი მოეპოვებათ. იგი ეხება ორგანიზმის ყველა ნიშან-თვისებას. მასთან დაკავშირებულია გენის ახალი ალელების წარმოქმნა, შესაბამისად ახალი ნიშან-თვისების ჩამოყალიბება. ამრიგად მუტაციები არის ელემენტარული მასალა ევოლუციის პროცესისათვის.
მუტაციური ცვალებადობის საყოველთაოდ მიღებული კლასიფიკაცია სადღეისოდ არ არის შემუშავებული. კლასიკური პოზიციებიდან გამომდინარე, მუტაციის ტიპების დაჯგუფება რამდენიმე პრინციპით ხორციელდება: 1. გენომის ცვლილების მიხედვით (გენური ქრომოსომული, გენომური) 2. ჰეტეროზიგოტაში გამოვლენის მიხედვით (დომინანტური, ნახევრადდომინანტური, რეცესიული). 3. მუტაციის გამომწვევი მიზეზების მიხედვით (სპონტანური, ინდუცირებული). 4. ნორმიდან (ველური ტიპიდან) გადახრის მიხედვით (პირდაპირი, შებრუნებული ანუ რევერსია) 5. უჯრედში ლოკალიზაციის მიხედვით (ბირთვული, ციტოპლაზმური) 6. შესაძლო მემკვიდრეობის მიხედვით (გენერაციული, სომატური).
გენური მუტაციები. ამ ტიპის მუტაციის დროს ხდება ცალკეული გენის სტრუქტურის ცვლილება. იგი სხვა ტიპის მუტაციათა შორის ყველაზე მაღალი სიხშირით ინდუცირდება და იწვევს ორგანიზმის მორფოლოგიური, ფიზიოლოგიური და ბიოქიმიური ნიშნების ცვლილებას. გენურ მუტაციებს იწვევს, რეპლიკაციის, ტრანსკრიპციის, ტრანსლაციის, რეპარაციის და რეკომბინაციის პროცესში მომხდარი შეცდომები. გენური მუტაციების დაჯგუფებას ახდენენ: 1. კოდის სტრუქტურის ცვლილების მიხედვით (ტრანზიცია, ტრანსვერსია, ნუკლეოტიდის ჩართვა ან ამოვარდნა). 2. კოდის ინფორმაციის ცვლილების მიხედვით (სეიმსენისი, მისენსი, ნონსენსი, ფრეიმშრიფტი).
ქრომოსომული მუტაციები. ევოლუციის პროცესში იცვლება არა მარტო ქრომოსომის ზომა, არამედ მისი ორგანიზაციაც. ქრომოსომის კონკრეტულმა მონაკვეთმა შეიძლება შეიცვალოს მდებარეობა ან გადავიდეს სრულიად სხვა არაჰომოლოგიურ ქრომოსომაში. ქრომოსომის ზომისა და ორგანიზაციის ცვლილებას ქრომოსომული მუტაციები (სინონიმებია: აბერაცია, ქრომოსომული გარდაქმნა) ეწოდება.
ამრიგად ქრომოსომულმა მუტაციებმა შეიძლება მოიცვას, როგორც ერთი ქრომოსომის მონაკვეთები, ისე სხვადასხვა (არაჰომოლოგიური) ქრომოსომების უბნები. გამოყოფენ შიდაქრომოსომულ (დეფიშენსი, დელეცია, დუპლიკაცია, ინვერსია) და ქრომოსომთაშორის (ტრანსლოკაცია) სტრუქტურულ დარღვევებს. შუალედური ადგილი უკავია ტრანსპოზიციას, რომელიც მიმდინარეობს, როგორც არაჰომოლოგიურ ქრომოსომთა შორის, ისე ერთი და იმავე ქრომოსომის ფარგლებში.
გენომური მუტაციები. გენომად უნდა მივიჩნიოთ ორგანიზმის მთლიანი მემკვიდრული სისტემა. იგი მოიცავს, როგორც დნმ-ის ელემენტების ნაკრებს, ისე მათ შორის კავშირის ხასიათს, რომელიც განსაზღვრავს კონკრეტულ გარემო პირობებში ონტოგენეზურ განვითარებას. გენომი შედგება ორი: ობლიგატური (ქრომოსომებში ლოკალიზებული გენები და მათი მარეგულირებელი უბნები) და ფაკულტატური (დნმ-ისა და რნმ-ის სტრუქტურები, რომელთა რაოდენობა და ტოპოგრაფია სხვადასხვა უჯრედში განსხვავებულია) ნაწილისაგან.
გენომის ობლიგატურ სისტემაში მომხდარი ცვლილებებით იცვლება ქრომოსომთა რაოდენობა. გამოყოფენ შემდეგ ფორმებს: 1. რობერტსონული ტრანსლოკაცია (ქრომოსომების ცენტრული შერწყმა ან დაშორება). 2. პოლიპლოიდია და ჰაპლოიდია (ქრომოსომთა რიცხვის ჯერადად გაზრდა ან შემცირება). 3. ანეუპლოიდია (ცალკეული ქრომოსომის რიცხობრივი ცვლილება, შემცირება ან გაზრდა). მუტაციის შედეგად წარმოქმნილი ცვლილებები ვრცლადაა განხილული გენეტიკისა და ზოგადი ბიოლოგიის სახელმძღვანელოში, ამდენად მასზედ აღარ შევჩერდებით (იხ. მითითებული ლიტერატურა).
|
|
7.3 5.3. მემკვიდრული ცვალებადობის კლასიკური და არაკანონიკური ფორმები |
▲ზევით დაბრუნება |
კლასიკური შეხედულების მიხედვით გენეტიკისა და ევოლუციურ თეორიაში არჩევენ მემკვიდრული ცვალებადობის ორ ფორმას: კომბინაციურს და მუტაციურს. მუტაციური ცვლილებები არის ელემენტარული ევოლუციური მასალა. სპონტანური მუტაციების ინდუქცია საკმაოდ დაბალი სიხშირით მიმდინარეობს. ერთსა და იმავე გენის სპონტანური მუტაციის სიხშირე მუდმივია. იგი სხვადასხვა გენისათვის განსხვავებულია და საშუალოდ ეს მაჩვენებელი 10-5-10-7 (გენზე ერთი თაობის განმავლობაში) ფარგლებში ვარირებს. სპონტანური მუტაციის ასეთი დონე ოპტიმალურია, რომელიც ევოლუციის პროცესში ჩამოყალიბდა. მუტაციის მაღალი სიხშირე გამოიწვევდა ევოლუციის პროცესის შენელებას და მას ქაოტურ ფლუქტუაციამდე დაიყვანდა.
XXს. 30-იან წლებიდან მოყოლებული მრავალი ფაქტობრივი მასალა დაგროვდა, რითაც დასტურდება, რომ მუტაციური პროცესი ექვემდებარება გენეტიკურ კონტროლს. მრავალ მცენარესა და ცხოველში გამოავლინეს გენი-მუტატორები, რომლებიც სხვა ლოკუსების მუტაბოლობას ზრდიდა. კ.მემპელმა დროზოფილას ერთ-ერთ პოპულაციაში აღმოაჩინა გენი-მუტატორი, რომელიც ჰეტეროზიგოტურ მდგომარეობაში ქრომოსომებში მუტაციას 34-ჯერ, ხოლო ჰომოზიგოტურ მდგომარეობაში ყოფნისას 70-ჯერ ზრდიდა. მ. როდსმა სიმინდში გამოავლინა გენი-მუტატორი, რომელიც სხვა ლოკუსის მუტაციურ არასტაბილობას იწვევდა.
ვინაიდან წარმოქმნილ მუტაციათა უმეტესობა საზიანო მოქმედებას ავლენს, წამოაყენეს იდეა ევოლუციის პროცესში მუტაციის სიხშირის შენელების და ყოველ სახეობაში მისი მინიმალურ დონეზე სტაბილიზაციის შესახებ (სტერტევანტი, 1937). კონკრეტული სახეობის ინდივიდების სიცოცხლის ხანგრძლიობა რაც უფრო დიდია, მით უფრო დაბალია მუტაციის ინდუქციის სიხშირე. წინააღმდეგ შემთხვევაში ჩანასახოვანი და სომატური უჯრედების ქრომოსომები გაივსებოდა საზიანო მუტაციებით.
მემკვიდრული ცვალებადობა თუ განპირობებულია გენთა კომბინაციით და რეკომბინაციით, წარმოადგენს კომბინაციურ ცვალებადობას. ამ შემთხვევაში გენები უცვლელია, იცვლება მხოლოდ მათი ურთიერთგანლაგება და გენოტიპის ერთიან სისტემაში ურთიერთდამოკიდებულების ხასიათი. იგი სამი მნიშვნელოვანი პროცესის შედეგია: 1. მეიოზის დროს ქრომოსომათა შემთხვევითი და დამოუკიდებელი განაწილების. 2. ჰომოლოგიურ ქრომოსომებს შორის მონაკვეთების რეციპროკული გაცვლის - კროსინგოვერის. ზოგჯერ იგი მიმდინარეობს ე.წ. ექტიპური ფორმით - არაჰომოლოგიურ ქრომოსომებს შორის (მაგ., სოკოებში). 3. განაყოფიერებისას გამეტების შემთხვევითი შეხვედრისა და შერწყმის. მუტაციური ზეწოლა პანმიქტური პოპულაციების ევოლუციაში მეტად უმნიშვნელოა. ახალი გენოტიპების ფორმირებაში დიდ როლს ასრულებს რეკომბინაცია და სხვა პოპულაციებიდან გენთა შემოდინება (გენთა ნაკადი). რეკომბინაციით მიიღწევა გენოტიპური ცვალებადობის მაღალი დონე (მაირი, 1968).
პოპულაციებში სპონტანური მუტაციური პროცესის შესწავლით დადგენილია ე.წ. „მოდის“ მოვლენა მუტაციაზე. ეს კანონზომიერება თავდაპირველად დროზოფილასა და ადამიანის პოპულაციებში აღწერეს, ხოლო მოგვიანებით სხვა ორგანიზმების პოპულაციებზეც დადასტურდა. გამოირკვა, რომ პოპულაციაში პერიოდულად მნიშვნელოვანი კონცენტრაციით ჩნდებიან მუტანტური ფენოტიპის ინდივიდები. ეს ფორმები მრავალი თაობის განმავლობაში შენარჩუნებულია, მოგვიანებით მათი სიხშირე თანდათანობით კლებულობს და ბოლოს ქრება. პოპულაციაში მათ ცვლის სხვა მუტაციური ფენოტიპის ინდივიდები. ეს მოვლენა პერმანენტულად მეორდება განსხვავებული მუტანტური ფენოტიპების მონაწილეობით. მუტანტური აფეთქება ზოგჯერ ვრცელ ტერიტორიას მოიცავს.
ჩრდილოეთ ნახევარსფეროში (კერძოდ ყოფილ საბჭოთა კავშირის ტერიტორიაზე, უკრაინიდან მოყოლებული შორეული აღმოსავლეთის ჩათვლით) შესწავლილ დროზოფილას პოპულაციებში 1937-1940 წწ. მაღალი სიხშირით აღინიშნებოდა სქესთან შეჭიდული y (yellow) ალელის მფლობელი ყვითელსხეულიანი ინდივიდები. 1945-1946 წწ. მათი წარმოქმნის სიხშირე დაქვეითდა და 1962-1964 წწ. ამ სახის მუტანტები აღარ შეიმჩნეოდა. ამავე ტერიტორიაზე 30 წლის განმავლობაში (1937-1967 წწ.) დროზოფილას პოპულაციებში X ქრომოსომაში ლოკალიზებული დომინანტი A ალელის (Abnormal abdomen - სეგმენტირებული მუცელი) 0.15%25-ს არ აღემატებოდა. მომდევნო 7 წელში მისმა სიხშირემ 21.7%25-ს მიაღწია. გენეტიკური ანალიზით გამოირკვა, რომ მუტაცია აბერაციის შედეგად იყო წარმოქმნილი. იგი, როგორც ჰომო - ისე ჰეტეროზიგოტების 80%25-ში ვლინდებოდა ე.ი. აღინიშნებოდა არასრული პენეტრანტულობა. ლოკუსში ინტეგრირებული მობილური ელემენტებით იგი გადატანილი აღმოჩნდა სხვადასხვა ქრომოსომებში.
1975 წ. შემდეგ A ლოკუსის მუტაციის აფეთქება თანდათანობით ჩაცხრა, სანაცვლოდ 1973 წლიდან აღინიშნებოდა სქესთან შეჭიდული sn (შერუჯული ჯაგარი) მუტაციის მკვეთრი მომატება. 1973-1977 წწ. მათი სიხშირე 0.3-0.4%25-ს შეადგენდა, ხოლო 1979-1982 წწ. 0,003%25-მდე დაქვეითდა. მომდევნო წლებში sn მუტაციის სიხშირე ნულს გაუტოლდა.
მუტანტურ გენთა სიხშირის ცვლილების ანალოგიური დინამიკა, მიოტოპიის გამომწვევი სამი განსხვავებული გენის მიმართ გამოვლენილია ადამიანის პოპულაციებში. დადგენილი ფაქტები მიუთითებენ, რომ სივრცობრივად იზოლირებული პოპულაციები შეიძლება ერთდროულად და მსგავსად შეიცვალოს. წარმოქმნილი გენეტიკური ცვლილებები დიდხანს არ რჩება პოპულაციებში, ისინი სხვა მუტაციებით იცვლებიან.
გამოთქმულია მოსაზრება, რომ ამდაგვარი ცვლილებები ვირუსული ბუნების ინფექციებითაა გამოწვეული. ვირუსის გენეტიკური მასალა ზემოქმედებს მასპინძლის გენომზე, რომელიც მის სპეციფიკურ ცვლილებას იწვევს. მოგვიანებით პოპულაციებში ჩნდებიან კონკრეტული ვირუსული ინფექციისადმი რეზისტენტული ფორმები. პოპულაციაში ვრცელდება მდგრადობის განმსაზღვრელი ალელები და ეპიზოოტია ქრება.
ბოლო პერიოდის გამოკვლევებით ახლებურად წარმოჩნდა მუტაციური პროცესის ბუნება. გამოირკვა, რომ სპონტანურად წარმოქმნილ მუტაციათა დიდი ნაწილი ინსერციული მუტაგენეზით არის ინდუცირებული. ამ გზით წარმოიქმნება, როგორც გენური, ისე ქრომოსომული მუტაციები. ინსერციული მუტაგენეზის დროს გენეტიკური მასალის მცირე მონაკვეთები ე.წ. მობილური (მიგრირებადი) გენეტიკური ელემენტები გადაადგილდებიან გენომში, ეს მექანიზმი იწვევს გენომის არასტაბილურობას.
დროზოფილას ბუნებრივ პოპულაციებში სპონტანურად შეცვლილ ლოკუსთა (folkend - ფიწლისებრი ჯაგრები, white - თეთრი თვალები, yellow - ყვითელი სხეული, cut - შეჭრილი ფრთები, bithorax - გაორმაგებული მკერდი და სხვ.) დიდი ნაწილი არასტაბილური აღმოჩნდა. ლოკუსთა არასტაბილურობას მათში ინტეგრირებული მობილური გენეტიკური ელემენტები იწვევდა. კერძოდ, არასტაბილური W ლოკუსის მუტირების სიხშირე 1თ10-3 შეადგენდა, მაშინ როდესაც ნორმალური ალელი 1თ10-6 სიხშირით მუტირებდა. მათში ინტეგრირებული აღმოჩნდა 10 ათასი ნ.წ. ზომის FB - ელემენტი. სპონტანურად ინდუცირებული W ლოკუსთა უმეტეს ნაწილს მობილური გენეტიკურ ელემენტთა განსხვავებული სისტემები აღმოაჩნდათ ინტეგრირებული. მათი მოქმედებით ლოკუსებს სხვადასხვა სახის გენეტიკური გარდაქმნები (მიკროდელეცია, დუბლიკაცია, მიკროინვერსია) აღენიშნებოდათ. X ქრომოსომაში ლოკალიზებული W ლოკუსთან შეჭიდული ტრანსპოზონი იწვევდა როგორც ლოკუსის ავტოსომურ ქრომოსომებში გადატანასა და ინტეგრაციას, ისე მაღალი სიხშირით ლეტალური მუტაციების ინდუქციას.
გამოვლენილია მობილური დისპერგირებული გენეტიკური (მდგ) ელემენტების როლი ინსერციულ მუტაგენეზში. დროზოფილას ცუტ ლოკუსში ინტეგრირებული მდგ-ელემენტი ავლენდა პლეიოტროპულ ეფექტს. იგი იწვევდა როგორც ლოკუსის არასტაბილურობას, ასევე ახალ ლოკუსთა წარმოქმნას. ტრანსპოზიცირებული ლოკუსები რამდენიმე ათეული ნუკლეოტიდური წყვილით იყვნენ ერთმანეთისგან დაცილებული. იგივე ტრანსპოზონი მკვეთრად ზრდიდა სხვა ლოკუსების მუტაბილობას. გამოვლენილია განსხვავებული კლასის მდგ-ელემენტების ერთჯერადი მასობრივი გადაადგილება (ე.წ. ტრანსპოზიციური აფეთქება). იგი იწვევდა მრავლობით მუტაგენეზს.
დადგენილია ტრანსპოზიციური მეხსიერების მოვლენა, კერძოდ მდგ-ელემენტების არაშემთხვევითი, მიმართული ჩართვა გენომის კონკრეტულ საიტებში. იგი მუტაგენეზს სპეციფიკურ ხასიათს ანიჭებდა. კერძოდ, განსაზღვრეს ხუთი განსხვავებული სისტემის მდგ-ელემენტის ლოკალიზაციის უნარი დროზოფილას განსხვავებულ ხაზებში. მათი განმეორებითი ლოკალიზაცია მიმდინარეობდა ძირითადად იმ ლოკუსებში რომელიც საწყის ხაზს აღენიშნებოდა.
არაკანონიკურ გენეტიკურ ცვალებადობას მიეკუთვნება აგრეთვე ჰიბრიდული დისგენეზი. იგი წარმოადგენს გენეტიკური ცვალებადობის კორელირებულ სინდრომს, რომელიც ნიშანდობლივია ხაზთაშორისი ჰიბრიდებისათვის, კერძოდ რეციპროკული შეჯვარებიდან ერთ-ერთისათვის. XXს. 70-იან წლებში დროზოფილაში სათანადოდ შერჩეული მშობლების შეჯვარებით გამოავლინეს ჰიბრიდული დისგენეზის მოვლენა. ამჟამად დროზოფილაში შესწავლილია ამ ფენომენის გამომწვევი რამდენიმე განსხვავებული სისტემა. ამ სისტემების ფენოტიპური ეფექტი მსგავსია, ისინი განსხვავდებიან მოლეკულურ-გენეტიკური მექანიზმებით. ჩვენ განვიხილავთ მხოლოდ ერთ-ერთ მათგანს, სადღეისოდ ყველაზე სრულყოფილად შესწავლილ P-M სისტემას (ტერმინებიდან პატერნალ და მატერნალ - შესაბამისად მამისეული და დედისეული). ამ სისტემების QMXWP შეჯვარების შემთხვევაში F1-ის ჰიბრიდებს აღენიშნებათ შემდეგი გენეტიკური ანომალიები: სრული ან ნაწილობრივი სტერილობა; რეკომბინაციის პროცესი მამრებში; მომატებული მუტაბილობა, რომელიც ხშირად ალელთა არასტაბილობას იწვევს; ქრომოსომული აბერაციებისა და ქრომოსომთა განურიდებლობის მაღალი სიხშირით ინდუქცია.
ჰიბრიდულ დისგენეზს P-ელემენტების მფლობელ მამისეული ქრომოსომებისა და M-ტიპის დედისეულ ციტოპლაზმას შორის შეუთავსებლობა იწვევს. იმ შემთხვევაში, თუ დედისეული ქრომოსომები ფლობენ P-ელემენტებს, მათში მცირე რაოდენობითაა და ანომალურია. P-ციტოტიპის ინდივიდს ელემენტის 30-60 ასლი გააჩნია, რომელიც გაფანტულია (დისპერგირებულია) გენომში. ასლების ნაწილი სრულია (ნორმალური), ნაწილი კი დეფექტური (გააჩნიათ სხვადასხვა სახის მიკროდელეციები). გენომში P-ელემენტების ტრანსპოზიციას ახორციელებს ფერმენტი - ტრანსპოზაზა. მეორე კოდირებული ფერმენტია - რეპრესორი (რეგულატორი): რეპრესორის დაგროვების შემთხვევაში, იგი ახდენს ტრანსპოზაზას ინჰიბირებას, რის გამოც P-ციტოტიპის უჯრედებში იბოჭება P-ელემენტების ტრანსპოზიცია. ჰიბრიდული დისგენეზის დროს P-ელემენტების ინტეგრაცია მაღალი სიხშირით მიმდინარეობს. P-ელემენტების ტრანსპოზიცია ნიშანდობლივია მხოლოდ ჩანასახის უჯრედებისათვის პრემეიოზურ სტადიაზე. მხოლოდ ამ უჯრედებში მიმდინარეობს P-ელემენტების ტრანსკრიპტის სპლაისინგი, ტრანსლაცია და ტრანსპოზაზას სინთეზი.
|
|
8 §6. ევოლუციის ელემენტარული მოვლენა - პოპულაციის გენეტიკური შედგენილობის ცვლილება |
▲ზევით დაბრუნება |
როგორც უკვე აღვნიშნეთ, ევოლუციის ელემენტარული სტრუქტურა (ერთეული) არის პოპულაცია, ხოლო ევოლუციის ელემენტარულ მოვლენად ამ სტრუქტურის მემკვიდრული ცვლილება არის მიჩნეული. ყველა ცოცხალ ორგანიზმში მიმდინარე სპონტანური მუტაციური პროცესის გამო თაობათა მანძილზე (დროის ამა თუ იმ მონაკვეთში) არსებული ნებისმიერი ერთობლიობა ჰეტეროგენული ხდება. ჰომოგენური მხოლოდ ახლად წარმოქმნილი წმინდა ხაზები და კლონებია. ფენოტიპურად მეტ-ნაკლებად ერთგვაროვანწევრებიანი პოპულაციაც კი განსხვავებული გენოტიპების მქონე ერთობლიობაა (ჰეტეროგენულია).
ჰეტეროგენულობას იწვევს როგორც მუტაციური პროცესის მუდმივი ზეწოლა, ისე გენეტიკური მასალის რეკომბინაცია (იხ. §5). თუ ფაქტორები პოპულაციის გენეტიკური სტრუქტურის (გენოფონდის) დინამიკაზე უმნიშვნელოდ მოქმედებს, მაშინ მისი გენეტიკური შედგენილობის უმნიშვნელო ფლუქტუაცია გვექნება. იგი ძირითადად ხანგრძლივი დროის განმავლობაში უცვლელი დარჩება. როდესაც ფაქტორები პოპულაციის გენეტიკურ შედგენილობაზე (გენოფონდზე) ხანგრძლივად და ინტენსიურად მოქმედებს, მაშინ იგი უსათუოდ შეიცვლება. თუ გენოფონდის ხანგრძლივი და მიმართული (ვექტორიზებული) ცვლილება მიმდინარეობს, მაშინ თავს იჩენს ევოლუციურად მნიშვნელოვანი ხდომილება ელემენტარული ევოლუციური მოვლენა. ელემენტარული ევოლუციური მოვლენა არის პოპულაციის გენოფონდის ხანგრძლივი, შეუქცევადი და მიმართული (ვექტორიზებული) ცვლილება.
ევოლუციის ელემენტარული მოვლენა ჯერ კიდევ არ წარმოადგენს ევოლუციურ პროცესს. ამასთანავე, ნებისმიერი ევოლუციური პროცესის მიმდინარეობას (უმცირესით დაწყებული, უდიდესი მასშტაბით დამთავრებული) გენოფონდის ცვლილება უდევს საფუძვლად. ამრიგად, ევოლუციის ელემენტარული სტრუქტურა არის პოპულაცია, ხოლო ელემენტარული მოვლენა კი მისი გენეტიკური სტრუქტურის ცვლილება. მოწოდებული თეორიული ფორმულირება მკაფიოდ და ცალსახადაა ჩამოყალიბებული. მიკროევოლუციის მოძღვრების განვითარების თანამედროვე დონეზე ამდაგვარი ფორმულირება სავსებით დამაკმაყოფილებლად შეიძლება მივიჩნიოთ.
მაგრამ ბუნებაში ხშირად ვხვდებით ისეთ კომპლექსურ მოვლენებს, რომლებიც მკაფიოდ არ არის გამოვლენილი, ან მისი საზღვრები წაშლილია. განსაკუთრებით ეს ეხება ყველა იმ დინამიურსა და ფლუქტუირებად სტრუქტურასა და მოვლენას, რომელიც დროსა და სივრცეში იცვლება. ამ სახის სტრუქტურათა რიცხვს მიეკუთვნება პოპულაცია და მისი გენეტიკური სტრუქტურა. ზუსტი და მკაფიო თეორიული ფორმულირება აუცილებელი, მაგრამ არასაკმარისი პირობაა. მაშასადამე აუცილებელია, მრავალ კონკრეტულ პოპულაციაში გენოფონდის ფლუქტუაციისა და ვექტორიზებული ცვლილებების შესწავლა. შესაძლებელია სახეობის მსგავსად პოპულაციაც დროის გარკვეულ მონაკვეთში დივერგირების (განტოტვის) პროცესში იმყოფებოდეს, რომლის შედეგადაც საწყისი პოპულაცია ორ ან ორზე მეტ პოპულაციად ჩამოყალიბდება.
პოპულაციის გენეტიკური სტრუქტურის დინამიკაზე მოქმედი ფაქტორი შესაძლოა მრავალი აღმოჩნდეს, ვინაიდან არაერთი მოვლენა სხვადასხვა გზით და მექანიზმებით გავლენას ახდენს გენოფონდზე. მოქმედების ხასიათისა და შედეგის მიხედვით შეიძლება გამოიყოს ძირითადი ელემენტარული ფაქტორები, რომლებიც აუცილებელი და საკმარისია ელემენტარული ევოლუციური პროცესის წარმართვისათვის. ევოლუციის ძირითად ელემენტარულ ფაქტორებს წარმოადგენს: ბუნებრივი გადარჩევა, მუტაციური პროცესი, გენთა დრეიფი, გენთა ნაკადი, იზოლაცია. მათგან გენოფონდის მდგრად, შეუქცევად მიმართულ ცვლილებას მხოლოდ ბუნებრივი გადარჩევა იწვევს.
|
|
9 §7. ევოლუციის ელემენატარული ფაქტორები |
▲ზევით დაბრუნება |
|
|
9.1 7.1. მუტაციური პროცესი - ევოლუციის ელემენტარული ფაქტორი |
▲ზევით დაბრუნება |
|
|
9.1.1 მუტაციის წარმოქმნის სიხშირე და მიზეზები |
▲ზევით დაბრუნება |
მუტაციური პროცესი პოპულაციის დინამიკაზე მოქმედი ფაქტორია. ყველა ცოცხალ ორგანიზმში მუტაციური პროცესი სპონტანურად და მუდმივად მიმდინარეობს. მუტაციური ცვალებადობის დროს ადგილი აქვს გენეტიკური მასალის თვისებრივ ან რაოდენობრივ ცვლილებას. მუტაციები ესაა ელემენტარული ევოლუციური მასალა. ადაპტური მნიშვნელობის მიხედვით მუტაციები შეიძლება იყოს სასარგებლო, ნეიტრალური ან საზიანო. ეს დაყოფა პირობითია, რადგანაც გარემო პირობების შეცვლისას სასარგებლო მუტაციები შეიძლება გახდეს საზიანო ან - პირიქით. გარემო პირობების შესაბამისად ერთი და იგივე მუტაცია შეიძლება პირობით საზიანო ან პირობით სასარგებლო აღმოჩნდეს. მუტაციების სადღეისოდ არსებული ყველა კლასიფიკაცია არასრულყოფილია (იხ. §5.2).
რ. ფონ ბერსტოლის კონცეფციის თანახმად, გენური მუტაციის სპონტანური ინდუქცია რეპლიკაციის, რეკომბინაციის ან რეპარაციის პროცესში მომხდარი დარღვევის შედეგია. მიკროორგანიზმების, მცენარეებისა და ცხოველების მრავალ სახეობაში, აგრეთვე ადამიანში შესწავლილია ზოგიერთი გენის სპონტანური მუტაციის სიხშირე. ეს მაჩვენებელი სხვადასხვა გენისათვის განსხვავებულია და საშუალოდ 10-5-10-7-მდე ფარგლებში ვარირებს (იხ.: ცხრილი 7.1), თუმცა ზოგიერთი გენი უფრო დაბალი ან მაღალი სიხშირით მუტირებს.
ცხრილი 7.1
სპონტანური მუტაციის წარმოქმნის სიხშირე
(ს. ინგე-ვეჩტომოვი, 1989)
სახეობა და ნიშანი |
მუტაციის სიხშირე გენომზე |
scherichia coli |
4•10-10
1•10-9
2•10-8
|
Clamidomonas reinhardti |
1•10-6 |
eurospora crassa |
4•10-8
|
Zea mays |
1•10-6 1•10-5 |
rosophila melanogaster |
4•10-5 1•10-4 3•10-5 |
us musculus |
8•10-6 |
გენის სპონტანური მუტაციის სიხშირეს გამოსახავენ ერთი თაობის გამეტების (ან გენომის) საერთო რაოდენობის თანაფარდობით მუტაციის შემცველ გამეტებთან. ერთი და იმავე გენის სპონტანური მუტაციის სიხშირე განსაზღვრულ ფარგლებში ვარირებს, რომელიც საკმაოდ დაბალია. იმის გამო, რომ ორგანიზმებს გენთა საკმაოდ დიდი რაოდენობა მოეპოვებათ, მუტაციის მქონე გამეტების საერთო რაოდენობა მნიშვნელოვან სიდიდეს აღწევს. კერძოდ, მისი საერთო რაოდენობა რამდენიმე პროცენტიდან (ბაქტერიები, ერთუჯრედიანი წყალმცენარეები და სოკოები) 25%25-მდე (დროზოფილა) ვარირებს. წარმოიქმნება საკმაოდ დიდი მასალა ბუნებრივი გადარჩევისათვის.
გამოირკვა, რომ მუტაციური პროცესი გენეტიკური მექანიზმებით კონტროლდება. გამოვლენილია გენები, რომლებიც ცვლიან (ზრდიან ან ამცირებენ) მუტაციის სიხშირეს. სპონტანურ მუტაციურ პროცესს განიხილავენ, როგორც ადაპტურ ნიშანს. იგი ბუნებრივი გადარჩევის მოქმედების გზით ყალიბდება. მუტირების რეგულირება ევოლუციის დროს სხვადასხვა გზით მიიღწევა: რეპარაციული სისტემის ეფექტურობით და უჯრედში ავტომუტაგენური ან ანტიმუტაგენური ნივთიერებების კონცენტრაციის ცვლილებით.
|
|
9.1.2 მუტაციის გამოვლენა და სპექტრი |
▲ზევით დაბრუნება |
მუტაციის გამოვლენა ერთი მხრივ შეცვლილი ალელის დომინანტურ-რეცესიულ მდგომარეობაზეა დამოკიდებული. დომინანტური მუტაცია პირველსავე თაობაში ვლინდება და ბუნებრივი გადარჩევის მოქმედებას ექვემდებარება. თუ მუტაცია უპირატესობას ანიჭებს ორგანიზმს, იგი შეინახება, საზიანო კი გამოირიცხება, რადგან მისი მფლობელი ინდივიდი შესაბამისად ტოვებს შთამომავლობას ან იღუპება. თუ ახლად წარმოქმნილი მუტაცია რეცესიულია, მაშინ იგი ჰეტეროზიგოტულ მდგომარეობაში მრავალი თაობის განმავლობაში შეიძლება ხანგრძლივად დარჩეს პოპულაციაში, რადგან იგი ორგანიზმში ჰეტეროზიგოტულ მდგომარეობაში მოქმედებას ვერ ავლენს. ბუნებრივი გადარჩევა მასზე გავლენას ახდენს მხოლოდ მაშინ, როდესაც მუტანტური ალელი ჰომოზიგოტურ მდგომარეობაში გადადის და ფენოტიპურად ვლინდება.
მეორე მხრივ, მუტანტური ალელის გამოვლენა დამოკიდებულია აგრეთვე იმ გარემოზე, რომელშიც მუტანტური ალელი ხვდება. ხშირად ერთი და იგივე მუტანტური ალელი სხვადასხვა ორგანიზმში განსხვავებულ ფენოტიპურ ეფექტს ახდენს, რაც ექსპრესიულობითა (გენის ფენოტიპურად გამოვლენის ხარისხი) და პენეტრანტულობით (პოპულაციაში გენის გამოვლენის სიხშირე) არის გამოხატული. გენის გამოვლენის სიხშირეს პენეტრანტულობა ეწოდება. ზოგიერთი გენის ერთი და იგივე ალელი პოპულაციის მხოლოდ ნაწილში ვლინდება, დანარჩენში კი - არა. პენეტრანტულობის ხარისხს განსაზღვრავენ ალელის მფლობელ ინდივიდთა საერთო რაოდენობის ფენოტიპურად გამოვლენილ ინდივიდებთან პროცენტული თანაფარდობით. სრული პენეტრანტულობისას მუტირებული გენის დომინანტური ან რეცესიული ალელი უკლებლივ ყველა ინდივიდში (100%25) ვლინდება. არასრული პენეტრანტულობის დროს გენის დომინანტური ან რეცესიული ალელის ეფექტი პოპულაციის წევრთა მხოლოდ ერთ ნაწილში ვლინდება.
ზოგჯერ ერთსა და იმავე ალელის გამოვლენის ხარისხი პოპულაციის წევრებში განსხვავებულია. ერთნაირი გენოტიპის ინდივიდებში გენის გამოვლენის ნაირგვარ დონეს ექსპრესულობა ეწოდება. პენეტრანტულობა გვიჩვენებს პოპულაციის წევრთა რა ნაწილში ვლინდება ნიშანი, ექსპრესულობა მიუთითებს, თუ რა ოდენობით ვარირებს გენის გამოვლენის დონე. ექსპრესულობა შინაგან და გარეგან გარემოზე ერთნაირი გენოტიპის მქონე ინდივიდების რეაქციას გამოხატავს. ამ რეაქციას ორგანიზმებისა და მთლიანად პოპულაციისათვის დიდი ადაპტური მნიშვნელობა აქვს. ამ უნარს ორგანიზმი ბუნებრივი გადარჩევით ინარჩუნებს.
ნებისმიერი ნიშან-თვისების (ფენის) ჩამოყალიბება გარემო პირობებისა და გენოტიპის ურთიერთქმედების შედეგად ხორციელდება. მაგალითად, დროზოფილაში მუტაცია ვესტიგიალ-ის („ჩანასახოვანი ფრთები“) გამოვლენა გარემოს ტემპერატურაზეა დამოკიდებული. 14Co ტემპერატურაზე განვითარებულ ინდივიდს ფრთები საერთოდ არ გააჩნია, ხოლო 31Co ტემპერატურაზე კი თითქმის ნორმალური ფრთები აქვს (სურ.7.1.).

სურ. 7.1. სხვადასხვა ტემპერატურის გავლენა დროზოფილას ფრთის განვითარებაზე. 1 - ფრთის ზომის ცვლილება სხვადასხვა ტემპერატურაზე განვითარებულ მუტანტ ბუზებში. 2 - მუტაცია „ვესტიგიალ“ - (ჩანასახოვანი ფრთები) განვითარებული 29C0 ტემპერატურაზე (გ. გულიაევი, 1971).
მუტაციის სპონტანური ინდუქცია ორგანიზმის ონტოგენეზის ნებისმიერ პერიოდში ხდება, გამეტებიდან და ზიგოტებიდან მოყოლებული, ვიდრე ორგანიზმის სიბერით დამთავრებული. იგი ორგანიზმის ნებისმიერი ნიშან-თვისების განმსაზღვრელ გენში მიმდინარეობს. სპონტანურ მუტაციურ პროცესს ფართო სპექტრი გააჩნია. განსხვავებული დონით ორგანიზმის ყველა ნიშნისა და თვისების (მორფოლოგიური, ფიზიოლოგიური, ბიოქიმიური, ეთოლოგიური და ა.შ.) განმსაზღვრელი გენების მუტაცია ხდება. მუტაციამ შეიძლება შეასუსტოს ან გააძლიეროს ნიშანი. მუტაციის შედეგად ნიშანი შეიძლება მკვეთრად იყოს გამოხატული (ლეტალური მუტაციის ჩათვლით) ან თითქმის უმნიშვნელოდ (ე.წ. „მცირე“ მუტაციები). წარმოიქმნება საკმაოდ მრავალფეროვანი მასალა ევოლუციის პროცესისათვის.
|
|
9.1.3 მუტაციების გავრცელება და მუტაციური პროცესის გავლენა პოპულაციის სტრუქტურაზე |
▲ზევით დაბრუნება |
მიკროორგანიზმთა, მცენარეთა და ცხოველთა პოპულაციების შესწავლისას გამოირკვა, რომ ისინი განსხვავდებიან მუტაციის შეხვედრის სიხშირით. ბუნებაში არ გვხვდება ერთსა და იმავე სახეობის ორი ერთნაირი გენეტიკური სტრუქტურის (გენოფონდის) მქონე პოპულაცია. ერთიმეორისაგან განსხვავდებიან როგორც მეზობელი, ისე საკმაოდ დაცილებული პოპულაციები. მცენარეების (დევისპირა, ია, თავცეცხლა და სხვ.), უხერხემლო ცხოველების (დროზოფილა, ჭიამაია), ხერხემლიანების (სახლის თაგვი, ვირთაგვა, მემინდვრია და სხვ.) პოპულაციების შესწავლისას გამოირკვა, რომ ისინი განსხვავდებიან ერთიმეორისაგან გენური, ქრომოსომული ან გენომური მუტაციების შეხვედრის სიხშირით.
ახლად წარმოქმნილი მუტაციების დიდი ნაწილი ორგანიზმისათვის საზიანოა, რადგან ისინი ცვლიან ევოლუციის პროცესში ჩამოყალიბებულ ერთიან გენეტიკურ სტრუქტურას. ინდივიდის გენეტიკური სტრუქტურის (გენოტიპის) ჩამოყალიბება ევოლუციის პროცესში უკეთესი კომბინაციების გადარჩევის გზით მიმდინარეობს. ნებისმიერი გადახრა ამ სისტემაში იწვევს დისბალანსს. ამიტომაც გადახრები მეტწილად საზიანოა. ბუნებრივ პოპულაციებში ერთი მხრივ, რეცესიული მუტაციების ფართო გავრცელება წარმოადგენს გენეტიკურ ტვირთს პოპულაციისათვის, მაგრამ მეორე მხრივ, ის, როგორც სამობილიზაციო რეზერვი, ემსახურება პროგრესულ ევოლუციას. გენთა ნაირგვარი მუტაციები, რომლებიც განსხვავებული სიხშირით წარმოიქმნება, არღვევს პოპულაციაში არსებულ წონასწორობას, ხოლო შემდგომ კვლავ მყარდება გენოტიპთა ახალი წონასწორობა. ამრიგად, პოპულაციის გენეტიკური სტრუქტურა იცვლება ერთი წონასწორობიდან მეორე წონასწორობამდე, რაც მისი დინამიკის ხასიათს განსაზღვრავს.
პოპულაციის გენოფონდი ყოველ თაობაში ახალი მუტაციებით ივსება. ამ პროცესს მუტაციური ზეწოლა ეწოდება. მუტაციური ზეწოლის დონე განსაზღვრავს პოპულაციაში სხვადასხვა გენის ალელების სიხშირეს. მუტაციის გავრცელებას პოპულაციაში განაპირობებს, უწინარეს ყოვლისა, გენის მუტაბილობა (ამა თუ იმ ლოკუსის მუტაციის უნარი), მაგრამ მისი დონე დამოკიდებულია იმაზე, თუ რა გავლენას ახდენს მუტაცია ინდივიდის ცხოველმყოფელობასა და ნაყოფიერებაზე.
სქესობრივი პროცესის მეშვეობით ხდება საზიანო მუტაციების „უვნებელყოფა“. კერძოდ, შეინიშნება ამ ტიპის მუტაციების ჰეტეროზიგოტულ მდგომარეობაში გადასვლა. მრავალი მუტაცია ჰეტეროზიგოტულ მდგომარეობაში მეტწილად აძლიერებს სიცოცხლის უნარიანობას (ხდება ე.წ. ჰეტეროზისი). სქესობრივი პროცესით ერთი მხრივ ხორციელდება მუტაციის გზით ახლად წარმოქმნილი გენეტიკური მასალის შენახვა, მეორე მხრივ, საზიანო მუტაციების უვნებელყოფა, გადაყავს რა იგი „ფარულ“ (ჰეტეროზიგოტურ) მდგომარეობაში.
მუტაციათა მცირე ნაწილი სასარგებლო ან ნეიტრალური აღმოჩნდება. ბუნებრივი გადარჩევის მოქმედების შედეგად ხდება, თაობათა მანძილზე მათი შენარჩუნება და შეიძლება მნიშვნელოვანი როლიც შეასრულოს ევოლუციის პროცესში. ამჟამად მიაჩნიათ, რომ ევოლუციაში არსებით როლს მცირე მუტაციები ასრულებენ, რადგან ისინი ნიშნის უმნიშვნელო ცვლილებას იწვევენ* სასარგებლო მცირე მუტაციების გენოტიპში დაგროვებისას ნიშნის არსებითი ცვლილება ხდება.
მრავალი მუტაცია პლეიოტროპული მოქმედებისაა: ზოგჯერ ერთ ნიშანზე დადებითად მოქმედებს, მეორეზე კი უარყოფითად. თუ დადებითი ეფექტი ჭარბობს უარყოფითს, მაშინ ასეთი მუტაცია ჰეტეროზიგოტულ მდგომარეობაში ინახება. მიმდინარეობს გენი-მოდიფიკატორებისა და მცირე მუტაციების მეშვეობით პლეიოტროპულ მუტაციაზე ზემოქმედება და უარყოფითი ეფექტის შესუსტება. როდესაც მუტანტურ ინდივიდებს შესამჩნევი უპირატესობა აქვთ საწყის ფორმებთან შედარებით: სწრაფად ვრცელდებიან ბუნებაში და სახლდებიან სახეობისათვის ადრე არსებობისათვის ნაკლებ ხელსაყრელ პირობებში.
ოკეანურ კუნძულებზე, სადაც განუწყვეტლივ ქრის ქარი, გავრცელებულია მწერები, რომლებსაც ფრენის უნარი არა აქვთ.უნდა აღინიშნოს, რომ ეს მოვლენა ჰიპოთეზის სახით ჯერ კიდევ ჩ. დარვინმა განიხილა. იშვიათი მუტაციებიც კი, რომლებიც ფრთების რედუქციას იწვევენ, სასარგებლო აღმოჩნდებოდა. ფრთიანი მწერები ქარს მიჰქონდა ზღვაში და მასიურად იღუპებოდნენ. დროზოფილაში ინდუცირებულია მრავალი სახის მუტაცია, რომლებიც ფრთის ცვლილებას იწვევს. მაგ. ვესტიგიალ, აპტეროუს („უფრთო“) და სხვა. mutacia vestigial-ი პლეიოტროპული მოქმედებისაა, იგი ნაყოფიერებას აქვეითებს.
ხელოვნური გაწონასწორებული პოპულაცია, რომელიც ჩანასახოვანფრთიანი და ნორმალურფრთიანი (ველური ტიპი) ინდივიდებისაგან შედგებოდა, ყუთებში მოათავსეს და ზღვის სანაპიროზე განალაგეს. ორი თვის განმავლობაში ქარიან ამინდში, პოპულაციის შემადგენლობა შეიცვალა (2,5%25-დან ჩანასახოვანფრთიანი ბუზების რაოდენობა 67%25-მდე გაიზარდა). უქარო ამინდის დროს კი პოპულაციის შემადგენლობა არ იცვლებოდა. როდესაც ყუთები დახურულ შენობაში მოათავსეს, ფრთიანმა ფორმებმა, მაღალი ნაყოფიერების გამო, ჩანასახოვანი ფორმები ძლიერ შეავიწროვეს. ოკეანის კუნძულებზე უფრთო ფორმების გაჩენა ბუნებრივი გადარჩევის შედეგად ექსპერიმენტულად იქნა დამტკიცებული.
განუწყვეტელი ცვალებადობა ორგანიზმისათვის საზიანო იქნებოდა, რადგანაც გენების სასარგებლო კომბინაციების დამყარების შესაძლებლობა მოისპობოდა. ამიტომ ევოლუციის პროცესში გამომუშავდა მექანიზმები, რომლებიც ზღუდავენ როგორც მუტაციის სიხშირეს, ისე სპექტრს. მოლეკულურ დონეზე ასეთი მექანიზმია კოდში სინონიმური ტრიპლეტების არსებობა, რეპარაციის სისტემები, გენეტიკური პროცესების გენეტიკური კონტროლი* უჯრედის დონეზე ქრომოსომების ინდივიდუალურობა და შეჭიდულ გენთა არსებობა* ორგანიზმის დონეზე მიტოზური გაყოფა და ქრომოსომთა დიპლოიდური კომპლექტის არსებობა* პოპულაციის დონეზე კი პანმიქსიის დარღვევა და არსებობისათვის ბრძოლაში ინდივიდთა გარკვეული ნაწილის დაღუპვა (ელიმინაცია).
|
|
9.1.4 მუტაციური პროცესის არამიმართული ხასიათი |
▲ზევით დაბრუნება |
გენეტიკასა და ევოლუციურ თეორიაში მნიშვნელოვანი კონცეფციაა ის, რომ მუტაციურ პროცესს არამიმართული და შემთხვევითი ხასიათი აქვს. ეს კი იმას ნიშნავს, რომ ახლად წარმოქმნილი მუტაცია არაადეკვატურია მასზე მომქმედი ფაქტორის მიმართ. მაგ., კოლხიცინით გამოწვეული პოლიპლოიდია, ორგანიზმს ამ ალკალოიდის მოქმედებისაგან არ იცავს, ხოლო რადიაციით ინდუცირებული მუტაციები არ იწვევენ მისგან დამცველი ნივთიერებების - რადიოპროტექტორების გამომუშავებას. ყოველი მუტაგენი მუტაციების მთელ სპექტრს იწვევს, ისინი სხვადასხვა სახის მუტაციის ინდუქციას იწვევენ. სხვადასხვა მუტაგენს კი აქვს მსგავსი მუტაციის ინდუქციის უნარი. დნმ-ის მოლეკულაში ერთი წყვილი ნუკლეოტიდის ცვლილება მუტაციის ინდუქციას იწვევს. რადიუსი, სადაც მუტაციური ცვლილება ხდება, საკმაოდ მცირეა და 10-7სმ-ის ტოლია. მუტაციური პროცესი შემთხვევითი პროცესია. შეუძლებელია წინასწარ განისაზღვროს, როდის და სად მოხდება ესა თუ ის მუტაცია. მაგრამ შეიძლება ვივარაუდოთ მოცემულ ლოკუსში რა სიხშირით მოხდება მუტაციის ინდუქცია.
|
|
9.1.5 მუტაციური პროცესის მნიშვნელობა ევოლუციაში |
▲ზევით დაბრუნება |
სხვადასხვა ტიპის მუტაციების წარმოშობის სიხშირით პოპულაციების განსხვავება, მუტაციების ორგანიზმის მნიშვნელოვან ნიშნებზე მოქმედება, ბუნებრივი პოპულაციების ჰეტეროგენულობა და მათი გაჯერება მუტაციებით საშუალებას გვაძლევს დავასკვნათ, რომ მუტაციები ევოლუციის ელემენტარული მასალაა.
მუტაციური პროცესის ევოლუციური როლი, აგრეთვე, იმაში მდგომარეობს, რომ იგი განაპირობებს პოპულაციაში მაღალი დონით ჰეტეროგენულობის შენარჩუნებას. ჰეტეროგენულობას კი ეფუძნება ევოლუციის სხვა ელემენტარული ფაქტორების, მათ შორის ბუნებრივი გადარჩევის, მოქმედება. ამრიგად, მუტაციური პროცესი არის ელემენტარული ევოლუციური მასალის მიმწოდებელი ფაქტორი.
|
|
9.2 7.2. გენთა დრეიფი როგორც ევოლუციის ელემენტარული ფაქტორი |
▲ზევით დაბრუნება |
პოპულაციაში ალელთა სიხშირეზე გავლენას ახდენს გენთა დრეიფი. ამერიკელმა მეცნიერმა ს.რაიტმა და რუსმა გენეტიკოსებმა ნ.დუბინინმა და დ. რომაშოვმა ერთმანეთისაგან დამოუკიდებლად და ერთდროულად დაადგინეს (1931) ნებისმიერ განსაზღვრულ პოპულაციაში თაობათა მანძილზე ალელთა მოცემული კონცენტრაციის განუსაზღვრელად შენარჩუნების შეუძლებლობა. პოპულაციაში ალელთა კონცენტრაციის დარღვევა შეიძლება მოხდეს მუტაციური პროცესისა და ბუნებრივი გადარჩევის გარეშეც. თაობათა მანძილზე (ბიოლოგიურ დროში) შემთხვევითი პროცესებით გამოწვეულ ალელთა სიხშირის ცვლილებას პოპულაციაში გენთა დრეიფი (გენეტიკურ-ავტომატური პროცესი) ეწოდება. გენთა დრეიფი განსაკუთრებით შესამჩნევია მცირერიცხოვან პოპულაციებში.
ალელთა წონასწორობის ჰარდი-ვაინბერგის კანონი 30-იან წლებამდე უნივერსალურად მიაჩნდათ. იგი არ ირღვევა, როდესაც პოპულაცია უსასრულოდ მრავალრიცხოვანია და მიმდინარეობს პანმიქსია (იხ.§4.). მაგრამ სინამდვილეში ბუნებრივ პოპულაციებში ეს პირობა არასდროს არ რეალიზდება. საარსებო პირობების ცვლილების შედეგად ხდება პოპულაციის რიცხოვნობის მკვეთრი ზრდა ან შემცირება, ე.წ. სიცოცხლის (პოპულაციური) ტალღები (ჩეტვერიკოვი, 1926). ნებისმიერი პოპულაცია გადის მაქსიმალური და მინიმალური რიცხოვნობის პერიოდებს, რომელსაც შეიძლება ჰქონდეს ციკლური ან აციკლური ხასიათი. ამ სახის ფლუქტუაციის მიზეზი საკმაოდ მრავალგვარია. იგი შეიძლება გამოიწვიოს როგორც აბიოტურმა, ისე ბიოტურმა ეკოლოგიურმა ფაქტორებმა. ორგანულ ბუნებაში ნაირგვარ პოპულაციურ ტალღებს ვხვდებით. განვიხილოთ ძირითადი მათგანი:
1. ხანმოკლედ მცხოვრები ორგანიზმების რიცხოვნობის ციკლური მერყეობა. იგი ახასიათებს ერთწლიან მცენარეებს, მრავალ მწერს, სოკოსა და მიკროორგანიზმს. რიცხოვნობის ამდაგვარი სეზონური მერყეობა მკაფიოდაა გამოხატული მიკროორგანიზმებში. მაგალითად, გრიპისა და სხვა მსგავსი დაავადებების საგაზაფხულო და საშემოდგომო ტალღები, რაც ვირუსების მასობრივი გამრავლებითაა გამოწვეული.
2. რიცხოვნობის აციკლური მერყეობა, გამოწვეული სხვადასხვა ფაქტორის შემთხვევითი თანხვედრით. იგი უმთავრესად გამოწვეულია კვებით ჯაჭვში კონკრეტული პოპულაციისათვის ხელსაყრელი პირობების შექმნით. მაგ. მტაცებლების რიცხვის შემცირება მსხვერპლთათვის, მტაცებლებისათვის საკვები რესურსების გაზრდა და სხვა.
3. ახალ რაიონებში სახეობათა რიცხოვნობის აფეთქებები, როდესაც არა ჰყავთ ბუნებრივი მტრები. საილუსტრაციოდ შეიძლება დავასახელოთ XIX-XXსს. ავსტრალიაში ბოცვერის მასობრივი გავრცელება* ჩრდილოეთ ამერიკაში - სახლის ბეღურისა* ევრაზიაში - კანადური ელოდეას, ამერიკული წაულას, ონდანტრასი და კოლორადოს ხოჭოსი, საქართველოში - დიდი ლაფანჭამიასი და სხვა (იხ.: სურ. 7.2). XVI-XVIIსს. ზღვაოსნობის განვითარების შედეგად ვირთაგვები (ღატტუს ნორვეგიცუს) განსახლდნენ მთელ მსოფლიოში. ადამიანის სამოსახლოსთან ნაგავსაყრელებმა ხელი შეუწყო ოთახის ბუზის (usca domestica) რიცხოვნობის მკვეთრ ზრდას.
4. ბუნებრივ „კატასტროფებთან“ დაკავშირებული რიცხოვნობის მკვეთრი მერყეობა. იგი გამოწვეულია ეკოცენოზების ან მთელი ლანდშაფტების დაშლით, გვალვით, ხანძრით და ა.შ. სწრაფი გადაადგილების უნარის მქონე ფორმები (მაგ., მსხვილი ძუძუმწოვრები, ფრინველები და სხვა) და ნიადაგის ღრმა ფენებში ბინადარი ინდივიდები, შედარებით ნაკლებად ნადგურდებიან, ხოლო ხმელეთის ზედაპირზე ბინადარი უძრავი ან ნაკლებმოძრავი (მცენარეები, მოლუსკები, მყვარები, რეპტილიები და სხვ.) ფორმები დიდი რაოდენობით იღუპებიან.
სურ. 7.2. ონდატრას და კოლორადოს ხოჭოს მიერ ტერიტორიის სწრაფი ექსპანსია. ა. ონდატრას (Ondatra zibetia) გავრცელება ცენტრალურ ევროპაში 1909-1927 წლებში. ბ. კოლორადოს ხოჭოს (Leptinotersa decimlineata) სწრაფი განსახლება 1930-1935 წლებში, ევროპაში (ნ. ტიმოფეევ-რესოვსკი და სხვ., 1977).
პოპულაციის გენოფონდის ცვლილება ხდება როგორც მაქსიმალური, ისე მინიმალური რიცხოვნობისას. რიცხოვნობის გაზრდისას ადრე კუნძულებრივად განლაგებული (განცალკევებული) პოპულაციების გენოფონდი ერთიანდება. იმის გამო, რომ თითოეული პოპულაცია თავისი გენეტიკური სტრუქტურით - გენოფონდით უნიკალურია, გაერთიანების გამო მათ საწყისისაგან განსხვავებული გენოფონდი უყალიბდებათ. რიცხოვნობის ზრდა პოპულაციათა შორის მიგრაციების ინტენსიფიკაციას იწვევს, რაც ხელს უწყობს გენოფონდში ალელთა კონცენტრაციის ცვლილებას. რიცხოვნობის ზრდას, ჩვეულებრივ, თან ახლავს შიგასახეობრივი არსებობისათვის ბრძოლის გამძაფრება და არეალის გაფართოება. როდესაც პოპულაციაში რიცხოვნობის მატება პიკს აღწევს, ინდივიდთა ზოგიერთი ჯგუფი არეალის საზღვრებს გარეთ მიგრირებს. ისინი მათთვის უჩვეულო საარსებო გარემოში აღმოჩნდებიან. განსხვავებული საარსებო პირობების გამო მეტწილად ასეთი მცირერიცხოვანი ჯგუფები იღუპებიან. იშვიათად, სასარგებლო გენეტიკური სტრუქტურის გამო ზოგიერთი პოპულაცია გადარჩება. ისინი იზოლირებული აღმოჩნდებიან პოპულაციის ძირითადი მასისაგან. ბუნებრივია, მათი გენოფონდი განსხვავდება ძირითადისაგან და მასზე ღარიბია.
დიდი რიცხოვნობის მრავალ ბუნებრივ პოპულაციაში გენთა დრეიფი უფრო ნაკლებ ეფექტს იწვევს, ვიდრე ბუნებრივი გადარჩევა და გენთა ნაკადი. როდესაც მცირერიცხოვან პოპულაციაში გადარჩევისა და გენთა ნაკადის ეფექტი მცირეა, მაშინ გენთა დრეიფით განპირობებული ხანგრძლივი გენეტიკური ცვლილება არსებით მნიშვნელობას იძენს. ნეიტრალური ევოლუციის თვალთახედვით, გენთა დრეიფი მნიშვნელოვან როლს ასრულებს დიდი რიცხოვნობის მქონე პოპულაციაში. კერძოდ, მუტაციის შედეგად წარმოქმნილი სელექციურად ნეიტრალური ვარიანტების სიხშირე პოპულაციაში გენთა დრეიფის მოქმედებით მნიშვნელოვნად იცვლება. შეზღუდული სიდიდის პოპულაციაში რიცხოვნობა ისე შეიძლება დაეცეს, რომ გენთა დრეიფმა სელექციურად მნიშვნელოვან ლოკუსებზე ან გენთა ნაკადზეც კი არსებითი გავლენა მოახდინოს. ზოგიერთ პოპულაციაში ზემოთ განხილული მოვლენების შედეგად რიცხოვნობა მკვეთრად ფლუქტუირებს. მრავალი სახეობის პოპულაციაში პოპულაციური ტალღები მოქმედებს, რომელიც „ბოთლის ყელის“ ეფექტს წარმოქმნის. დგება პერიოდი, რომლის დროსაც პოპულაციაში რიცხოვნობა და სიმჭიდროვე მინიმუმამდეა შემცირებული (იხ.: სურ 7.3). შემთხვევით გადარჩება პოპულაციის შემადგენელი ინდივიდების მცირე რაოდენობა, დიდი ნაწილი კი ელიმინირდება. პოპულაციის „ბოთლის ყელში“ გასვლის შემდეგ მასში რიცხოვნობა სწრაფად მატულობს.

სურ. 7.3. პოპულაციაში მუტაციის კონცენტრაციის შემთხვევითი მერყეობის სქემა A, B, C, D, E, F - სხვადასხვა გენები, რომელთა ერთობლიობა წარმოადგენს ინდივიდს, A, B, C, D, E, F - შესაბამისი მუტაცია, რომელიც პოპულაციაში, სხვადასხვა კონცენტრაციითაა წარმოდგენილი, თუ რიცხოვნობის ცვლილების შედეგად მხოლოდ კვადრატში მოთავსებული ჯგუფები გადარჩებიან (მთელი ერთობლიობის 10%25-ს შეადგენს) მუტაციის კონცენტრაცია პოპულაციაში მკვეთრად შეიცვლება (ბაუერით, ნ. ტიმოფეევ-რუსოვსკიდან 1943).
როდესაც პოპულაციის რიცხოვნობა მკვეთრად ეცემა, შემთხვევით ინდივიდთა მცირე რაოდენობა გადარჩება. ასეთი პოპულაციის გენური სიხშირე საწყისისაგან განსხვავებული იქნება. თუ შემდგომში რიცხოვნობა კვლავ გაიზრდება, მაშინ ახალ აფეთქებას დასაბამს მისცემს ინდივიდთა ეს მცირერიცხოვანი ჯგუფი. ამდაგვარი პოპულაციის გენოფონდს ამ მცირე ჯგუფის გენეტიკური შემადგენლობა განსაზღვრავს. რაც უფრო მცირერიცხოვანია ინდივიდთა ამონაკრები, მით უფრო მაღალია ალელთა სიხშირის მერყეობა. ალელთა ეს მერყეობა (გენთა დრეიფი) შემთხვევითია და გადარჩევის შედეგს არ წარმოადგენს. როდესაც პოპულაცია ბუნებრივ გადარჩევას ექვემდებარება, მაშინ იმავე მიმართულებით მიმდინარე ნებისმიერი შემთხვევითი ცვლილება გადარჩევის ეფექტურობას ზრდის ე.ი. აძლიერებს ან აჩქარებს. პირუკუ, გადარჩევის საწინააღმდეგოდ მიმართული ნებისმიერი შემთხვევითი ცვლილება ანელებს გადარჩევას. გენთა დრეიფი არამიმართული პროცესია, ამდაგვარმა პერეტრუბაციამ შეიძლება გამოიწვიოს ადაპტური ღირებულების მქონე ალელების ელიმინაცია და მოხდეს ნაკლები ადაპტური მნიშვნელობის ალელების ფიქსაცია. ამ შემთხვევაში ჩნდება შეუქცევადი ცვლილება (თუ დაკარგული ალელი მუტაციის შედეგად კვლავ არ ინდუცირდა). თუ გენთა დრეიფი შეგუებულობას მკვეთრად აქვეითებს, მაშინ ასეთი ინდივიდები შეიძლება ამოწყდეს. ამრიგად, გენთა დრეიფის შედეგად წარმოქმნილი ყველა ცვლილება როდი შენარჩუნდება. მაგრამ ზოგჯერ წარმოიქმნება მეტად შეგუებული ინდივიდები. ე.მაირმა ამ შემთხვევას „გენეტიკური რევოლუცია“ უწოდა. ზოგიერთი ევოლუციონისტი მიიჩნევს, რომ ახალი სახეობა სწორედ ამდაგვარი პროცესის შედეგად ყალიბდება. მაგრამ ამ მოსაზრებას მკვლევართა დიდი ნაწილი არ იზიარებს.
თუ რატომაა ეფექტური მცირერიცხოვან პოპულაციებში გენთა დრეიფი, ამის საილუსტრაციოდ განვიხილოთ შემდეგი მაგალითი: დავუშვათ, რომ ორი განსხვავებული რიცხოვნობის პოპულაციაში გენთა სიხშირეები თანაბარია და იშვიათი ალელი ინდივიდთა 1%25-ში გვხვდება (Q=0,01). შემთხვევითი მოვლენის შედეგად მოხდა ამ ალელის მქონე ინდივიდთა ელიმინაცია პოპულაციიდან, ე.ი. Q=0. თუ პოპულაცია 1.000.000 წევრისაგან შედგება, მაშინ იშვიათი ალელი 10.000 ინდივიდს ექნება. ხოლო თუ მეორე პოპულაცია 100 წევრისაგან შედგება, მაშინ ალელი მხოლოდ 1 ინდივიდს აღმოაჩნდება. ბუნებრივია ალელის შემთხვევითი დაკარგვის ალბათობა მცირერიცხოვანი (მეორე შემთხვევა) პოპულაციიდან გაცილებით მაღალია, ვიდრე მრავალრიცხოვანიდან.
გენთა დრეიფთანაა დაკავშირებული ე.წ. დამფუძნებლის პრინციპი (ე.მაირი, 1974). ამ მოვლენას ვხვდებით იმიგრაციის შედეგად ახალი პოპულაციის ფორმირებისას. როგორც უკვე აღინიშნა, საწყისი პოპულაციიდან შემთხვევითი ამორჩევით ინდივიდთა ძალზე მცირე ჯგუფი შეიძლება განსახლდეს და სხვა ტერიტორია დაიჭიროს. ბუნებრივია, ინდივიდთა ეს მცირერიცხოვანი ამონაკრები ალელთა შედგენილობით არატიპურია და განსხვავდება საწყისი პოპულაციისაგან. ზოგიერთი ალელის კონცენტრაცია მაღალი იქნება, ზოგიერთი კი მათში საერთოდ არ აღმოჩნდება. ასორტატული შეჯვარების შედეგად ასეთ პიონერულ პოპულაციაში ყალიბდება ახალი გენოფონდი, რომელიც საწყისისაგან ალელთა კონცენტრაციითაა განსხვავებული. გენთა დრეიფით, იშვიათ ალელთა დაკარგვის გამო წესისამებრ, გენეტიკური ცვალებადობა ქვეითდება. ხანგრძლივი ასორტატული შეჯვარება ამცირებს პოპულაციაში ჰეტეროზიგოტების რაოდენობას, მკვეთრად ზრდის ჰომოზიგოტების წილს. ამრიგად, პოპულაცია მეტად დაბალი პოლიმორფულობით ხასიათდება. ახალი პოპულაცია სათავეს ამდაგვარი გაღარიბებული გენოფონდიდან იღებს. პოპულაციის დინამიკაზე მოქმედი ფაქტორების (მუტაციური პროცესი, ბუნებრივი გადარჩევა და სხვ.) ზემოქმედებით იცვლება გენთა კონცენტრაცია გენოფონდში და იგი პოლიმორფული ხდება. საწყისი განსაზღვრული გენოფონდის გავლენას პოპულაციის შემდგომ ბედზე დამფუძნებლის პრინციპი ეწოდება.
ოკეანის კუნძულებზე დასახლებული მრავალი სახეობა, რომლებიც ამჟამად დიდი რიცხოვნობით გამოირჩევა, ოდესღაც შემთხვევით მოხვედრილი ერთი (განაყოფიერებული მდედრი, ჰერმაფროდიტი ორგანიზმი, თესლი და სხვ.) ან რამდენიმე ინდივიდისაგან იღებს სათავეს. დამფუძნებლის ეფექტი გამოვლენილია ადამიანის მრავალ მცირერიცხოვან პოპულაციაში.
აშშ-ს პენსილვანიის შტატის ქალაქ ლანკასტერში სახლობს რელიგიური იზოლატი (დახშული პოპულაცია), ქორწინება მხოლოდ პოპულაციის წევრებს შორის ხდება. პოპულაციაში მაღალი სიხშირით გვხვდება ელისა - ვან კრეველდის (EVC) სინდრომი. დამახასიათებელია განსაკუთრებული ფორმის ჯუჯა სხეული და პოლიდაქტილია. ნიშანი რეცესიულია და ჰომოზიგოტებს უყალიბდებათ. რელიგიურ იზოლატს სამმა ცოლქმრულმა წყვილმა დაუდო სათავე, რომლებიც 1770 წელს ევროპიდან ამერიკაში გადასახლდნენ. პოპულაციის 40 საგვარტომოდან, რომელიც 13000 ადამიანს მოიცავს, 82 სინდრომის მატარებელია. ინბრიდინგის გათვალისწინებით გამოთვლილია რეცესიული გენის საშუალო სიხშირე, რომელიც დაახლოებით 0,066 შეადგენს, ხოლო დაავადების სიხშირე - 0,005-ის ტოლია. თემის დამფუძნებელი ცოლქმრული წყვილებიდან მხოლოდ ერთი - სამუელ კინგი, ან მისი მეუღლე იყო ჰეტეროზიგოტი. ამ მონაცემებიდან ირკვევა, რომ რელიგიურ იზოლატში დაავადების მაღალი სიხშირე დამფუძნებლის ეფექტითაა გამოწვეული (ფ. ჰედრიკი, 2004).
იშვიათ ან გაქრობის პირას მისულ სახეობებში მცირერიცხოვან პოპულაციებს დიდი მნიშვნელობა ენიჭება. უსისტემო ნადირობის შედეგად ზღვის სპილო (irounga leonina) XIX საუკუნეში მთლიანად ამოწყდა. კუნძულ გვადალუპეს (მექსიკა) ზოგიერთ პლაჟზე შემთხვევით 20-მდე ინდივიდი გადარჩა. დაცვის უმკაცრესი ზომების შედეგად ზღვის სპილოს რაოდენობამ ამჟამად 100.000 ინდივიდს მიაღწია, რომელიც სათავეს ამ მცირერიცხოვანი კოლონიიდან იღებს.
მრავალი სახეობა, როგორიცაა პრჟევალსკის ცხენი, კალიფორნიული კონდორი, მექსიკური მგელი გადაშენების პირასაა მისული. ამ სახეობის ამჟამად მცხოვრები ინდივიდები ხელოვნურ პირობებშია გამრავლებული და წარმოშობილია მცირერიცხოვანი ჯგუფიდან. მაგალითად, პრჟევალსკის ცხენი 1900 წელს დაჭერილი 13 ინდივიდიდანაა მიღებული, კონდორი კი 1970 წელს მექსიკაში დაჭერილი 14 ფრინველიდანაა წარმოშობილი. ბუნებრივია, ყველა ჩამოთვლილ შემთხვევაში დამფუძნებლის ფენომენი მოქმედებს.
გენეტიკური დრეიფის თეორია ბუნებრივ და ხელოვნურ პოპულაციებში ჩატარებული გამოკვლევების საფუძველზე არაერთხელ გადაისინჯა. ამ მოვლენის აღმომჩენები ს. რაიტი და ნ. დუბინინი თავდაპირველად გენთა დრეიფს თითქმის ისეთსავე მნიშვნელობას ანიჭებდნენ, როგორიც ბუნებრივ გადარჩევას აქვს. მოგვიანებით თვით ავტორებმა შეიცვალეს პოზიცია. ევოლუციურ თეორიასა და პოპულაციის გენეტიკაში დამკვიდრდა შეხედულება, რომ მიკროევოლუციაში პრიმატი სელექციურ ფაქტორებს ენიჭება, ხოლო სტოქასტური პროცესები მეორეხარისხოვან როლს ასრულებენ. XX საუკუნის 60-იან წლების შემდეგ გენეტიკური დრეიფის ევოლუციური როლის შესახებ დისკუსია გამწვავდა, კერძოდ, ცილების მაღალი პოლიმორფულობის გამოვლენასთან დაკავშირებით წამოაყენეს ნეიტრალური ევოლუციის კონცეფცია (იხ.: §14.1). ამ კონცეფციის მიხედვით, მოლეკულურ დონეზე ევოლუციურ პროცესში წამყვან როლს გენთა დრეიფი ასრულებს. საზოგადოდ, გენთა დრეიფი და ინბრიდინგი ჰეტეროზიგოტულობაზე მსგავს მოქმედებას ავლენენ, საბოლოო მოსალოდნელი შედეგებით კი - განსხვავდებიან.
გენეტიკური დრეიფის როგორც ევოლუციური ფაქტორის როლი ჯერ კიდევ არ შეიძლება ჩაითვალოს საბოლოო გადაწყვეტილად და შემდგომ შესწავლას საჭიროებს. გენთა დრეიფი არის შემთხვევითი, არამიმართული პროცესი და მუტაციურ პროცესთან ერთად ელემენტარული მასალის ერთგვარ „მიმწოდებელ” ფაქტორს წამოადგენს.
|
|
9.3 7.3. გენთა ნაკადი როგორც ევოლუციის ელემენტარული ფაქტორი |
▲ზევით დაბრუნება |
ნებისმიერი სახეობა არის რთული, მრავალსაფეხურიანი იერარქიული სისტემა. მასში გაერთიანებულია სხვადასხვა შიდასახეობრივი კატეგორიები, როგორებიცაა ქვესახეობა, ეკოტიპი, რასა, დემი, პოპულაცია და ა.შ. (იხ.: §11.4). სახეობის ელემენტარულ-სტრუქტურულ ერთეულს პოპულაცია წარმოადგენს. ამასთანავე, სახეობა წარმოადგენს ინტეგრირებულ სისტემას. სახეობის შემადგენელ სტრუქტურებს შორის ინტეგრაცია გენთა ნაკადით მყარდება. გენთა ნაკადი მიეკუთვნება პოპულაციის გენეტიკური სტრუქტურის დინამიკაზე მოქმედ ფაქტორს. პოპულაციათა შორის გენთა მიმოცვლას, გენთა ნაკადი ეწოდება. ერთი პოპულაციიდან მეორეში ინდივიდთა მცირე ჯგუფის იმიგრაცია იწვევს, მასში განსხვავებულ ალელთა შემთხვევით მოხვედრას. იმის გამო, რომ ერთ პოპულაციას ალელთა გარკვეული ფორმები აკლდება, მეორეს კი ემატება, თითოეულის გენოფონდში ალელთა ხვედრითი პროპორციები იცვლება. გენთა ნაკადის მეშვეობით პოპულაციაში ცვალებადობის დონე იზრდება. ამ პროცესის უშუალო შედეგი მუტაციების წარმოქმნის პროცესის მსგავსია. გენთა ნაკადი ალელთა კონცენტრაციის გაცილებით სწრაფ ცვლილებას იწვევს, ანუ მუტაციურ პროცესზე უფრო ეფექტურია. მართალია, გენთა ნაკადით გენოფონდი იცვლება, მაგრამ ევოლუციური ცვლილების თვალსაზრისით, მისი მოქმედება კონსერვატულია. პოპულაციათა შორის მიგრაციების მეშვეობით სახეობის გენოფონდი ერთიანი ხდება. ამრიგად, გენთა ნაკადი, ერთი მხრივ პოპულაციათა გენეტიკური ცვალებადობის წყაროა, მეორე მხრივ, არსებით როლს ასრულებს სახეობის, როგორც ევოლუციის ეტაპის, მდგრადობის შენარჩუნებაში. ამდენად, პოპულაციათა შორის გენთა ნაკადის შეწყვეტა (საიზოლაციო მექანიზმების განმტკიცება) ახალ სახეობათა ფორმირების ერთ-ერთი წინაპირობაა.
სახეობაში გაერთიანებულ სტრუქტურების ერთმანეთთან კავშირის ხარისხი მათ შორის მიმდინარე გენთა ნაკადის ინტენსივობაზეა დამოკიდებული. როდესაც ჯგუფებს შორის გენთა ნაკადის ინტენსივობა მაღალია, გენთა ნაკადი იწვევს სახეობის შემადგენელ სტრუქტურებს შორის არსებულ გენეტიკური ცვალებადობის ჰომოგენიზაციას (ნიველირებას). იმ შემთხვევაში, როცა გენთა ნაკადის ინტენსივობა სუსტია, მაშინ გენთა დრეიფით, გადარჩევითა და მუტაციური ზეწოლით შესაძლებელია მოხდეს ცალკეულ ჯგუფთა გენეტიკური დიფერენცირება.
ზოგიერთი ტიპის მიგრაცია (მაგალითად, ფრინველების სეზონური მიმოფრენა) (იხ.: §3) პოპულაციათა შორის გენთა მიმოცვლას არ იწვევს. ინდივიდთა გადაადგილება ან გავრცელება სარჩოს საძიებლად, რომლის დროსაც შეჯვარება და რეპროდუქცია არ ხდება, ასევე არაეფექტურ გენთა ნაკადს წარმოადგენს. სქესმწიფე ინდივიდების იმიგრაცია სხვადასხვა სტრუქტურებში, რომელთაც წვლილი არ შეაქვთ ახალი გენოფონდის ფორმირებაში, გენთა ნაკადს არ წამოადგენს. ენდლერმა ერთიმეორისაგან გამიჯნა გენთა ნაკადი, მიგრაციები და ინდივიდთა გავრცელება. სახეობის შემადგენელი სტრუქტურების ერთიან გენეტიკურ სტრუქტურად - გენოფონდად ჩამოყალიბებას მხოლოდ გენთა ნაკადი განაპირობებს.
გენთა ნაკადის ინტენსივობა განსაზღვრულია ადამიანის ისეთ პოპულაციაში, რომელიც რამდენიმე პოპულაციიდან არის ჩამოყალიბებული. აშშ-ში თეთრკანიანებსა (კავკასოიდური რასა) და ზანგებს შორის შერეულ ქორწინებათა შედეგად გაჩენილ შთამომავლებს (მულატი) ზანგებს აკუთვნებენ. ამდენად, შერეული ქორწინებები შეიძლება ჩაითვალოს როგორც გენთა ნაკადი თეთრკანიანებიდან ზანგთა პოპულაციაში. აფრო-ამერიკელებში გენთა ნაკადის განსასაზღვრავად სისხლის ჯგუფობრიობის ლოკუსებს იყენებენ. მაგრამ გამოირკვა, რომ პოპულაციებში მარკერებად გამოყენებული ლოკუსების სიხშირე ბუნებრივი გადარჩევის მოქმედებით და გენთა დრეიფით იცვლება. ამდენად, ამ მაჩვენებელზე დაყრდნობით მიღებული შედეგები სარწმუნო არ არის. ამერიკელმა მკვლევარმა პარამ, თანამშრომლებთან ერთად (1998), ადამიანში 10 პოპულაციურ-სპეციფიკური ალელი გამოავლინა, რომლის გამოყენებითაც განსაზღვრა გენთა ნაკადის ინტენსივობა. 7.2 ცხრილში ნიმუშის სახით მოტანილია მხოლოდ ოთხი ლოკუსის მიხედვით მიღებული შედეგები. ასევე, მოცემულია ათივე ლოკუსის ანალიზის შედეგად მიღებული გასაშუალოებული (მ) მონაცემები. მკვლევრებმა ანალიზისთვის შეარჩიეს ისეთი ორალელიანი ლოკუსები, რომელთა რესტრიქციულ საიტთაგან ზოგიერთი პოლიმორფულია, ხოლო ზოგიც - არა. სპეციფიკური ალელური სიხშირით ხასიათდებოდა FY-NULL მარკერი. ის მხოლოდ გამოკვლეულ კავკასოიდური რასის პერსონებს მოეპოვებოდათ.
ცხრილი 7.2.
პოპულაციურ-სპეციფიკური ალელების სიხშირე ნეგროიდულ,
კავკასოიდურ და აშშ აფრო-ამერიკულ პოპულაციაში
(პარა და თანაავტ. 1998)
პოპულაცია |
- |
C 2 |
R2300 |
G-I |
მ |
ნეგროიდები
კავკასოიდები
აფრო-ამერი-
მეივუდი
ნიუ-იორკი
ფილადელფია
ჩარსტოუნი
ნიუ-ორლეანი
იამაიკა |
0.000
1.000
0.185
0.210
0.160
0.112
0.200
0.065 |
0.098
0.769
0.203
0.220
0.137
0.208
0.284
0.091 |
0.920
0.333
0.776
0.821
0.802
0.888
0.842
0.870 |
0.824
0.156
0.710
0.738
0.771
0.765
0.669
0.790 |
0.188±
0.198±
0.138±
0.116±
0.225±
0.068± |
გენთა ნაკადის დაბალი ინტენსივობა შეინიშნებოდა იამაიკისა და ჩარლსტოუნის (სამხრეთი კაროლინა) ერთობლიობებში. მიაჩნიათ, რომ აშშ-ის სამხრეთ პოპულაციების გენოფონდის ფორმირებაში კავკასოიდური რასის ხვედრითი წილი უმნიშვნელოა. ნიუ-ორლეანის ნაკრებში შეინიშნებოდა გენთა ნაკადის ყველაზე მაღალი ინტენსივობა. ამდენად, კავკასოიდური რასის ელემენტების წილი გენოფონდის ფორმირებაში საკმაოდ მნიშვნელოვანია. ამავე მკვლევრებმა განსაზღვრეს დედისა და მამის ხაზით (მტ-დნმ-სა და Y-ქრომოსომების ვარინტების მიხედვით) გადაცემული გენთა ნაკადის ინტენსივობა. მიტოქონდრიული დნმ-ს მარკერების მიხედვით გენთა ნაკადის სიდიდემ 0,140 შეადგინა, ხოლო სასქესო Y-ქრომოსომული მარკერების მიხედვით - 0,248. კავკასოიდური რასის მდედრობითი და მამრობითი სქესის განსხვავებული თანაფარდობით მონაწილეობა გენოფონდის ფორმირებაში ისტორიულად ჩამოყალიბდა. კერძოდ, ამერიკელი თეთრკანიანი მამაკაცებისა და შავკანიანი ქალების ქორწინება გაცილებით ჭარბობს თეთრკანიანი ამერიკელი ქალებისა და შავკანიანი მამაკაცების ქორწინებებს.
მე-17 საუკუნის დასასრულიდან (დაახლოებით, 300 წელი) აფრიკის მკვიდრი მოსახლეობის დიდი ნაწილი მონებად შემოჰყავდათ ამერიკაში. დავუშვათ, რომ გენთა ნაკადის ინტენსივობა ამ დროიდან მოყოლებული დღემდე ერთნაირი იყო, ხოლო თაობის საშუალო ხანგრძლივობა კი - 25 წელი. მაშინ 300 წლის განმავლობაში თაობათა რაოდენობა, რომლის პერიოდშიც გენთა ნაკადი მიმდინარეობდა, 12 თაობას შეადგენს. გამოთვლილია, რომ ნიუ-ორლეანის ზანგებს გენოფონდში გენთა დაახლოებით 21%25 თეთრკანიანი პოპულაციიდან, გენთა ნაკადის მეშვეობით აქვთ მიღებული, ჩარლსტოუნში მოსახლე ზანგებს კი - მხოლოდ 10%25.
გენთა ნაკადის ინტენსივობას პოპულაციაში ზოგჯერ უშუალოდ, სათანადო მეთოდების გამოყენებით საზღვრავენ. პოპულაციაში ახდენენ ინდივიდების იდენტიფიცირებას. სანიმუშოდ მოვიყვანთ სათანადო მაგალითს, რომლითაც საბუთდება პოპულაციის შემადგენელი წევრებისა და გენთა ნაკადის შეფარდებითი როლი შემდგომი გენოფონდის ფორმირებაში. განიოსმა და თანაავტორებმა (1997) გამოკვლევები ჩაატარეს დასავლეთ აფრიკაში გავრცელებულ შიმპანზეს სუბპოპულაციაში პოტენციური მამის გამოსავლენად. გენოტიპის დადგენას მიკროსატელიტური დნმ-ის იდენტიფიცირებით ახდენდნენ. გამოსაკვლევი სუბპოპულაცია 52 ინდივიდისაგან შედგებოდა. შიმპანზეს ჯოგისთვის ნიშანდობლივია გამოკვეთილი იერარქიული სტრუქტურა. მას დომინანტი ლიდერი მამრი უდგას სათავეში. მამობის დასადგენად ანალიზი 14 დედა-შვილსა და 6 პოტენციურ მამაში (მათ შორის ლიდერში) ჩაატარეს. წესისამებრ სუბპოპულაციის წევრები წყვილდებიან. უმეტეს შემთხვევაში ლიდერ მამრს მონოპოლიზებული აქვს მდედრებთან შეწყვილება. ზოგჯერ ეს პროცესი შემთხვევით ხასიათს ატარებს და ჯოგში გაერთიანებული მამრები ეწყვილებიან მენზისურ (ესტრუსულ) მდგომარეობაში მყოფ მდედრებს. ჩატარებული მოლეკულურ-გენეტიკური ანალიზით დაადგინეს, რომ 7 ნაშიერის მამა ჯოგის წევრი იყო, ხოლო დანარჩენი 7 ნაშიერის კი - სხვა ჯოგის მამრები. ამრიგად, 14 ნაშიერიდან შვიდმა გენები სხვა ჯოგში მცხოვრები მამრებისაგან მიიღო. შესაბამისად, გენთა ნაკადი M=(0,00+0,50)/2=0,25-ს. ყველა სქესმწიფე მდედრი პერიოდულად თავის ჯოგს ტოვებს. გამოირკვა, რომ ის მდედრები, რომლებიც იმავე ჯოგის წევრებს ეწყვილებიან, გაცილებით მცირე ხნით ტოვებენ ჯოგს, ვიდრე ისინი, ვინც სხვა ჯოგის მამრებთან ამყარებენ სქესობრივ კონტაქტებს.
გენთა ნაკადი არამიმართული და შემთხვევითი პროცესია. გენთა ნაკადის გავლენა პოპულაციის დინამიკაზე დამოკიდებულია როგორც პოპულაციათა შორის სივრცობრივ მანძილზე, ისე გამეტების და ინდივიდების გავრცელების რაოდენობასა და სიხშირეზე. მაგალითად, მფრინავი ცხოველები, მტვრის მარცვლები, თესლები აქტიურად ან პასიურად სწრაფად და ადვილად ვრცელდებიან სხვა ადგილებში. გენთა ნაკადის გენეტიკური შედეგი ისაა, რომ თუ ემიგრანტების გენოტიპური თავისებურებანი ახალ პირობებში მათ ადაპტურ უპირატესობას მიანიჭებს, ისინი გამრავლდებიან, თავიანთ წვლილს შეიტანენ პოპულაციის გენოფონდში და ამით გავლენას მოახდენენ პოპულაციის გენეტიკურ სტრუქტურაზე. ხოლო თუ, მათი ადაპტური ღირებულება დაბალი აღმოჩნდება, ისინი პოპულაციიდან ელიმინირდებიან.
|
|
9.4 7.4. ინბრიდინგი - პოპულაციის დინამიკაზე მოქმედი ფაქტორი |
▲ზევით დაბრუნება |
ჰარდი-ვაინბერგის (პოპულაციაში ალელთა წონასწორობის) კანონის მოქმედების ერთ-ერთი მნიშვნელოვანი პირობაა უსასრულოდ მრავალრიცხოვან წევრთა თავისუფალი შეჯვარება (პანმიქსია). პოპულაციაში ინდივიდთა სასრული რაოდენობის არსებობა კიდევ ერთ მნიშვნელოვან შედეგს ინბრიდინგს იწვევს. ერთი პოპულაციის ინდივიდთა ახლონათესაურ შეჯვარებას ინბრიდინგი ეწოდება. რადგან პოპულაცია სასრული რაოდენობის წევრებისაგან შედგება და დაყოფილია შიგაპოპულაციურ სტრუქტურებად (სუბპოპულაციებად), ამიტომ მიმდინარეობს ასორტატული ანუ შერჩევითი შეჯვარება. ადამიანთა საზოგადოებაც კი ოჯახური და ტომობრივი კავშირების, გეოგრაფიული ფაქტორების, ეროვნების, ასევე კულტურული, რელიგიური, სოციალური და ეკონომიკური ფაქტორების შედეგად ფენებადაა დაყოფილი. ერთსა და იმავე ფენის წევრებს შორის ქორწინება გაცილებით მაღალი სიხშირით მიმდინარეობს, ვიდრე სხვადასხვა ფენების წევრებს შორის. ყოველივე ზემოთ აღნიშნული ასორტატული ანუ შერჩევითი შეჯვარების ალბათობას ზრდის.
ადამიანთა შორის ნათესაობა გაცილებით მაღალია, ვიდრე ეს ერთი შეხედვით შეიძლება მოგვეჩვენოს. განვიხილოთ კონკრეტული მაგალითი: თითოეულ ჩვენგანს ორი მშობელი ჰყავს, ბებია-პაპების რაოდენობა ოთხია, დიდი ბებია-პაპების - რვა და ა.შ. თუ თვალს გავადევნებთ წარსულს, თითოეულ ჩვენგანს ჰყავს 2ნ წინაპარი. ბოლო 20 თაობის მანძილზე თითოეული ჩვენგანის წინაპრების რაოდენობა 220=700.000 შეადგენს. დავუშვათ, რომ ყოველ საუკუნეში ადამიანის თაობათა რაოდენობა ოთხია, მაშინ ოცი თაობისათვის ხუთი საუკუნეა საჭირო. შემდეგი მსჯელობიდან ნათელია, თუ რამდენად რეალურია აბსოლუტურად არანათესაური ქორწინებები. თუ დავუშვებთ, რომ აშშ მთელი მოსახლეობის გარკვეულ ნაწილს არა აქვს ერთმანეთთან ნათესაური კავშირი (ყოველი 20 ადამიანიდან ერთი, ხოლო წარსულში წინაპართა ნათესაურად სრულიად დამოუკიდებელი ჯგუფები ჰყავდათ), მაშინ XV საუკუნეში ქვეყნის მოსახლეობა 3 ტრილიონი უნდა ყოფილიყო. ეს რიცხვი 500-ჯერ აღემატება პლანეტის ამჟამინდელ მოსახლეობას.
ცხრილი 7.3.
პოპულაციაში გენოტიპთა თანაფარდობა
თვითგანაყოფიერებით (ინბრიდინგი) გამრავლებისას
თაობა |
გენოტიპი |
||
|
AA |
A |
AA |
0 1 2 3 4 5 10 N |
- 1 3 7 15 31 1023 2ნ-1 |
1 2 2 2 2 2 2 2 |
- 1 3 7 15 31 1023 2ნ-1 |
პოპულაციის გენოტიპურ სტრუქტურაზე ინბრიდინგის მოქმედება განვიხილოთ ალელთა ერთნაირი სიხშირის მქონე პოპულაციაზე P=Q=0,5. ასეთ პოპულაციაში გენოტიპების თანაფარდობაა: 0,25AA:0,50A:0,25AA. ნულოვან თაობაში შთამომავლობის ნახევარი ჰეტეროზიგოტულია (0,50 Aა), მეორე ნახევარი * ჰომოზიგოტური (0,25AA+0,25AA). თუ ამის შემდგომ გამრავლება მხოლოდ თვითგანაყოფიერებით განხორციელდება, ყოველ მომდევნო თაობაში ჰეტეროზიგოტულ მდგომარეობაში მყოფი ალელების ნახევარი გადადის ჰომოზიგოტულ მდგომარეობაში. ამრიგად, ჰეტეროზიგოტების წილი ყოველ მომდევნო თაობაში ნახევრდება, რაც პრაქტიკულად იწვევს მთლიან ჰომოზიგოტიზაციას (იხ. ცხრილი 7.3). ზოგადი სახით მოსალოდნელი ჰეტეროზიგოტების რაოდენობა ნ თაობაში შეადგენს 2PQX (1/2)ნ; სადაც 2PQ- ჰეტეროზიგოტების ხვედრითი წილია ნულოვან თაობაში, რომლიდანაც დაიწყო თვითგანაყოფიერება. ინბრიდინგის დროს ჰომოზიგოტურ მდგომარეობაში გადადიან და ვლინდებიან რეცესიული გენები. მათ შორის არიან ისეთებიც, რომლებიც ორგანიზმის ცხოველმყოფელობაზე მოქმედებენ და მის დაკნინებას, ე.წ. ინბრიდულ დეპრესიას იწვევენ.
ინბრიდინგი პოპულაციაში იწვევს რამდენიმე შედეგს, კერძოდ:
1. ჰომოზიგოტურობის გაზრდას*
2. რეცესიული ალელების გამოვლენას*
3. რეცესიული ალელების უარყოფითი ეფექტის დროს გამოვლინდება ე.წ. ინბრიდული დეპრესია*
4. ჰომოზიგოტურ მდგომარეობაში მრავალი ალელის გადასვლა იწვევს ფენოტიპური ცვალებადობის ამაღლებას.
|
|
9.5 7.5. იზოლაცია როგორც ევოლუციის ელემენტარული ფაქტორი |
▲ზევით დაბრუნება |
იზოლაცია ევოლუციის მნიშვნელოვან ელემენტარულ ფაქტორს წარმოადგენს. შიდასახეობრივი იზოლაცია არის პოპულაციების ნებისმიერი ბარიერით გამიჯვნა, რომლის შედეგადაც წყდება გენთა ნაკადი. ევოლუციის პროცესში იზოლაციის მნიშვნელობა ცხადყო ჩ. დარვინმა. ოკეანური კუნძულების ფლორისა და ფაუნის შესწავლის საფუძველზე მან დაადგინა, რომ იზოლაციამ შეიძლება გამოიწვიოს მონათესავე ორგანიზმთა შორის სხვაობის გაჩენა, მათი თანდათანობითი გაღრმავება და ბოლოს ბუნებრივი გადარჩევის მოქმედებით ახალი სახეობების ფორმირება. ჩ.დარვინის ამ განზოგადებულ დასკვნას მნიშვნელობა ამჟამადაც არ დაუკარგავს.
მიკროევოლუციის პროცესში იზოლაციის როლი XXს. 30-40-იან წლებში შეისწავლეს და განისაზღვრა მისი, როგორც ევოლუციის ელემენტარული ფაქტორის, მნიშვნელობა. ტერმინი „საიზოლაციო მექანიზმები“ შემოთავაზებულია ფ.დობჟანსკის მიერ. როდესაც პოპულაციები ან პოპულაციის გარკვეული ნაწილი თაობათა მანძილზე (ხანგრძლივად) რჩება გამიჯნული, შეიძლება მოხდეს გენეტიკური სტრუქტურით მათი დივერგირება ან დიფერენცირება. ეს განსაკუთრებით მაშინაა შესაძლებელი, როდესაც ბუნებრივი გადარჩევა გამიჯნულ პოპულაციებზე განსხვავებული მიმართულებით მოქმედებს. ამდაგვარი პოპულაციების დიფერენცირებამ შესაძლოა ახალ სახეობათა ფორმირება გამოიწვიოს. იზოლაცია გამოიხატება პოპულაციებსა ან მათ გარკვეულ ნაწილთა შორის ამა თუ იმ სახის ბარიერის არსებობით. ბუნებაში ბარიერთა ხასიათის შესაბამისად იზოლაციაც განსხვავებულია. გამოყოფენ იზოლაციის ორ ძირითად ფორმას: სივრცობრივსა და ბიოლოგიურს.
სივრცობრივი (ტერიტორიული) იზოლაცია გულისხმობს პოპულაციათა გამიჯვნას გეოგრაფიული ზღუდეებით (მდინარე, სრუტე, ხრამი, მთები და სხვ.) იგი შეიძლება სხვადასხვა ფორმით განხორციელდეს. ხმელეთის ბინადართ მიჯნავს წყლის ბარიერი, ჰიდრობიონტებს კი - ხმელეთის ბარიერი. დაბლობის პოპულაციებს - მთები, მთის პოპულაციებს კი დაბლობი და ა.შ. სივრცობრივი იზოლაცია ასევე შეიძლება ეკოლოგიური ფაქტორებით (მაგ., არახელსაყრელი ფიზიკურ-ქიმიური პირობები) ან გაუვალი ეკოცენოზებით (მაგ., ტყის მასივები, უდაბნოები და სხვ.) მოხდეს.
სურ. 7.4. xlakunas isgurnus fossilis (1) და ცისფერი კაჭკაჭის Cyanopicca cyana (2) წყვეტილი არეალი (სხვადასხვა ავტორების მონაცემებით, ლ. კაიდანოვი, 1996).
ვულკანური წარმოშობის (ოკეანურ) პატარა კუნძულ ოახუზე ხმელეთის ლოკოკინას, გვარ chatinella-ში გაერთიანებული 25 ენდემური სახეობაა გავრცელებული. კუნძულზე ერთიმეორისაგან გამიჯნული, ტროპიკული ტყით დაფარული 25 დაბლობია, თითოეულ მათგანზე ცალკეული სახეობა ბინადრობს. სივრცობრივი იზოლაციის არსებობა ზოგ შემთხვევაში გარკვეულ ტერიტორიაზე სახეობის ისტორიული განვითარებით შეიძლება აიხსნას. მაგალითად, ცისფერ კაჭკაჭს (Cyanopica cyana) წყვეტილი (დიზუნქციური) არეალი აქვს. იგი გვხვდება როგორც პირინეის ნახევარკუნძულზე, ასევე შორეულ აღმოსავლეთში. მსგავს წყვეტილ არეალს ფლობს თევზი ხლაკუნა (isgurnus fossilis), რომელიც გავრცელებულია დასავლეთ ევროპასა და შორეულ აღმოსავლეთში. ორივე სახეობის გამიჯვნა გამყინვარებითაა გამოწვეული (იხ.: სურ. 7.4). ხანგრძლივი ბიოლოგიური დროის გასვლის მიუხედავად გამიჯნულ პოპულაციებში არსებითი მორფო-ფიზიოლოგიური ცვლილებები არ მომხდარა. ისინი ერთსა და იმავე სახეობას მიეკუთვნებიან. სივრცობრივი იზოლაციის მიზეზად რიგ შემთხვევაში ანთროპოგენური ფაქტორი გვევლინება. ტიპურ მაგალითს ევრაზიაში გავრცელებული სიასამურის (artes zibellina) წყვეტილი არეალი წარმოადგენს. იგი XX ს. დასაწყისში ცხოველის უსისტემო რეწვამ გამოიწვია (იხ.: სურ. 7.5.). სივრცობრივი იზოლაცია შეიძლება გამოიწვიოს როგორც გეოგრაფიული ზღუდით გამიჯვნამ, ისე სხვადასხვა მიზეზით გამოწვეულმა შორი მანძილით განსახლებამ.
სურ. 7.5. სიასამურის (artes zibellina) არეალის სივრცობრივი სტრუქტურა. 1 - არეალის საზღვრები; 2 - კუნძულებრივი არეალი XXს. 30-იან წლებში; 3 - არეალი XXს; 70-იან წლებში, დაცვის ღონისძიების გატარების შემდეგ (ა. ნასიმოვიჩი, ტიმოფეევი, 1973).
სივრცობრივი იზოლაცია შეიძლება იყოს სრული ან არასრული (ფარდობითი). მაგ., საქართველოში გავრცელებული პრომეთესეული მემინდვრია (rometheomys schaposchnicovi) სრულად იზოლირებული ოთხი პოპულაციითაა წარმოდგენილი. ორი პოპულაცია დიდ კავკასიონზე გვხვდება (მის დასავლეთ ნაწილსა და არაგვის სათავეებში), ორიც - მცირე კავკასიონზე (მესხეთისა და არსიანის ქედებზე). დიდ და მცირე კავკასიონზე გავრცელებული არჩვის (Rupicapra rupicapra) პოპულაციები არასრულადაა გამიჯნული, მათ ლიხის ქედი აკავშირებს. სივრცობრივი გამიჯვნა არ იწვევს რეპროდუქციულ იზოლაციას. თუ ინდივიდებმა ზღუდე გადალახეს, მაშინ ისინი ერთიმეორეს თავისუფლად შეეჯვარებიან. იგი ფიზიკურად მიჯნავს პოპულაციებს და აბრკოლებს გენთა ნაკადს. ამრიგად სივრცობრივი იზოლაცია ფიზიკური ხასიათისაა და არსებითად განსხვავდება ბიოლოგიური იზოლაციისაგან.
ბიოლოგიური იზოლაცია ორი ტიპის პრეკოპულაციური და პოსტკოპულაციური მექანიზმით ხორციელდება. ამის გამო შეუძლებელი ხდება თავისუფალი შეჯვარება (პანმიქსია) და წყდება გენთა ნაკადი. პრეკოპულაციურ მექანიზმებში ერთიანდება იზოლაციის ყველა ის ფორმა, რომელიც ინდივიდთა შეჯვარებას აბრკოლებს. მათ მიეკუთვნება ეკოლოგიური, მორფო-ფიზიოლოგიური და ეთოლოგიური იზოლაციის ფორმები. პოსტკოპულაციური იზოლაციის მექანიზმები შეჯვარებულ ინდივიდებში გენომთა შეუთავსებლობას იწვევს, მას მიეკუთვნება საკუთრივ გენეტიკური იზოლაცია. ზოგჯერ მორფო-ფიზიოლოგიურ, ეთოლოგიურ და საკუთრივ გენეტიკურ იზოლაციას ერთ ჯგუფში - რეპროდუქციულ იზოლაციაში - აერთიანებენ.
იზოლაციის ეკოლოგიურ ფორმას სახეობის ის საარსებო პირობები განსაზღვრავს, რომელიც გამრავლების ადგილსა და ვადებთანაა დაკავშირებული. ამიტომ გამოყოფენ სეზონურ და ბიოტოპურ იზოლაციას. მონათესავე ინდივიდების შეჯვარებას აბრკოლებს სქესობრივი აქტივობისა და გამეტების მომწიფების ვადების სხვადასხვაობა. გამრავლების სეზონური გამიჯვნა მკვეთრ და სწრაფ დაცილებას იწვევს.
მცენარეებში დიდი მნიშვნელობა აქვს განვითარებისა და ვეგეტაციის პერიოდების დაცილებას (ასინქრონულობას). მათში გამოვლენილია ისეთი სახეობები, რომლებშიც გაერთიანებული ინდივიდები ყვავილობისა და მტვრის მარცვლების გავრცელების პერიოდით განსხვავდებიან. ნ.ცინგერმა ხრიალაზე (Alectorolophus major) დაკვირვებისას (1920) შენიშნა მემკვიდრეობით განსაზღვრული საადრეო და საგვიანო ფორმები. ადამიანის მიერ თაობათა მანძილზე წარმოებულმა შეუგნებელმა გადარჩევამ (გათიბვამ) ხრიალაში ფენოლოგიურად განსხვავებული ფორმების ჩამოყალიბება გამოიწვია (იხ.: სურ. 7.6). სეზონური იზოლაცია გამოვლენილია ცხოველებშიც. სეზონური (ე.წ. „საგაზაფხულო“ და „საშემოდგომო“) რასები აღწერილია სალამურაში (ampetra), თევზებიდან ძირითადად გვხვდება ორაგულისებრთა, ზუთხისებრთა და კობრისებრთა რიგში გაერთიანებულ სახეობებში. ტოფობის ვადებით მკვეთრად განსხვავდებიან ერთსა და იმავე ტბაში ბინადარი თევზები. მაგ., ორხიდისა (მაკედონია) და სევანის (სომხეთი) ტბაში გავრცელებული კალმახის ენდემური სახეობები განსხვავდებიან ტოფობის პერიოდით, საქვირითე ადგილითა და სიღრმით. იზოლაციის მსგავსი ფორმები გვხვდება მწერებშიც. მაგ., მუხის აბრეშუმხვევიას (asiocanpa quercus) პეპლები ჩნდებიან ივლის-აგვისტოში, მისი მორფოლოგიურად მსგავსი ქვესახეობა (.q. collunae) მაის-ივნისში. ამდენად, მათი შეჯვარება გამორიცხულია.
ბუნებაში გაცილებით უფრო ხშირად გვხვდება ბიოტოპური იზოლაცია. პოტენციური პარტნიორები გამრავლების პერიოდში ერთიმეორეს ვერ ხვდებიან. მაგალითად, გომბეშოს ერთი ფორმა (ufo americanus) მრავლდება ჭაობებსა და ტბებში, მეორე (fewleri) გუბურებსა და ნაკადულებში. ბიოტოპური იზოლაციის საინტერესო მაგალითია სიმპატრიული შიდასახეობრივი ფორმების არსებობა ჩვეულებრივ გუგულში (Cuculus conorus). ევროპაში გუგულის რამდენიმე განსხვავებული ფორმა ბინადრობს, რომლებიც მხოლოდ გენეტიკურად დეტერმინირებული კვერცხის შეფერილობით განსხვავდებიან. კერძოდ, აღმოსავლეთ ევროპაში გავრცელებული გუგულის ერთი ფორმა დებს ცისფერ კვერცხს, რომელსაც ბოლოცეცხლას ან მდელოს ოვსადის ბუდეში ათავსებს, მეორე კი თეთრ დაწინწკლულ კვერცხს დებს, რომელსაც პატარა ბეღურისნაირ ფრინველთა ბუდეში ათავსებს. გუგულის განსხვავებულ ფორმებს შორის იზოლაციის შენარჩუნება მასპინძელი ფრინველის მიერ ნაკლებშენიღბული კვერცხების განადგურების შედეგად ხორციელდება.
იზოლაციის მორფო-ფიზიოლოგიური ფორმა გამრავლების ორგანოების განსხვავებული აგებულებითა და ფუნქციითაა გამოწვეული. იზოლაციის ამ ფორმის დროს მკვეთრად ქვეითდება განაყოფიერების ალბათობა, ე.ი. რეპროდუქტულ პერიოდში სასქესო პროდუქტების (გამეტების) შეხვედრის ალბათობა მეტად დაბალია. მცენარეებში ასეთ ბარიერს ქმნის ჰეტეროსტილია (ყვავილში მტვრიანების და ბუტკოს სხვადასხვა დონეზე განლაგება). იგი აბრკოლებს თვითდამტვერვას. მაგ., ჩვეულებრივ ფურისულაში (rimula vulgaris) პოპულაციები გრძელ და მოკლებუტკოიანი ყვავილების თანაფარდობით განსხვავდება (§9.1 და 10.1). ზოგიერთი ენტემოფილური ყვავილი რთული აგებულებისაა, რაც ყვავილის დამტვერავ მწერებთან შეგუებითაა გამოწვეული. მცენარეებში იზოლაციის ამავე ფორმას მიეკუთვნება გენეტიკურად დეტერმინირებული მტვრის მარცვლის განსხვავებული გაღივების უნარი და სისწრაფე. თუ დინგზე სხვა სახეობის მცენარის მტვერი მოხვდა, იგი ნელა ან ვერ ღივდება. სამტვერე მილი ნელა ყალიბდება, რომელიც ხშირად წყვეტს განვითარებას. ყოველივე ზემოთ აღნიშნული აბრკოლებს გამეტების შეხვედრას.

სურ. 7.6. ხრიალას (lectorolophus major) ქვესახეობების ჩამოყალიბების სქემა. საწყისი ფორმა .m. montanus (1) გავრცელებულია ყამირ ალპურ მდელოებზე. სათიბებზე გვხვდება ადრე მოყვავილე m. aestinalis (2). ორჯერადი გათიბვის შედეგად მისგან ჩამოყალიბდა ორი ქვესახეობა: m. vernalys (3) მწიფდება ადრე ზაფხულში და m. polycladus (4). მწიფდება გვიან ზაფხულში. ჭვავის ყანაში გავრცელებული სარეველა (5), ქვესახეობა m. apterus (ნ. ცინგერი, ი. ჭუჭულაშვილით, 1992).
ცხოველებში მონათესავე სახეობებს ზოგიერთ შემთხვევაში განსხვავებული აგებულების საკოპულაციო ორგანოები აქვთ, რის გამოც შეუძლებელი ხდება განაყოფიერება. იგი აღწერილია ზოგიერთ ფილტვიან მოლუსკებში, მწერებსა და მღრღნელებში. მაგ., სასქესო ორგანოების განსხვავებული აგებულების გამო ვერ შეჯვარდებიან ერთსა და იმავე ბიოტოპში გავრცელებული მოლუსკები ჩეპეაე ნამორალის და ჩ.ჰორტენსის. ზოგჯერ ახლომდგომ სახეობებში კოპულაცია ნორმალურად მიმდინარეობს. მაგრამ იმუნური შეუთავსებლობის გამო მდედრის სასქესო გზებში სპერმატოზოიდები იღუპებიან. ეს შემთხვევა აღწერილია rosophila-ს გვარში გაერთიანებულ სახეობებში.
იზოლაციის ეთოლოგიური ფორმა ქცევის თავისებურებათა განსხვავების მეშვეობით უზრუნველყოფს მონათესავე ფორმების გამიჯვნას. ბუნებრივია, იზოლაციის ეს სახე გვხვდება მხოლოდ იმ ცხოველებში, რომელთაც კარგად აქვს განვითარებული ნერვული სისტემა (მაგ., მწერები, თევზები, მყვარები ანუ ამფიბიები, რეპტილიები, ძუძუმწოვრები). მრავალ მათგანს აქვს სპეციფიკური ნიშნები (მეორადი სასქესო ნიშნები), რითაც მონათესავე ფორმები ადვილად პოულობენ ერთიმეორეს, ასევე ასხვავებენ სხვა სახეობის ინდივიდებს. ახლომდგომი ცხოველები სპეციფიკური სიგნალებით ადვილად ცნობენ და უკავშირდებიან ერთიმეორეს. ასეთი სიგნალები სხვადასხვა გამღიზიანებლებია (მაგ., მხედველობითი, სმენითი, ყნოსვითი და სხვ.). ეთოლოგიური იზოლაციის ნაირგვარი მექანიზმებია გამოვლენილი, მაგ., საქორწინო რიტუალში, ასევე მხედველობით, სმენით გაღიზიანებაში უმნიშვნელო ცვლილება ინდივიდების შეხვედრის ალბათობას მკვეთრად აქვეითებს. ცხოველები გამოყოფენ სპეციფიკურ ნივთიერებებს - ფერომონებს, რომელიც შორ მანძილზე ვრცელდება. გენეტიკურად ახლომდგომი ინდივიდები ადვილად პოულობენ ერთიმეორეს და შეჯვარდებიან. საკმარისია შეიცვალოს ფერომონის ქიმიური შედგენილობა, რომ ინდივიდებს შორის ჩნდება ბარიერი. მაგ., გარეული იხვი (Anas crecცო) და კუდსადგისა იხვი (A.acuta) ერთსა და იმავე ბიოტოპში ბინადრობენ, მაგრამ ბუნებაში ძალიან იშვიათად შეჯვარდებიან, ვოლიერებში კი შეჯვარდებიან და მათი შთამომავლობა ფერტილურია.
ჩრდილოეთ ამერიკაში გავრცელებული ციცინათელების გვარში (Photurus) გაერთიანებულია გენეტიკურად ახლომდგომი სახეობები და ზოგჯერ ერთი სახეობის პოპულაციებიც კი ნათების ხანგრძლივობით, სიხშირით და ინტენსიურობით განსხვავდებიან (იხ.: სურ. 7.7.).
საიზოლაციო მექანიზმების მეორე ჯგუფი (ე.წ. პოსტკოპულაციური) შეჯვარების შემდეგ მოქმედებენ და იწვევენ გამიჯვნას. მას საკუთრივ გენეტიკური იზოლაცია მიეკუთვნება. აქვე უნდა აღინიშნოს, რომ ზემოთ განხილული პრეკოპულაციური მექანიზმები გენეტიკური ფაქტორების მოქმედებითაა გამოწვეული. მაგრამ საჭიროა გამოიყოს საკუთრივ გენეტიკური ფაქტორები. მათ მიეკუთვნება: 1. პოლიპლოიდია* 2. ქრომოსომული აბერაციები* 3. ბირთვულ-ციტოპლაზმური შეუთავსებლობა* 4. მუტაციის შედეგად გამოწვეული ცალკეული გენის ექსპრესიის შეუთავსებლობა.

სურ. 7.7. ეკოლოგიურ-ეთოლოგიური იზოლაცია ციცინათელებში (გვარი Photurus) შავი ლაქით მოცემულია ნათების ხანგრძლიობა და ინტენსიობა (ე. მაირი, 1968).
გენეტიკური საიზოლაციო მექანიზმების მოქმედებით იღუპება ზიგოტები ან ონტოგენეზის ადრეულ ეტაპზე ჩანასახები. მართალია, ზოგჯერ ახლომდგომი სახეობების შეჯვარებით წარმოქმნილი ჰიბრიდები სიცოცხლისუნარიანნი არიან, მაგრამ მათში ნაყოფიერება მკვეთრადაა დაქვეითებული, მეტწილად სტერილურები არიან. ისინი საწყის ფორმებს კონკურენციას ვერ უწევენ და ელიმინირდებიან. ამრიგად, როგორც გენეტიკური, ისე ბიოლოგიური იზოლაციის სხვა ფორმები ამაღლებენ მონათესავე ინდივიდების შეჯვარების ალბათობას, რის გამოც მკვეთრად ადიდებენ ინბრიდინგს პოპულაციაში.
იზოლაცია შემთხვევით, არამიმართულ და განუზღვრელი მოქმედების ფაქტორს წარმოადგენს. იგი, როგორც ევოლუციის ფაქტორი, არ წარმოქმნის ახალ გენოტიპებსა და შიდასახეობრივ ფორმებს. ევოლუციის პროცესში იზოლაციის მნიშვნელობა იმაში მდგომარეობს, რომ იგი გენეტიკური დიფერენცირების საწყის სტადიებს განამტკიცებს, აღრმავებს და აძლიერებს. აგრეთვე, იზოლირებულ პოპულაციებსა ან პოპულაციის შემადგენელ ნაწილზე გარდაუვალია ბუნებრივი გადარჩევის განსხვავებული ზეწოლა. იზოლაცია დივერგირებულ ფორმებში ჩამოყალიბებულ სპეციფიკური გენოფონდის შენარჩუნებას აპირობებს. ხანგრძლივი მოქმედება-იზოლაციის, როგორც ევოლუციის ელემენტარული ფაქტორის, მთავარი დამახასიათებელი ნიშანია. იზოლაციის მეშვეობით საწყისი ერთიანი პოპულაცია ორ ან მეტ შემადგენელ ნაწილად იყოფა. ინდივიდთა თითოეული ჯგუფი ხანგრძლივი დროით გამიჯნული რჩება. ამრიგად, ევოლუციის პროცესის ნებისმიერი ხანგრძლივი ეტაპი იზოლაციის მოქმედებით მიმდინარეობს.
ბიოლოგიური იზოლაციის ყველა მექანიზმი ევოლუციის შედეგია და იგი ჩამოყალიბდა როგორც ადაპტური თვისება, რომელიც სახეობის მდგრადობას უზრუნველყოფს. ამიტომაც იზოლაციის ეკოლოგიურ, მორფოფიზიოლოგიურ და გენეტიკურ ფორმას განიხილავენ როგორც სახეობის ძირითად კრიტერიუმს.
|
|
10 §8. ბუნებრივი გადარჩევა - ევოლუციის მთავარი და წარმმართველი ფაქტორი |
▲ზევით დაბრუნება |
როგორც აღვნიშნეთ, ევოლუციის ელემენტარული ფაქტორები: გენთა ნაკადი, მუტაციური პროცესი, გენთა დრეიფი და იზოლაცია არამიმართული, სტოქასტური და განუსაზღვრელი მოქმედებისაა. ევოლუცია ქაოსური როდია, არამედ შეუქცევადი და მიმართული პროცესია, რომლისთვისაც ნიშანდობლივია ახალი ადაპტაციების ფორმირება, ახალ სახეობათა წარმოქმნა და ზოგიერთის ამოწყდომა, ტაქსონების იერარქიული სისტემის წარმოქმნა, ორგანული ბუნების პროგრესული განვითარება. ცნობილია მხოლოდ ერთადერთი მიმართული ფაქტორი - ბუნებრივი გადარჩევა, რომელიც აწესრიგებს და წარმართავს ევოლუციურ პროცესს.
ბუნებრივი გადარჩევის ხელშემწყობი პირობაა: 1. პოპულაციის გენეტიკური ჰეტეროგენულობა* 2. გამრავლების პროგრესია* 3. ბრძოლა არსებობისათვის.
|
|
10.1 ბუნებრივი გადარჩევის წინამძღვრები |
▲ზევით დაბრუნება |
|
|
10.1.1 8.1. პოპულაციის ჰეტეროგენულობა |
▲ზევით დაბრუნება |
გენეტიკური ჰეტეროგენულობა პოპულაციის ერთ-ერთი მნიშვნელოვანი თავისებურებაა. პოპულაციათა გენეტიკური შესწავლით გაირკვა მნიშვნელოვანი მოვლენა, კერძოდ, ის რომ ბუნერივი პოპულაციები ფარული მემკვიდრული ცვლილებებით (მუტაციებით) ყოფილა გაჯერებული. პოპულაციის ყოველი ინდივიდის გენოტიპი ჰეტეროზიგოტულ მდგომარეობაში შეიცავს, როგორც ახლად ინდუცირებულ, ისე ადრე წარმოქმნილ და მემკვიდრეობით გადმოცემულ მუტაციებს. ჰეტეროგენულობა (იხ. §9.3) მუტაციური პროცესის განუწყვეტელი მოქმედებით და რეკომბინაციით შენარჩუნდება. სხვადასხვა მეთოდებით დადგენილია ცილებისა და მათი განმსაზღვრელი ლოკუსების მაღალი პოლიმორფულობა. გამოირკვა, რომ მრავალი ლოკუსი პოპულაციაში ორზე მეტი ალელითაა (ე.წ. პოლიალელური ლოკუსები) წარმოდგენილი. მცენარეებისა და ცხოველების მრავალრიცხოვან სახეობებში ჩატარებული გამოკვლევებით მათში განისაზღვრა საშუალო ჰეტეროგენულობის დონე. გამოირკვა, რომ ინდივიდში ლოკუსთა საერთო რაოდენობიდან 15-53%25 პოლიალელურია, ხოლო 4-15%25 ჰეტეროზიგოტური. დადგენილია, რომ ასევე მაღალი ჰეტეროგენულობა ახასიათებთ პართენოგენეზულ პოპულაციებს (მაგ., ზოგიერთი პეპელა, ხოჭო, ტარაკანი და სხვ.) და თითქმის არ განსხვავდებიან ტიპური სქესობრივი გამრავლების უნარიანი (პანმიქტური) პოპულაციებისაგან. ჰეტეროგენულობით უზრუნველყოფილია მემკვიდრული ცვალებადობის „სამობილიზაციო რეზერვის არსებობა“. იგი ბუნებრივი გადარჩევის მოქმედების ერთ-ერთი მნიშვნელოვანი ხელშემწყობი პირობათაგანია.
|
|
10.1.2 8.2. გამრავლების პროგრესია |
▲ზევით დაბრუნება |
ბუნებრივი გადარჩევის მეორე აუცილებელ წინაპირობას გამრავლების პროგრესია წარმოადგენს. ბუნებრივი გადარჩევა მხოლოდ იმ შემთხვევაშია შესაძლებელი, როდესაც მშობლების მიერ პროდუცირებული ინდივიდები მათი შეცვლისათვის საჭირო რაოდენობას აღემატება. თუ პოპულაციის თითოეული წყვილი მხოლოდ ორ ზრდასრულ ინდივიდს დატოვებს, მაშინ ბუნებრივი გადარჩევის გზით ევოლუცია ვერ განხორციელდება.
ცხრილი 8.1
სეზონის განმავლობაში ერთი ინდივიდის
რეპროდუქციის რიცხოვნობა
სახეობა |
ლათინური |
შთამომავალთა |
ადამიანი |
Homo sapiens |
1 |
ძაღლი |
Canis familaris |
7 (1 - 22) |
მგელი |
Canis lupus |
3 - 10 |
შოშია |
Sturnus |
5 - 8 |
ჩვეულებრივი ანკარა |
artix natrix |
15 - 30 |
გიურზა |
Vipera lebetina |
15 - 20 |
სამეფო კობრა |
Phiphagus |
20 - 40 |
ჩვეულებრივი გომბეშო |
ufo bufo |
1200 - 7000 |
ტბის ბაყაყი |
Rana |
5000-10000 |
ბაყაყი - ხარი |
Rana |
6000-20000 |
ვირთევზა |
Cadus |
2,5 - 10 |
კალმახი |
Salmo truta |
200 - 6000 ქვირითი |
ადამიანის |
Scaris |
200000 |
ოთახის ბუზი |
Usca |
600 - 2000 |
ხილის ბუზი |
Rosophila |
100 კვერცხი |
ლენცოფა |
Yascyanus |
446500 |
წიწმატურა |
Capseila |
73000 |
მინდვრის |
Cirsium |
35000 |
ბაბუაწვერა |
Taraxacum |
100000 |
ხაშხაში |
Apaver |
30000 |
ორგანულ სამყაროში ხდება გამრავლების პროგრესია. ყოველი ორგანიზმი შთამომავლობის მოცემის დიდი უნარით გამოირჩევა (იხ. ცხრილი 5-3). იგი ყოველ სახეობაში ინდივიდთა ჭარბ რაოდენობას წარმოქმნის. ინდივიდები გაცილებით დიდი რაოდენობით ჩნდებიან, ხოლო ზრდასრულ ფორმას უმნიშვნელო რაოდენობა თუ აღწევს. ინდივიდების გეომეტრიული პროგრესიით გამრავლების უნარიანობას მრავალმა მეცნიერმა (უ. სენტ-ილერი, კ. ლინე, თ. მალთუსი) მიაქცია ყურადღება. მათ გამოავლინეს საგულისხმო ფაქტი, რომ ყოველ სახეობას ჰქონია უნარი გამრავლების პოტენციური შესაძლებლობის რეალიზაციის შემთხვევაში უმოკლეს დროში განსახლდეს და დედამიწის ზედაპირი დაფაროს. ჩ.დარვინამდე ვერავინ შეძლო ამ მოვლენის არსს ჩასწვდომოდა. მან განსაზღვრა გამრავლების პროგრესიის მნიშვნელობა ევოლუციის პროცესში, რაც არსებობისათვის ბრძოლის წინაპირობაა.
ცხრილი 5-3-დან ჩანს, რომ ინდივიდები გამრავლების დიდი უნარით გამოირჩევიან. ერთი ქაშაყი ყოველწლიურად 40 ათასამდე ქვირითს ყრის, ვირთევზა 10 მილიონამდე, ხოლო ტბის ბაყაყი 10 ათასამდე. წყვილ ბეღურას 10 წლის მანძილზე 200 მილიარდი ინდივიდის მოცემა შეუძლია. გამრავლების პროგრესიის დასახასიათებლად პირობით (ჰიპოთეზურ) მაჩვენებელს-ბიოტურ პოტენციალს იყენებენ, რითაც გამოხატავენ თუ ინდივიდს შეუზღუდველი გამრავლების შემთხვევაში დროის რა მონაკვეთში შეუძლია თანაბარი შრით დაფაროს დედამიწა. ბაბუაწვერას ერთი მცენარე მეორე წელს 100 ინდივიდს წარმოქმნის. 5 წლის განმავლობაში 107-ს, 10 წლის განმავლობაში კი-1017-ს. 10 წლის განმავლობაში მას შეუძლია დედამიწის ზედაპირი 20 სმ-ის სისქის ფენით დაფაროს. ხაშხაშს კი ჩვენი პლანეტის ზედაპირის დაფარვა 3-4 წლის განმავლობაში შეუძლია. გაცილებით მაღალი გამრავლების უნარი ახასიათებს მიკროორგანიზმებს. მაგალითად, ფიშერის ბაქტერიას 1,5 დღე-ღამის განმავლობაში შეუძლია დედამიწის ზედაპირის ფართობი თხელი შრით დაფაროს. გამრავლების პროგრესია ორ მნიშვნელოვან შედეგს იწვევს: 1. იქმნება ერთგვარი „სიცოცხლის ზეწოლა“, რაც არსებობისათვის ბრძოლის საფუძველია* 2. მიიღება მემკვიდრული ცვალებადობის ფართო სპექტრი, რომელიც მასალაა ბუნებრივი გადარჩევისათვის.
|
|
10.1.3 8.3 ბრძოლა არსებობისათვის |
▲ზევით დაბრუნება |
ორგანიზმის აქტივობას, მიმართულს სიცოცხლის შესანარჩუნებლად და შთამომავლობის არსებობის უზრუნველსაყოფად, არსებობისათვის ბრძოლა ეწოდება. არსებობისათვის ბრძოლას საფუძვლად უდევს ორგანიზმთა ტენდენცია განუსაზღვრელი გამრავლებისაკენ და გარემოს ბიოტური და აბიოტური ფაქტორების წინააღმდეგობა ამ გამრავლებისადმი. როგორც უკვე აღინიშნა, ორგანიზმები გამრავლების დიდი უნარით გამოირჩევიან, მაგრამ მათი არსებობისათვის აუცილებელი პირობები (ტერიტორია, საკვები, წყალი და მისთ.) შეზღუდულია. ამდენად იქმნება შეუსაბამობა ინდივიდთა რაოდენობასა და საარსებო პირობებს შორის, რომელიც არსებობისათვის ბრძოლას უზრუნველყოფს. ბუნებაში გამრავლების პოტენციური შესაძლებლობის სრული რეალიზაცია შეუძლებელია. ყველა გაჩენილ ინდივიდს ზრდა სრული ასკისათვის რომ მიეღწია, მაშინ მათთვის საარსებო პირობები აღარ იქნებოდა. ამრიგად, ორგანიზმებს შორის ყოველთვის მიმდინარეობს ბიოლოგიური შეჯიბრი ანუ კონკურენცია საარსებო პირობების მოსაპოვებლად. მეორე მხრივ, ორგანიზმები წინააღმდეგობას უწევენ გარემოს ბიოტური (მტრები, პარაზიტები და სხვ.) და აბიოტური (ტემპერატურის ცვალებადობა, ჟანგბადის, წყლის, სინათლის ნაკლებობა ან სიჭარბე და მისთ.) ფაქტორების მავნე ზემოქმედებას. ორგანიზმებისა და გარემო ფაქტორების ყველა ურთიერთმიმართება ჩ. დარვინმა გააერთიანა კრებსითი ცნებით - ბრძოლა არსებობისათვის. როგორც კლასიკურ დარვინიზმში, ისე ევოლუციის სინთეზურ თეორიაში ცნება - არსებობისათვის ბრძოლა მეტაფორული გაგებით იხმარება.
ამრიგად, ცნებაში „ბრძოლა არსებობისათვის“ იგულისხმება ორგანიზმის აქტივობა, რომლითაც მიიღწევა როგორც თავისი, ისე მისი შთამომავლობის სიცოცხლის უზრუნველყოფა. ეს ცნება არ უნდა გავიგოთ ვულგარულად. როგორც ინდივიდთა უბრალო ფიზიკური დაპირისპირება. იმის კითხვა თუ რა ფაქტორების მიმართ უხდებათ ბრძოლა? ინდივიდებს ბრძოლა უხდებათ ყველა იმ ფაქტორის მიმართ რომლებიც ზღუდავენ მათ რიცხოვნობას. ასეთი ფაქტორები მეტად მრავალრიცხოვანია. ინდივიდთა აქტივობა არსეობისათვის ბრძოლის ყველა ფორმაში მჟღავნდება და ცოცხალი სისტემის ერთ-ერთი მნიშვნელოვანი თვისებაა.
არსებობისათვის ბრძოლაში ორგანიზმს აქტივობის განსაზღვრისათვის ჩვეულებრივ, მთავარ როლს გამრავლების გეომეტრიულ პროგრესიას მიაწერენ. ამ მახასიათებლით მხოლოდ გამრავლების ინტენსივობის პროცენტული დახასიათებაა შესაძლებელი. გამრავლების გეომეტრიული პროგრესიით შეუძლებელია ვიმსჯელოთ კონკრეტულ გარემოში ორგანიზმზე მოქმედ „სიცოცხლის ზეწოლაზე“, ასევე თუ რამდენად ეკონომიურად იყენებს ორგანიზმი საარსებო რესურსებს. ასე მაგალითად, ქარიშხალა წელიწადში მხოლოდ ერთ კვერცხს დებს, მაგრამ მისი აქტივობა სიცოცხლის შესანარჩუნებლად გაცილებით მაღალია, ვიდრე სოკოსი (მილიარდობით სპორას წარმოქმნის) ან მტკნარი წყლის მოლუსკის - მარგალიტანასი (3 მილიონამდე კვერცხს ყრის). ეს ორი უკანასკნელი უმნიშვნელო აქტივობას ავლენს.
ორგანიზმის რეალური გამრავლების უნარი, მისი ნაყოფიერებით განისაზღვრება ე.ი. მის მიერ მთელი სიცოცხლის განმავლობაში წარმოქმნილი შთამომავლობის რაოდენობით. სქესობრივი გამრავლებისას, ყვავილოვან მცენარეში ეს არის მომწიფებული თესლების საერთო რაოდენობა, ფრინველებში - დადებული კვერცხების. ბუნებრივია, ეს მაჩვენებელი კონსტანტური არ არის. იგი სახეობაში გაერთიანებულ ინდივიდებში საარსებო გარემოს მიხედვით და ეკოლოგიური ფაქტორების (კლიმატური ფაქტორების ცვალებადობა, ბიოლოგიური მტრებისა და საკვები რესურსების რაოდენობის ცვლილება, ადამიანის ზემოქმედება და მისთ.) ზემოქმედებით წლიდან წლამდე მერყეობს. ინდივიდის ნაყოფიერება სახეობისათვის ნიშანდობლივ საშუალო მაჩვენებლის მახლობელ (რეაქციის ნორმის) ფარგლებში ვარირებს.
როდესაც კონკრეტული სახეობის ინდივიდთა საშუალო რიცხოვნობა გარკვეულ პერიოდში მუდმივია, ეს იმას ნიშნავს, რომ ყოველწლიურად ჩნდება იმდენივე ინდივიდი, რამდენიც სხვადასხვა მიზეზით იღუპება. რეპროდუქციის უნარიან ინდივიდთა რაოდენობა არ იზრდება. კონკრეტული სახეობის მიერ გამოვლენილი „სიცოცხლის ზეწოლა“ მუდმივი რჩება. თუ N რაოდენობის სქესმწიფე ინდივიდები, ყოველწლიურად ნ რაოდენობის ნაშიერს ან კვერცხს წარმოქმნის, ხოლო განსხვავებული ასაკის (ძირითადად სქესობრივად მოუმწიფებელი) ნ რაოდენობის ინდივიდი იღუპება, მაშინ კონკრეტულ პოპულაციაში (ასევე სახეობაში) ინდივიდთა საერთო რაოდენობა (N+N-N=N) უცვლელი რჩება.
ამდენად, გამრავლების გეომეტრიული პროგრესია, არ ასახავს „სიცოცხლის ზეწოლას“, ვინაიდან მკვდრადშობილი (მცენარეებში მოუმწიფებელ თესლები), სტერილური ან გენერაციულ სტადიამდე დაღუპული ინდივიდები შთამომავლობას არ ტოვებენ. რეალური „სიცოცხლის ზეწოლა“ განისაზღვრება ფაქტობრივად არსებული ინდივიდებით და არა ინდივიდთა იმ რიცხვით რომლებიც შეიძლება გაჩენილიყვნენ ან შთამომავლობა მოეცათ ხელსაყრელ საარსებო პირობებში. გაჩენილ ინდივიდთა საშუალო რაოდენობა თუ ტოლია დაღუპულ ინდივიდთა რაოდენობის, მაშინ არსებობისათვის ბრძოლის ინტენსიობის გარკვეულ მაჩვენებელს ვღებულობთ.
ორგანიზმის ნაყოფიერება (მაგალითად, მთელი სიცოცხლის მანძილზე დადებული კვერცხების ან განვითარებული თესლების რაოდენობა) დროის გარკვეულ მონაკვეთში არსებობისათვის ბრძოლის საერთო ინტენსიობის საზომ სიდიდეს წარმოადგენს, ე.ი. ყველა ასაკში (ჩასახვიდან სქესმწიფე ასაკის ჩათვლით) ორგანიზმის ჯამური ელიმინაციის სიდიდეს გამოხატავს.
|
|
10.1.3.1 8.3.1. არსებობისათვის ბრძოლის ფორმები |
▲ზევით დაბრუნება |
ნებისმიერი სახეობის პოპულაცია დამოუკიდებლად და განმხოლოებულად როდი არსებობს, იგი ბიოცენოზის შემადგენელი კომპონენტია. მისი წარმატებული არსებობა დიდად არის დამოკიდებული იმაზე, თუ როგორ ურთიერთობას დაამყარებენ კონკრეტული პოპულაციის წევრები ერთმანეთთან ან სხვა სახეობის პოპულაციებთან. ამ რთული ურთიერთდამოკიდებულების ფორმირება აბიოტური ფაქტორების ფონზე მიმდინარეობს. ამდენად ორგანულ სამყაროში არსებობისათვის ბრძოლის სახეები, როგორც გამოვლენის ისე თავისი მოქმედების შედეგებით მეტად მრავალფეროვანია. არსებობისათვის ბრძოლის სხვადასხვა ფორმა დაკავშირებულია ბუნებრივი გადარჩევის სხვადასხვა მიმართულებასთან. ამდენად, მისი არსის გაგებისათვის აუცილებელია ამ ფორმების დაჯგუფება. ევოლუციური თეორიის სახელმძღვანელოში ზოგიერთი კომენტატორი ნაკლებ ყურადღებას უთმობს ამ საკითხს და მის როლს ევოლუციის პროცესში. ზოგჯერ არსეობისათვის ბრძოლას აიგივებენ უშუალო შებრძოლებასთან (ჩხუბი, აგრესია, განადგურება და მისთ.) ან მტაცებელი - მსხვერპლის დამოკიდებულებამდე დაჰყავთ.
დარვინისეული გაგებით არსებობისათვის ბრძოლაში იგულისხმება, როგორც გარემო (აბიოტური და ბიოტური) ფაქტორების მოქმედება ორგანიზმზე, ისე ორგანიზმის აქტიობა - მათი გავლენა გარემოზე. დარვინი გამოყოფდა არსებობისათვის ბრძოლის ორ ძირითად ფორმას: შიგასახეობრივსა და სახეობათა შორისს. ლ. მორგანმა (1906) და ლ. პლატემ (1912) დამატებით გამოყვეს მესამე ფორმა: კონსტიტუციური ბრძოლა არსებობისათვის, რომელიც ევოლუციურ თეორიაში საკმაოდ არის დამკვიდრებული. უნდა აღინიშნოს, რომ ჩ. დარვინი თავის ნაშრომში „სახეობათა წარმოშობა“ ვრცლად მსჯელობს ბრძოლის ამ ფორმაზე, მაგრამ ფორმულირება არ მოუხდენია.
1. კონსტიტუციური ბრძოლა არაორგანული ბუნების ფაქტორებთან მიმდინარეობს იმ შემთხვევაში, როდესაც ინდივიდები ხვდებიან აბიოტური ფაქტორების (გვალვა, სიცივე, ჭარბი ან ნაკლები მარილიანობა, ჟანგბადი, განათება და მისთ.) მავნე ზემოქმედების პირობებში. იგი გამოიხატება ფიზიოლოგიური რეაქციებით. მაგ., ძუძუმწოვრებში ხშირი ბეწვის, ფრინველებში ღინღლის განვითარება, ზამთარში ან ზაფხულის გვალვებისას - ძილქუში, ინცისტირება, ანაბიოზში გადასვლა და მისთ. იგი ვლინდება აგრეთვე ქცევითი აქტებით, როგორიცაა სოროში ჩაძვრომა, ნიადაგში ჩამალვა, ადგილის შეცვლა, მიგრაცია და სხვ. ბრძოლის ამ ფორმის პროცესში იღუპებიან კონკრეტულ კლიმატურ პირობებთან ნაკლებ შეგუებული ფორმები, ხოლო გადარჩებიან და შთამომავლობას ტოვებენ უფრო ამტანი კონსტიტუციის მქონე ინდივიდები.
2. შიგასახეობრივი ბრძოლა არსებობისათვის მიმდინარეობს ერთი და იმავე სახეობის (პოპულაციის) ინდივიდებს შორის და განსაკუთრებული სიმძაფრით გამოირჩევა. ჩ. დარვინის შეხედულებით, ბრძოლის ეს ფორმა მეტად დაძაბულია, ძირითადად იგი განსაზღვრავს ინდივიდთა გადარჩენას ან ელიმინაციას, ამდენად ევოლუციისათვის მეტად მნიშვნელოვანია.
ეკოლოგიურად რაც უფრო ახლოს იმყოფებიან ერთმანეთთან კონკურენტი ინდივიდები, მით მეტი პარამეტრის (გარემო ფაქტორთა) მიხედვით მიმდინარეობს კონკურენცია. ცალკეული ორგანიზმის კონკურენტუნარიანობას თითოეული მათგანის ინდივიდური თავისებურებები განსაზღვრავს. ძირითადად იგი ვლინდება საკუთარი სიცოცხლის დაცვისა და შთამომავლობის უზრუნველყოფისათვის უფრო ეფექტურ საშუალებათა გამომუშავებაში, რაც უპირატესობას ანიჭებს მას სხვა ინდივიდებთან კონკურენციაში, მტრებისა და გარემოს არახელსაყრელ პირობებისადმი წინააღმდეგობის გაწევისას. ბრძოლის ამ ფორმას აშკარა ანუ პირდაპირი ხასიათი იშვიათად აქვს, რის გამოც იგი ნაკლებ თვალსაჩინოა.

სურ. 8.1. შიგასახეობრივი კონკურენცია წიწვოვან ტყეში (შმალგაუზენი, 1969).
სახეობის შიგნით (პოპულაცია, სახესხვაობა, ქვესახეობა და მისთ.) ურთიერთდამოკიდებულება უაღრესად მრავალფეროვანი, რთული და წინააღმდეგობრივია. მეტწილად ამ ურთიერთობის შედეგად ზოგიერთი ინდივიდის სიცოცხლისუნარიანობა ან ნაყოფიერება მკვეთრად ქვეითდება, რის გამოც მომდევნო თაობათა (გენოფონდის) ფორმირებაში მათი წვლილი მეტად უმნიშვნელო ან ნულის ტოლია. უმეტესად კონკურენტი ინდივიდები ერთიმეორეზე უშუალოდ არ ურთიერთქმედებენ. საარსებო რესურსების ექსპლუატაციის გამო კონკურენტებს უძნელდებათ სრულყოფილი საკვების და სხვა რესურსების (წყალი, სინათლე და მისთ.) მოპოვება. საილუსტრაციოდ შეიძლება მოვიყვანოთ ტოლასაკოვანი ფიჭვის ტყე. მასში გვხვდება კარგად განვითარებული (ფართოტოტებიანი, მძლავრად განვითარებული ფესვთა სისტემა) ინდივიდები. ისინი საარსებო რესურსების (მზის სხივური ენერგია, CO2, წყალი და მინერალური მარილები) მეტ ექსპლუატაციას ახდენენ ვიდრე მეზობლად მდგომი სუსტად განვითარებული ხეები. სრულყოფილად განვითარებული ფიჭვები მეტ გირჩებს (შესაბამისად თესლს) იკეთებენ, სუსტად განვითარებულნი საერთოდ არ იკეთებენ (ზოგი კი იღუპება) ან ძალიან მცირე რაოდენობით წარმოქმნიან გირჩებს. ბუნებრივია, მომდევნო გენოფონდის ფორმირებაში წარმატებულია იმ მცენარეთა ხვედრითი წილი, რომლებიც დიდი რაოდენობით იძლევიან თესლს (იხ. სურ. 8.1).
ზოგიერთ შემთხვევაში ბრძოლა შეიძლება განსხვავებული სახითაც გამოვლინდეს, კერძოდ ერთი ინდივიდი აბრკოლებს სხვებს, რომ მის მიერ მონიშნული ტერიტორია სხვა ინდივიდმა დაიკავოს. იგი ტერიტორიას იცავს სხვებისგან, რათა არ გამოიყენონ საარსებო რესურსები (სრულფასოვანი საკვები, სინათლე, წყალი და მისთ.) ამ სახის ურთიერთდამოკიდებულება (ე.წ. ინტერფერენცია) მკაფიოდ არის გამოხატული ხერხემლიან ცხოველებში, თუმცა შეინიშნება უხერხემლო ცხოველებსა და მცენარეებშიც. მაგალითად, მურა დათვს და ჯიქს საარსებო ტერიტორია ხეზე ნაკაწრებით აქვთ მონიშნული, მგელი შარდით, ხოლო წიწკანები და სკვინჩები გალობით აგებინებენ ერთიმეორეს ტერიტორიის დაკავებას. ცხოველები ერიდებიან მონიშნული ტერიტორიის საზღვრების დარღვევას.
პოპულაციის შემადგენელი ინდივიდებისათვის ნიშანდობლივია ურთიერთდახმარებაც: ერთობლივად ნადავლის მოპოვება და კვება, შთამომავლობის დაცვა და გამოზრდა. ეს თვისება მკაფიოდაა გამოხატული საზოგადოებრივ ცხოველებში (მაგალითად, ჭიანჭველების ბუდე, ფუტკრების სკა, ლომების პრაიდი, მაიმუნების ჯოგი და მისთ.). ურთიერთდახმარება შეინიშნება მცენარეებშიც. ხშირად ერთი და იმავე პოპულაციის ახლომდგომ ხემცენარეთა ფესვები ურთიერთშეზრდილია. იგი აძლიერებს ხეთა დგომას და ქარმედეგობას, აგრეთვე უზრუნველყოფს შეწოვილი წყლისა და მინერალური მარილების, ასევე სინთეზირებული ორგანული ნაერთების ურთიერთმიწოდებას. მაგალითად ხშირ ნაძვნარში ყოველი 100 მცენარიდან დაახლოებით 30-40-ს ფესვები შეზრდილი აქვს.
პოპულაციის შემადგენელ წევრებს შორის შეინიშნება ანტაგონიზმიც, მაგალითად, საფუარ სოკოებში, ქალამანაში ე.წ. „კილერი“ ინდივიდის მიერ გამოყოფილი ცილა ტოქსინით ახდენს მეორე მგრძნობარე ორგანიზმის ელიმინაციას (იხ. §10). ანტაგონიზმის უკიდურესი გამოვლენაა კანიბალიზმი (მაგალითად, ქორჭილა, კალმახი, ორაგული და სხვ.). იგი მართალია საზიანოა ცალკეული ინდივიდისათვის მაგრამ სასარგებლოა მთლიანად პოპულაციისათვის. ამ გზით რეგულირდება პოპულაციაში რიცხოვნობა და მიიღწევა საარსებო რესურსების რაციონალური გამოყენება.
პოპულაციის შემადგენელ ინდივიდებს შორის ურთიერთდამოკიდებულება სხვადასხვა პერიოდსა და გარემოსთან მიმართებით შეიძლება იყოს ნეიტრალური, კონკურენტული ან შესაძლებელია გამოვლინდეს ურთიერთდახმარების ფაქტები. მაგალითად, როდესაც ხელოვნურ საკვებ არეში დაბალია დროზოფილას ლარვების (მატლების) სიმჭიდროვე ისინი ერთმანეთს ეხმარებიან, რათა თავიდან აიცილონ საკვების გამოშრობა და მასზე ობის სოკოს განვითარება. საშუალო სიმჭიდროვის დროს ლარვებს შორის დამოკიდებულება ნეიტრალურია, მაღალი სიმჭიდროვის დროს კი მძაფრი კონკურენცია აღინიშნება.
3. სახეობათაშორისი ბრძოლა არსებობისათვის გაცილებით ნაირგვარი და რთულია ვიდრე ზემოთ განხილული ორი ფორმა. არსებობისათვის ბრძოლა სხვადასხვა სახეობის ინდივიდებს შორის ხშირად პირდაპირი ხასიათისაა. მაგ., ურთიერთდამოკიდებულება მცენარეებსა და ფიტოფაგ ცხოველებს შორის, მტაცებელსა და მსხვერპლს შორის, პარაზიტსა და მასპინძელს შორის, ასევე საკვების, ბუდის წართმევა და სხვ. ზოგჯერ ბრძოლის ეს ფორმა ფარულ (არაპირდაპირ) ხასიათს ატარებს. მაგალითად, ერთი სახეობის მიერ მეორის დათრგუნვა ან არეალიდან თანდათანობით გაძევება. სხვადასხვა სახეობის ინდივიდებს შორის ურთიერთდამოკიდებულება განსხვავებულია და სიმძაფრის ხარისხი დამოკიდებულია არსებობისათვის აუცილებელი პირობების მსგავსების ხარისხზე. ურთიერთდამოკიდებულება შეიძლება იყოს დადებითი (მაგ. სიმბიოზი) უარყოფითი (მაგ. მტაცებელი - მსხვერპლი) ან ნეიტრალური.
არსებობისათვის ბრძოლის სხვადასხვა ფორმას ევოლუციაში თუმცა დიდი მნიშვნელობა აქვს, მაგრამ ახალი ფორმების წარმოქმნა მხოლოდ კონკრეტული პოპულაციის შიგნით ინდივიდებს შორის კონკურენციის გზით მიმდინარეობს. დარვინისეული ბრძოლა არსებობისათვის ორ ძირითად მომენტს შეიცავს. ერთის მხრივ ის დაკავშირებულია გარკვეული სახეობის ინდივიდების დაღუპვასთან ე.ი. სხვადასხვა სახის ელიმინაციასთან. ეს თითქოს ბრძოლის უარყოფითი მახასიათებელია. მეორე მხრივ სახეობაში გაერთიანებულ ინდივიდებს შორის აქტიური ან პასიური კონკურენციის ელემენტები არსებობისათვის ბრძოლის მნიშვნელოვან დადებით დახასიათებას იძლევიან.
|
|
10.1.3.2 8.3.2 ელიმინაციის ფორმები |
▲ზევით დაბრუნება |
საარსებო პირობების შეზღუდვის ან არახელსაყრელი აბიოტური ფაქტორების მოქმედების გამო, არსებობისათვის ბრძოლის ყველა ფორმას თან სდევს ორგანიზმთა მეტ-ნაკლები ნაწილის ელიმინაცია (მოსპობა). ელიმინაციაში გამოიხატება გარემოს წინააღმდეგობა ორგანიზმთა გამრავლების მიმართ, ელიმინაციის კლასიფიკაცია შეიძლება მოვახდინოთ მოქმედების დროის მიხედვით, - მუდმივი და პერიოდული ელიმინაცია, მაელიმინირებელი ფაქტორების - ფიზიკური (სიცივე, გვალვა და სხვა), ფიზიოლოგიური (შიმშილი), ბიოლოგიური (მტაცებელი, პარაზიტი, ეპიზოოტია), ობიექტების (ინდივიდი, ოჯახი, პოპულაცია, სახეობა) ან ობიექტების ასაკის (გამეტა, ჩანასახი, ლარვა, ზრდასრული ინდივიდი) მიხედვით. რადგანაც ყველა ეს მახასიათებელი გარკვეული მნიშვნელობისაა და ყოველი მათგანი ევოლუციის მიმართულებაზე არსებობისათვის ბრძოლის სხვადასხვა ფორმის გავლენის სპეციფიკის გათვალისწინების საშუალებას იძლევა, ამიტომ განვიხილოთ შმალჰაუზენის კომბინირებული კლასიფიკაცია.
ზოგადი ანუ შემთხვევითი ელიმინაცია. ცალკეული ინდივიდის სიკვდილი დამოკიდებულია მხოლოდ შემთხვევაზე. სიცოცხლისა და სიკვდილის საკითხები ორგანიზმის ინდივიდუალური თვისებებით არ წყდება, რადგანაც იგი შემთხვევით აღმოჩნდება გამანადგურებელი ფაქტორის წინაშე, რომელიც ანადგურებს სახეობის ნებისმიერ ინდივიდს, რაც კი მისი მოქმედების არეში ხვდება.
ზოგად ელიმინაციას შეიძლება ჰქონდეს მუდმივი ხასიათი, თუ ის გარკვეულ სეზონთან არ არის დაკავშირებული, ან პერიოდული (წვიმები, გვალვი, ყინვები, პარაზიტების გამრავლება). იგი შეიძლება გარკვეულ ასაკს მოიცავდეს (კვერცხი, ჩანასახი, ზრდასრული ინდივიდი, თესლი, აღმონაცენი, მცენარე) - ასაკობრივი ზოგადი ელიმინაცია. ერთი შეხედვით, რაკი ზოგადი ელიმინაცია, არ ატარებს ამორჩევით ხასიათს, დამოკიდებულია ბრმა შემთხვევაზე და არა აქვს ორგანიზმის ინდივიდუალურ თვისებებს მნიშვნელობა, შეიძლება ვივარაუდოთ, რომ იგი არ არის დაკავშირებული ბუნებრივ გადარჩევასთან, მაგრამ ეს ასე არ არის. გადარჩევა ხდება შთამომავლობის შემთხვევითი გადარჩევით. საქმე იმაშია, რომ შემთხვევითი ელიმინაციის დროს, გადარჩენის მეტი შანსი აქვთ ნაყოფიერი წყვილის მრავალრიცხოვანი შთამომავლობის ერთეულ ინდივიდებს, ამ შემთხვევაში ბუნებრივი გადარჩევა მიმდინარეობს მაქსიმალური ნაყოფიერების მიმართულებით. ეს არის სტატისტიკური კანონზომიერების შედეგი - მრავალრიცხოვანი შთამომავლობის ინდივიდების გადარჩენის მეტი ალბათობაა, ვიდრე მცირერიცხოვანი შთამომავლობისა.
ასეთია შედარებით ნაკლებად თავდაცული პლანეტორული ორგანიზმების ელიმინაციის ხასიათი. მათი მაღალი ნაყოფიერება არის მაღალი ზოგადი ელიმინაციის შედეგი და მაჩვენებელიც. თუ ზოგად ელიმინაციას ასაკობრივი ხასიათი აქვს, მაშინ პოპულაციაში ასაკობრივი თანაფარდობა იცვლება. ზრდასრული ცხოველების მაღალი სიხშირით ამოწყვეტის შემთხვევაში ხდება ლარვული ფაზის გახანგრძლივება ზრდასრული სტადიის შემოკლების ხარჯზე. იგი გვხვდება ზოგიერთი მწერის სასიცოცხლო ციკლში. პირიქით, ლარვების მასიური ამოწყვეტისას ზრდასრული ფაზის გახანგრძლივება ხდება. ასეთია ჭიების, კიბოსნაირების, მწერების, მოლუსკების, ეკალკანიანების, თევზებისა და ამფიბიების სასიცოცხლო ციკლი, რომელთა ლარვები დიდი რაოდენობით იღუპება თევზებით, მტაცებელი მწერებითა და ფრინველებით. ყველა ამ შემთხვევაში ორგანიზმებს მაღალი ნაყოფიერებაც ახასიათებთ.
ინდივიდური ანუ ამორჩევითი ელიმინაცია. ინდივიდურ ელიმინაციას შეიძლება ჰქონდეს მუდმივი ან პერიოდული ხასიათი. ის შეიძლება იყოს განსხვავებული ასაკის მიხედვით. მაგრამ ყველა შემთხვევაში ჩვეულებრივ შეიმჩნევა უკეთ შეგუებული ფორმების გადარჩენა. ამორჩევითი ელიმინაცია ხდება ყველგან და ყოველთვის, სანამ ინდივიდური განსხვავებებია. არსებობის ოპტიმალურ პირობებში როგორც ლაბორატორიულ ასევე ბუნებრივ პირობებშიც ასეთი ელიმინაცია არასოდეს წყდება და ბუნებრივი გადარჩევა მიმდინარეობს.
ინდივიდური ელიმინაცია შეიძლება იყოს პირდაპირი, თუ ცალკეული ინდივიდები ფიზიკური ან ბიოლოგიური ფაქტორების ზემოქმედებით იღუპებიან. არაპირდაპირია თუ ცალკეული ინდივიდები ფიზიოლოგიური ფაქტორით (შიმშილით) სუსტდება და შემდეგში იღუპება სხვა ფაქტორებით, ან ხდება გამრავლებისაგან ჩამოშორება.
1. ფიზიკური ფაქტორებით გამოწვეული პირდაპირი ელემინაცია, ნიშნავს არახელსაყრელი კლიმატური ფაქტორების (ტემპერატურა, ნალექები, ქარი, წყლის დინებები, ზღვის ზვირთცემა და მისთ.) ზემოქმედებით ცალკეული ინდივიდის სიკვდილს. გადარჩებიან თავისი ზოგადი კონსტიტუციით ამ ფაქტორებისადმი მდგრადი ფორმები. მათ მოეპოვებათ სამარჯვეები (საფარველი რომელიც იცავს წყლის დაკარგვისა ან ტემპერატურის ზემოქმედებისაგან, ჯავშანი - იცავს მექანიკური დაზიანებისაგან და ა.შ.) რითაც თავს იცავენ ფიზიკური ფაქტორების მავნე ზემოქმედებისგან. ასევე გადარჩებიან ის ინდივიდები, რომლებიც მოძრაობითი რეაქციებით და ინსტინქტებით სწრაფად აღწევენ თავს ფიზიკური ფაქტორების ზემოქმედებას. მაღალი სიხშირით ამოწყდომა სხვა თანაბარ პირობებში დაკავშირებულია მაღალი ნაყოფიერების უნარის მფლობელ ინდივიდების გადარჩევასთან.
2. პირდაპირი ელიმინაცია ბიოლოგიური ფაქტორებით ნიშნავს ცალკეული ინდივიდების დაღუპვას მტაცებლებისაგან, პარაზიტებისა და ინფექციური დაავადებებისაგან. ამ დროს გადარჩებიან ის ინდივიდები, რომლებიც მეტად დაცულნი არიან თავისი ზოგადი აქტიურობით და ძალით, აგრეთვე აქტიური დაცვის სპეციალური საშუალებებით (შეიარაღება, გრძნობათა ორგანოები, გადაადგილების სიჩქარე), მეტიც დაცულები ფიზიოლოგიური საშუალებებით (იმუნიტეტი) ან პასიური თავდაცვის საშუალებებით (ნემსები, ეკლები, მფარველობითი შეფერილობა, სათანადო ინსტინქტები), გარდა ამისა, გადაირჩევიან აქტიური ინდივიდები, რომლებიც კარგად განვითარებულ გრძნობათა ორგანოებით, ინსტინქტებით და ნერვული მოქმედების უმაღლესი ფორმებით ხასიათდებიან, რის მეშვეობითაც ისინი მტაცებლებს თავს ადვილად არიდებენ, ამიტომაც ელიმინაციის ამ ფორმას დიდი მნიშვნელობა აქვს პროგრესულ ევოლუციაში, ორგანიზაციის მორფოფიზიოლოგიურ სრულყოფაში. ამოწყდომის მაღალი ხარისხი ამ შემთხვევაშიც იწვევს უფრო ნაყოფიერი ფორმების გადარჩევას, რომლებსაც განაყოფიერების საიმედო საშუალებები და კვერცხის შენახვისა და შთამომავლობის კვების და დაცვის საშუალებები აქვს.
3. არაპირდაპირი ელიმინაცია ფიზიოლოგიური ფაქტორებით ნიშნავს ორგანიზმის დასუსტებას შიმშილით, საკვები რესურსებისათვის კონკურენციაში, ნაყოფიერების დაქვეითებას და გამრავლებისაგან ჩამოცილებას (ნაწილობრივ სქესობრივი კონკურენციის შედეგად). ამ ტიპის ელიმინაციის ინტენსივობა იზრდება ინდივიდთა სიმჭიდროვის ზრდასთან ერთად („ჭარბდასახლება“).
ორგანიზმის დასუსტება შიმშილით (მეტწილად სეზონური) იწვევს მათ გაძლიერებულ ამოწყვეტას სხვა ფაქტორებით (ფიზიკური და ბიოლოგიური) ასე, რომ შიმშილი იშვიათად წარმოადგენს ცხოველის სიკვდილის უშუალო მიზეზს.
არაპირდაპირი ელიმინაცია, დაკავშირებული ნაკლებად დასუსტებული ორგანიზმების გადარჩენასთან, ნიშნავს ისეთი ინდივიდების გადარჩევას, რომელთაც წარმატებით შეუძლიათ საკვების მოპოვება, მეტად ეკონომიურად და პროდუქტიულად მისი გამოყენება. ეს იწვევს გრძნობათა ორგანოების განვითარებას, თავდასხმის საშუალებების, და სათანადო ინსტინქტების განვითარებას. ამავე დროს ეს ნიშნავს ისეთი ინდივიდების გადარჩევას, რომელთაც მაქსიმალურად შეუძლიათ საკვების ეკონომიკური მოპოვება და პროდუქტიულად გამოყენება, რაც დაკავშირებულია არა მარტო საჭმლის მომნელებელ ორგანოთა პროგრესთან, არამედ მთელი ორგანიზაციის, კერძოდ ფიზიოლოგიური კონსტიტუციის პროგრესთან. საკვების მაქსიმალური გამოყენება, როგორც წესი, ნიშნავს კვების სპეციალიზაციას და რეზერვების დაგროვების სხვადასხვა საშუალებების სრულყოფას. და ბოლოს, ნაკლებად დასუსტებული ინდივიდების გადარჩენა ნიშნავს ნაკლებ ნაყოფიერი ინდივიდების გადარჩევას, რომლებიც ნაკლებ ენერგიას ხარჯავენ სასქესო პროდუქტების წარმოქმნაზე. ამიტომ არაპირდაპირი ელიმინაცია იწვევს ნაყოფიერების დაქვეითებას (კომპენსაციის სახით იზრდება სიცოცხლის ხანგრძლივობა). ნაყოფიერების დაქვეითება დაკავშირებულია მცირერიცხოვანი შთამომავლობის სიცოცხლის მაქსიმალურ უზრუნველყოფასთან.
ოჯახური ელიმინაცია. ერთი წყვილის მთელი შთამომავლობის ამოწყვეტა არც თუ იშვიათი შემთხვევაა, ისე როგორც მთელი ოჯახების ამოწყვეტა, რადგან ერთად მცხოვრებთ ერთი და იგივე საშიშროება ემუქრებათ. მით უმეტეს, რომ ერთი წყვილის შთამომავლობა გენეტიკურად უფრო ერთგვაროვანია, ვიდრე პოპულაციის შემადგენელი ინდივიდები. ეს იმას ნიშნავს, რომ ყველა უპირატესობა, რომელიც იწვევს ცალკეული ინდივიდების გადარჩენას და ის უარყოფითი მომენტები, რაც იწვევს მათ დაღუპვას, ხშირად ვრცელდება ერთი წყვილის მთელ შთამომავლობაზე. ამიტომ ოჯახური ელიმინაცია გვხვდება იქაც, სადაც არ არის ორგანიზებული ოჯახი.
ოჯახური ელიმინაცია ხელს უწყობს ორგანიზებული ოჯახის ჩამოყალიბებას, ოჯახური ელიმინაციის დროს განსაკუთრებით ევოლუირებს ისეთი ნიშნები, როგორიცაა ზრუნვა შთამომავლობაზე, ბუდის შენების ინსტინქტები, შთამომავლობის დაცვა, კვება და ა.შ.
ჯგუფური ელიმინაცია. ასეთი ელიმინაციის საფუძველს წარმოადგენს გენეტიკური სიახლოვის გამო პოპულაციის ინდივიდებს შორის დიდი მსგავსება, ქვესახეობის ან სახეობის და სხვა ტოლფასოვან ჯგუფის ინდივიდებთან შედარებით.
ეს მსგავსება ერთი ბუნებრივი ჯგუფის ყველა წარმომადგენელს ერთნაირ მდგომარეობაში აყენებს არსებობისათვის ბრძოლაში. ამიტომ ელიმინაცია ხშირად ჯგუფის ყველა წარმომადგენელს ეხება, რომლებიც ნაკლებადაა შეგუებული საარსებო პირობებთან, ვიდრე სხვა ორგანიზმები. შედეგად ხდება მთელი ჯგუფის (პოპულაციის, სახეობის) ამოწყვეტა ან გაძევება კონკრეტული ტერიტორიიდან სხვა ფორმების მიერ, რომლებიც აქ წარმოიშვნენ ან სხვა ტერიტორიიდან მოვიდნენ.
ელიმინაციის ეს ფორმა გადარჩევის გამანადგურებელ და გამანაწილებელ როლზე მიუთითებს.
დიდი მნიშვნელობა აქვს ჯგუფური ელიმინაციის ინდივიდუალურ ელიმინაციასთან შეერთებას. თუ ყოველი ჯგუფი ცნობილ უპირატესობას ფლობს რეალურ პირობებში, ასეთი შეერთება, ჯგუფის ევოლუციას უზრუნველყოფს, იწვევს ნიშანთა დაცილებას და სხვადასხვა ბიოტოპების მიხედვით ფორმათა განაწილებას.
ბოლოს, ნაკლებად უპირატესი ჯგუფის ელიმინაცია და მისი გაძევება უფრო პროგრესული ფორმების მიერ ნიშნავს დიდი მაშტაბის ევოლუციას (მაკროევოლუციას), რომელიც განსაზღვრავს ორგანიზაციის არამარტო საერთო პროგრესს, არამედ ევოლუციის მექანიზმის გარდაქმნასაც. მაგრამ არ უნდა დაგვავიწყდეს ყოველი უპირატესი ჯგუფის პროგრესული განვითარება განისაზღვრება მისი ინდივიდების მხოლოდ ამორჩევითი ელიმინაციით.
|
|
10.1.3.3 8.3.3. შეჯიბრის ფორმები |
▲ზევით დაბრუნება |
ელიმინაციას ამორჩევითი ე.ი. ინდივიდუალური ხასიათი აქვს მხოლოდ პოპულაციის შიგნით გენეტიკურად განსხვავებული ინდივიდების შეჯიბრის შედეგად. შეჯიბრში ვლინდება არსებობისათვის ბრძოლის დადებითი მხარე, რაც დაკავშირებულია თვით ორგანიზმთან, მის აქტივობასთან, მის მისწრაფებასთან გამრავლებისაკენ და ბუნებრივი გადარჩევის შემოქმედებით როლთან.
ჯგუფშიდა შეჯიბრი. ჯგუფშიდა, ანუ ინდივიდუალური შეჯიბრი მიმდინარეობს ერთი პოპულაციის ცალკეულ ინდივიდებს შორის, მას შეიძლება ჰქონდეს უფრო აქტიური ხასიათი სიცოცხლის შენარჩუნების და გამრავლებისათვის ბრძოლაში კონკურენციისას, ხოლო უფრო პასიური გარემოს არასასურველ ფიზიკურ და ბიოლოგიურ ფაქტორებთან ბრძოლაში.
ინდივიდუალური აქტიური შეჯიბრი გამოიხატება სასიცოცხლო საშუალებებისათვის, ე.ი. საკვების (ჟანგბადი, სინათლე, წყალი და მასში გახსნილი მინერალური მარილები) კონკურენციაში. იგი მწვავდება მოსახლეთა სიმჭიდროვის მატებისას და პირდაპირ დამოკიდებულებაშია გამრავლების ენერგიასთან. აქტიური შეჯიბრი ძირითადად განისაზღვრება საკვების შეზღუდვით და ამდენად, ხშირად მძაფრდება არახელსაყრელ სეზონურ პირობებში (ზამთარში ან ზაფხულში). ზომიერი და ცივი სარტყლის ცხოველებში იგი განსაკუთრებით მძაფრდება ზამთარში (ან ზაფხულში გვალვებისას), როცა მკვეთრად მცირდება საკვები რესურსების მოპოვების საშუალება. აქტიური შეჯიბრება დაკავშირებულია არაპირდაპირ ელიმინაციასთან.
თუ საკვების უკმარისობა შიმშილს იწვევს, თუნდაც პერიოდულს, ბრძოლა არსებობისათვის მიმდინარეობს არა მარტო საკვების მოსაპოვებლად, არამედ მარაგის შესანახად თვით ორგანიზმის მიერ საკვები მასალისა და საკუთარი რეზერვების მეტად ეკონომიკურად გამოსაყენებლად. ეს დაკავშირებულია კვების სპეციალიზაციასთან, მორფოლოგიური კონსტიტუციის სრულყოფასთან და ზოგიერთ სპეციალური შეგუებების წარმოქმნასთან (ცხიმოვანი დეპო, მიგრაცია, ძილი). რადგანაც შეჯიბრების ამ ფორმაში გადარჩენა განისაზღვრება შიმშილის დროს ინდივიდის გამოფიტვით, დადებითი მნიშვნელობა აქვს სასქესო უჯრედებისა და კვერცხების პროდუქციის ეკონომიას. საბოლოო ჯამში შეჯიბრი საკვებისათვის იწვევს მორფოლოგიური კონსტიტუციის ბუნებრივ გადარჩევას, რაც დაკავშირებულია კვების სპეციალიზაციასთან და ნივთიერებათა ცვლის მეტად ეკონომიურობასთან, ასევე ნაყოფიერების დაქვეითებასთან, რაც შეიძლება კომპენსირებულ იქნეს სიცოცხლის გახანგრძლივებით და გამრავლების მრავალჯერადობით.
აქტიური კონკურენცია შეიძლება გამოვლინდეს შეჯვარებისათვის სქესობრივ კონკურენციაში. იგი დაკავშირებულია სქესობრივ გადარჩევასთან და მეორადი სასქესო ნიშნების და შესაბამისი ინსტინქტების ევოლუციას იწვევს.
ინდივიდუალური პასიური შეჯიბრება ვლინდება უშუალოდ ფიზიკურ და ბიოლოგიურ ფაქტორებთან ბრძოლაში სიცოცხლის შენარჩუნებისა და შთამომავლობის დატოვებისათვის. აქ საუბარია არასასურველი კლიმატური პირობების, მტაცებლების და პარაზიტების მიერ პირდაპირ ელიმინაციაზე. შეჯიბრს აქ პასიური ხასიათი აქვს, ვინაიდან ზოგიერთი ინდივიდის უპირატესობა თავდაცვის საშუალების არსებობით საერთო მტრისაგან, სხვა ინდივიდების ინტერესებს უშუალოდ არ ეხება. იგი არ მწვავდება პოპულაციის ინდივიდთა რაოდენობის გაზრდით, მაგრამ ეს არ ნიშნავს, რომ სიცოცხლისათვის და გამრავლებისათვის ბრძოლაში ორგანიზმი ნამდვილად პასიურია. პირიქით, გარკვეული აქტივობა ორგანიზმისა მის ზოგად დახასიათებას იძლევა არსებობისათვის ბრძოლის ყველა ფორმისას. თუმცა, მოცემულ შემთხვევაში არ არის ჭარბდასახლება და იგი არ განსაზღვრავს კონკურენციის სიმძაფრეს, მაგრამ იგი მაინც პირდაპირ დამოკიდებულებაშია გამრავლების ინტენსივობასთან. შესაბამისად, ორგანიზმის აქტიურობა გამოიხატება აქ თავისი სიცოცხლის დაცვისა და გამრავლებისაკენ მისწრაფებაში. პასიური შეჯიბრი დაკავშირებულია ბუნებრივ გადარჩევასთან და მეტად ეფექტური თავდაცვის საშუალებების გამომუშავებასთან. ამ შემთხვევაში მნიშვნელობა აქვს ძლიერ კონსტიტუციას, ორგანიზმის საერთო აქტიურობას და ძალას, გრძნობათა ორგანოების განვითარების ხარისხს, ინსტინქტებისა და სპეციალურ შეიარაღებას სიცოცხლის აქტიური დაცვისა და პასიური დაცვის საშუალებებით (ჯავშანი, მფარველობითი შეფერილობა და ფორმა).
პასიური შეჯიბრი გამრავლებასთანაცაა დაკავშირებული. რაც უფრო მეტად ხდება ფიზიკური და ბიოლოგიური ფაქტორებით კონკრეტული სახეების ამოწყვეტა, მით უფრო შანსი აქვთ შთამომავლობის დატოვებისა, ნაყოფიერ ინდივიდებს სასქესო ნიშნებისა და ინსტინქტების გამოსაცნობი ნიშნების განვითარებას, ნაყოფიერების გაზრდას და შთამომავლობის სიცოცხლის (ყვითრი, კვერცხის გარსები) უზრუნველყოფას.
ოჯახური შეჯიბრი. რადგანაც ერთი ოჯახის შთამომავლობა მემკვიდრულად უფრო მსგავსია ერთმანეთთან, ამიტომ სიცოცხლისათვის შეჯიბრში ისინი მსგავს პირობებში ვარდებიან. გადარჩენა ეხება მთელ ოჯახს. დიდ როლს ასრულებს ნაშიერთა სპეციალური დაცვის საშუალებები - ნაშიერთა ფარული განლაგება, გამოკვება, დაცვა მშობლების მიერ. ეს დაკავშირებულია შთამომავლობაზე ზრუნვის სხვადასხვა ფორმების ევოლუციასთან, რაც გამოიხატება, როგორც სპეციალურ ინსტინქტებში (ბუდობა, მოვლა), ისე კვერცხის, ჩანასახის და ნაშიერის დაცვაში (ჩანთები, ჯირკვლები).
ჯგუფური შეჯიბრი. ჯგუფთაშორისი შეჯიბრი გამოიხატება სხვა წევრთა კონკურენციაში საერთო საკვებისა და გამრავლების პირობებისათვის (აქტიური ფორმა შეჯიბრის) ან სხვადასხვა ჯგუფის ბრძოლა საერთო მტრებთან და ფიზიკურ პირობეთან (პასიური შეჯიბრი).
1. აქტიური შეჯიბრი ნიშნავს სხვა ჯგუფის ინდივიდთა კონკურენციას ერთნაირი საკვებისთვის (ნაწილობრივ, გამრავლების პირობებისათვის). ეს კონკურენცია არ აღწევს იმ სიმძაფრეს, როგორც ჯგუფის შიგნით. თუ ერთი ჯგუფის ინდივიდები უფრო ხელსაყრელ პირობებში აღმოჩნდებიან სასიცოცხლო პირობების ინტენსიურობასა და განსაზღვრულობის შემთხვევაში, მაშინ განუსაზღვრელი გამრავლებისაკენ (ჯგუფის შიგნით) მისწრაფებისას ხდება მოცემული ადგილიდან მეორე ჯგუფის მთელი პოპულაციის განდევნა. ეს ეხება ძირითადად გეოგრაფიულ ფორმებს ან ქვესახეობებს.
2. პასიური შეჯიბრი, სხვადასხვა ჯგუფის ინდივიდთა ბრძოლა საერთო მტერთან და არახელსაყრელ ფიზიკურ ფაქტორებთან.
თუ ჯგუფის აქტიური შეჯიბრის საფუძველს საარსებო საშუალებების ერთიანობა წარმოადგენს, პასიური შეჯიბრის საფუძველი საერთო საშიშროებაა, რომელიც სხვადასხვა ჯგუფის ინდივიდთა სიცოცხლეს ემუქრება. ეს უფრო მეტად მონათესავე ჯგუფებს ეხება, რადგანაც დაცილებულ ჯგუფებს მტრებიც განსხვავებული ჰყავთ. ამ შემთხვევაში ჯგუფი, რომელიც გარკვეულ უპირატესობას ფლობს, დევნის მეორე ფორმას. მაგრამ აქ ერთი ჯგუფის მიერ მეორე ჯგუფის განდევნა ტერიტორიიდან არ ნიშნავს ამ ჯგუფის აუცილებელ დაღუპვას, რადგანაც სხვა საარსებო პირობებში მეორე ჯგუფისათვის შეიძლება უფრო ხელსაყრელი პირობები იყოს. ამიტომ ჯგუფთაშორის შეჯიბრში ვლინდება გადარჩევის არა მარტო გამანადგურებელი, არამედ გამანაწილებელი როლიც.
ინდივიდუალური და ჯგუფთაშორისი შეჯიბრის გაერთიანებას დიდი მნიშვნელობა აქვს ნიშანთა დაცილების პროცესში. რაც დაკავშირებულია დაცილებული ფორმების განაწილებასთან სხვადასხვა ოლქებს, ბიოტოპებსა და ეკოლოგიურ ნიშებში.
ჯგუფთაშორისი შეჯიბრი ორგანიზაციის საერთო პროგრესს იწვევს, რაც უფრო სრულყოფილი ფორმების მიერ მოძველებული ორგანიზაციის ფორმის განდევნასთან არის დაკავშირებული. ჯგუფთაშორისი შეჯიბრი იწვევს აგრეთვე ევოლუციის ფაქტორთა ევოლუციას, რადგანაც დაკავშირებულია ისეთი ფორმების განდევნასთან, რომლებიც ევოლუციის ტემპების და ხარისხის მხრივ ჩამორჩებიან. მაგრამ ამ ცვლილებათა საფუძველს მაინც ჯგუფშიდა შეჯიბრი წარმოადგენს, რადგანაც იგი იწვევს მოცემული ჯგუფების უპირატესობას სხვა ჯგუფებთან ბრძოლაში. მეორე მხრივ, მხოლოდ ჯგუფთაშორისი შეჯიბრის დროს ყალიბდება ნიშნები, რომლებიც მთელი ჯგუფების სიცოცხლეს და განსახლებას უწყობენ ხელს, მაგრამ არ აძლევენ ინდივიდებს უპირატესობას მათ ინდივიდურ ცხოვრებაში (მაღალი ნაყოფიერება, შთამომავლობაზე ზრუნვა, სიცოცხლის მცირე ხანგრძლივობა და ა.შ.).
ევოლუციის პროცესში არსებობისათვის ბრძოლას დარვინი განიხილავდა, როგორც ბუნებრივი გადარჩევის წანამძღვარს. არსებობისათვის ბრძოლის ერთგვარ ობიექტურ საზომს წარმოადგენს სხვაობა ჩასახულ და ზრდასრულ ინდივიდებს შორის. არსებობისათვის ბრძოლა რთულ ფენომენს წარმოადგენს, რომელსაც ბიოლოგიის ფუნდამენტური დარგი - ეკოლოგია შეისწავლის.
|
|
10.2 ბუნებრივი გადარჩევა და მისი ძირითადი ფორმები |
▲ზევით დაბრუნება |
|
|
10.2.1 8.4. ბუნებრივი გადარჩევის არსი |
▲ზევით დაბრუნება |
ბუნებრივი გადარჩევა ევოლუციის მთავარი მამოძრავებელი ფაქტორია. მოძღვრება ბუნებრივი გადარჩევის შესახებ ევოლუციის სინთეზური თეორიის ქვაკუთხედს წარმოადგენს. როგორც აღინიშნა, ბუნებრივი გადარჩევის თეორია ჩამოაყალიბა ჩ.დარვინმა (1837-58). მისი არსებობა დარვინისაგან დამოუკიდებლად დაადგინა ა. უოლესმა (1858). ჩ. დარვინის განმარტებით, ბუნებრივი გადარჩევა არის სასარგებლო ინდივიდუალურ განსხვავებათა ან ცვლილებათა მქონე ინდივიდების შენარჩუნება და საზიანოთა მოსპობა. დარვინის მიხედვით ბუნებრივი გადარჩევა არსებობისათვის ბრძოლის შედეგია.
მუტაციური და კომბინაციური ცვალებადობის შედეგად ნებისმიერი პოპულაცია ჰეტეროგენულია. მუტაციური ცვალებადობა იძლევა მასალას ბუნებრივი გადარჩევისათვის. პოპულაციაში ინდივიდთა ჰეტეროგენულობა და გამრავლების პროგრესია განაპირობებს სასიცოცხლო საშუალებათა (საკვები, თავშესაფარი და მისთ.) ნაკლებობას და წარმოადგენს არსებობისათვის ბრძოლის წანამძღვარს, რომლის დროსაც პოპულაციის შემადგენელ ინდივიდთა ერთი ნაწილი იღუპება (ან ვერ ტოვებს შთამომავლობას), ნაწილი კი გადარჩება. მეტწილად იღუპებიან საზიანო ვარიაციის მქონე ინდივიდები, ხოლო გადარჩებიან სასარგებლო ვარიაციის მქონენი. ე.ი. ადგილი აქვს დიფერენციალურ სიკვდილიანობას. გადარჩენილი ინდივიდები ნიშან-თვისებებს გადასცემენ შთამომავლობას. ყოველ მომდევნო თაობაში სასარგებლო ვარიაციების წილი გაცილებით მეტი იქნება, ვიდრე წინა თაობაში იყო. ამ პროცესის მრავალჯერადი განმეორება თაობათა მანძილზე (ბიოლოგიურ დროში) იწვევს საზიანო ვარიაციათა მქონე ინდივიდების ელიმინაციას და სასარგებლო ვარიაციათა მქონე ფორმების გადარჩენას, რაც უზრუნველყოფს არსებობისათვის ბრძოლაში გამარჯვებას.
ერთი და იმავე პოპულაციის ინდივიდები განსხვავებული რაოდენობით იძლევიან შთამომავლობას. დავუშვათ, რომ ერთსა და იმავე საარსებო პირობებში გავრცელებული ინდივიდები მხოლოდ ამ ნიშნით განსხვავდებიან. მშობელთა რომელი წყვილის შთამომავალი მიაღწევს რეპროდუქციულ ასაკს? ბუნებრივია, მშობელთა იმ წყვილის, რომელიც შედარებით მეტ შთამომავლობას იძლევა, რადგანაც მეტია ალბათობა, რომ რომელიმე მათგანი ზრდასრულ ასაკს მიაღწევს და შთამომავლობას დატოვებს. ამ პროცესს დიფერენციალური გამრავლება ეწოდება. ამრიგად, ბუნებრივი გადარჩევის პროცესში მთავარია ინდივიდთა დიფერენციალური სიკვდილიანობა, დიფერენციალური გამრავლება და არა ინდივიდთა გაჩენა ან დაღუპვა. ევოლუციაში პოპულაციის გენეტიკური სტრუქტურის (გენოფონდის) ფორმირებაში მთავარი როლი ენიჭება ინდივიდის გენოტიპს. გენოფონდში იმ ინდივიდს შეაქვს მეტი წვლილი, რომელიც დიდი რაოდენობით ტოვებს რეპროდუქციული უნარის მქონე შთამომავლობას. გამრავლების გარეშე რაგინდ დიდხანს უნდა იარსებოს ინდივიდმა, იგი არავითარ როლს არ ასრულებს გენოფონდის ფორმირებაში. იგივე ითქმის იმ ინდივიდებზე, რომლებიც რეპროდუქციულ ასაკამდე დაიღუპნენ. გარემოს მალიმიტირებელი ფაქტორებისა და პოპულაციის რიცხოვნობის ერთდროული მოქმედება ბუნებრივი გადარჩევის ზეწოლას იწვევს. მისი მოქმედებით ამა თუ იმ ალელის გავრცელება პოპულაციაში დიდდება ან იზღუდება. გენოფონდში ალელის სიხშირის ცვლილებამ შეიძლება ევოლუციური ცვლილება გამოიწვიოს. პოპულაციაში გარკვეული ალელების ან გენთა კომპლექსის წარმატებული გავრცელება და გამყარება ელემენტარულ ევოლუციურ მოვლენას იწვევს. იგი ნებისმიერი ევოლუციური პროცესის საფუძველს წარმოადგენს.
ბუნებრივი გადარჩევის გენეტიკურ-ევოლუციური არსი იმით გამოიხატება, რომ პოპულაციაში დიფერენცირებულად (არაშემთხვევითად) ინახება გარკვეული გენოტიპები, ასევე მომდევნო თაობას გენები შერჩევით გადაეცემა. ამრიგად, ბუნებრივი გადარჩევა არის უკეთ შეგუებული ინდივიდების გადარჩენა და შთამომავლობის დატოვება ერთ კონკრეტულ საარსებო გარემოში გარკვეული გენოტიპების შერჩევითი შენარჩუნება და მომდევნო თაობებისათვის გადაცემა.
|
|
10.2.2 8.5. ბუნებრივი გადარჩევის ობიექტი და მოქმედების სფერო |
▲ზევით დაბრუნება |
ბუნებრივი გადარჩევა მჭიდროდაა დაკავშირებული არსებობისათვის ბრძოლასთან და მთლიანად მასზეა დამოკიდებული. იგი არის ელიმინაციის საწინააღმდეგო და შეჯიბრით განპირობებული პროცესი. მხოლოდ ბუნებრივი გადარჩევა წარმოადგენს მიმართულ ფაქტორს, რომელიც ინახავს უკეთ შეგუებულ ვარიანტებს (გენოტიპებს) და თავს უყრის ერთიან გენეტიკურ სისტემაში - გენოფონდში. იგი უზრუნველყოფს ნაირგვარ ინდივიდურ და ჯგუფურ (სახეობრივ) ადაპტაციათა ჩამოყალიბებას, ასევე წარმართავს და აორგანიზებს მთლიანად მიკროევოლუციის პროცესს (შმალგაუზენი, 1990).
დარვინმა ბუნებრივი გადარჩევა განსაზღვრა როგორც სასარგებლო ინდივიდური გადახრების მფლობელ ინდივიდების გადარჩენა და საზიანო გადახრების მქონეთა ელიმინაცია. მაგრამ ეს ფორმულირება გადარჩევის მნიშვნელოვან გენეტიკურ შედეგებს ვერ გამოხატავს. დარვინი ხშირად იყენებდა გამოთქმას „უკეთ შეგუებულთა გადარჩევას“ და ამიტომ მას ტავტოლოგიაში დებენ ბრალს. „რომელი ინდივიდი გადარჩება? - მეტად შეგუებული; რომელია მეტად შეგუებული? - რომელიც გადარჩება“. მიიჩნევენ, დარვინი შეგუებულობაში გულისხმობდა, რომ უკეთეს გენოტიპს გადარჩენისა და შთამომავლობის დატოვების მეტი შანსი მოეპოვება, ვიდრე უარეს გენოტიპს.
როგორც უკვე აღვნიშნეთ ბუნებრივი გადარჩევის პროცესში მთავარი მნიშვნელობა ინდივიდთა დიფერენცირებულ გამრავლებას ენიჭება. პოპულაციის გენოფონდის ფორმირებაში იმ ინდივიდს შეაქვს მეტი წვლილი, რომელიც დიდი რაოდენობით ტოვებს სქესმწიფე შთამომავლებს. ამრიგად ბუნებრივი გადარჩევა არის პროცესი, რომელიც განსაზღვრავს ინდივიდების დიფერენცირებულ წვლილს მომდევნო თაობის ჩამოყალიბებაში. იგი გენოტიპთა ნაირგვარი ვარიანტებიდან ამოარჩევს და ინახავს უკეთეს ფორმებს. გენოფონდის ფორმირებაში ამდაგვარ შეფარდებით წვლილს შეგუებულობა ანუ ადაპტური ღირებულება ეწოდება. გენოფონდის ფორმირებაში მონაწილე შთამომავლობის რაოდენობა მრავალ ფაქტორზეა დამოკიდებული (პოპულაციაში რიცხოვნობის ციკლური მერყეობა, წელიწადის სეზონური ცვლილებები, კლიმატური პირობების მერყეობა, საკვების რაოდენობა და ხარისხი და სხვ.). დროის ნებისმიერ მონაკვეთში პოპულაციის ზოგიერთი წევრი სხვებთან შედარებით გაცილებით მეტ შთამომავალს ტოვებს.
გადარჩევის ობიექტია არა ცალკეული გენი, არამედ მათი ერთობლიობა - გენოტიპი, მისი მფლობელი ინდივიდი. გენოტიპში არსებული ინფორმაცია ინდივიდური განვითარების პროცესში (ონტოგენეზში) ფენოტიპად რეალიზდება. სხვადასხვა გენოტიპები ადაპტური ღირებულებით ერთმანეთისგან განსხვავდებიან. ბუნებრივი გადარჩევის მოქმედებით არაადაპტური გენების მფლობელი გენოტიპები იცხრილება, რის გამოც გენოფონდში თანდათანობით ადაპტური ღირებულების მფლობელი გენოტიპების კონცენტრაცია იზრდება. გადარჩევის კონტროლს ექვემდებარება აგრეთვე ოჯახი. დემი, დემთა ჯგუფი, პოპულაცია, პოპულაციათა ჯგუფი და ბოლოს მთლიანად ეკოსისტემა, მაგრამ მხოლოდ პოპულაციის საზღვრებში ხდება ინდივიდთა გენოტიპების გადარჩევა. გენოფონდი ფორმირდება ძირითადად ისეთი გენოტიპებით, რომელთაც სხვებთან შედარებით ადაპტური უპირატესობა მოეპოვებათ. მაშასადამე პოპულაცია წარმოადგენს ევოლუციის ელემენტარული ფაქტორის ბუნებრივი გადარჩევის მოქმედების არეს.
უნდა აღინიშნოს, რომ ბუნებრივი გადარჩევა გენსა და გენოტიპზე უშუალოდ არ მოქმედებს. იგი არაპირდაპირ - ფენოტიპის გზით მოქმედებს გენოტიპზე. პოპულაციაში გენთა სიხშირის ცვლილება ფენთა სიხშირის ცვლილებით ვლინდება. როდესაც გენოტიპური ცვალებადობა ფენოტიპურად არ ვლინდება (რეცესიული ალელები ჰეტეროზიგოტურ მდგომარეობაში, გენის პენეტრანტულობა), მაშინ მასზე ბუნებრივი გადარჩევა ვერ მოქმედებს. გადარჩევის საფეხურებრივი მოქმედება, გენოტიპების გადარჩევა ფენოტიპების მეშვეობით, განსაზღვრავს ევოლუციაში ფენოტიპისა და ფენოტიპური ცვალებადობის მნიშვნელობას.
ნებისმიერი ადაპტაცია ამა თუ იმ ფენის სახით ვლინდება, რომელიც ინდივიდური განვითარების პროცესში ყალიბდება. ამდენად ევოლუციური თვალთახედვით მეტად მნიშვნელოვანია გენოტიპის ფენოტიპად რეალიზაციის პროცესი. ნებისმიერი მუტაცია, რომელიც უკეთ იცავს ორგანიზმს გარემო ფაქტორების უარყოფითი ზემოქმედებისაგან ან მეტაბოლიზმის პროცესში შესაძლო შეცდომებისგან, ადაპტაციის გაძლიერებასა და სრულქმნას იწვევს. არსებობისათვის ბრძოლა გენებსა და ფენებს შორის როდი მიმდინარეობს, არამედ მათ მფლობელ ინდივიდებს შორის. მაშასადამე ინდივიდები ბუნებრივი გადარჩევის ელემენტარულ ობიექტს წარმოადგენენ.
ბუნებრივი გადარჩევის დამამტკიცებელი არგუმენტები ორი კატეგორიისაა: 1. მათემატიკური ანალიზით დამაჯერებლად მტკიცდება, რომ მცირე სელექციური უპირატესობაც კი იწვევს პოპულაციაში იმ გენების დაგროვებას, რომლებიც კონკრეტულ ადაპტურ ნიშან-თვისებას განსაზღვრავენ (რ. ფიშერი, ს. რაიტი, ჯ. ჰოლდეინი და სხვ.). 2. ნატურალისტების მიერ მოპოვებული მონაცემები, რომლითაც ბუნებრივი გადარჩევის მოქმედება დასტურდება. (იხ. §8.4).
ანტიდარვინისტები უარყოფენ გადარჩევის სტატისტიკურ ხასიათს. მათი შეხედულებით ინდივიდთა სიკვდილიანობა მეტწილად შემთხვევითია და სელექციასთან (არსებობისათვის ბრძოლა) კავშირი არ გააჩნია. ანალოგიისათვის ასახელებენ შემდეგ მაგალითს, დიდი თევზი ან ვეშაპი პლანქტონით კვებისას არ არჩევს ცხოველებს. მათი განადგურება შემთხვევითობაზეა დამოკიდებული. ამასთანავე ისინი გვერდს უვლიან იმ ფაქტს, რომ გადარჩენილი ინდივიდების წარმატებული რეპროდუქცია სელექციურ ფაქტორებზეა დამოკიდებული. შეგუების შეფარდებითი მიზანშეწონილობა ასევე ბუნებრივი გადარჩევის არასელექციურ როლად მიაჩნიათ.
ე. მაირი სამართლიანად აღნიშნავს, რომ სელექციონისტები (პოპულაციონისტები) როდი აიდიალებენ ბუნებრივი გადარჩევის როლს, არავინ ამტკიცებს, რომ გადარჩევა უკვდავებას იწვევს. იგი მხოლოდ განსაზღვრავს პოპულაციის ამა თუ იმ წევრის გადარჩენადობას და ადაპტურ ღირებულებას. ეს ფაქტორი თავისი მოქმედებით ხელს უწყობს შეფარდებით წარმატებულ გამრავლებას. ნებისმიერი გენი, რომელიც ამ პროცესზე უშუალოდ ან არაპირდაპირ დადებითად მოქმედებს. გადარჩევის მოქმედებით ხდება მათი გამყარება პოპულაციაში.
ანტიდარვინისტები ეჭვს გამოთქვამენ გენთა სელექციურ ღირებულების მიმართაც, კერძოდ იმ გენებზე, რომლებიც უმნიშვნელოდ ზრდიან შეგუებულობას. მათ მხედველობიდან რჩებათ ის ფაქტი, რომ გადარჩევა მოქმედებს არა ცალკეულ გენზე არამედ ფენოტიპზე. პოპულაციაში თანდათანობით (გრადუალურად) ხდება სელექციური ღირებულების ნიშნების განმსაზღვრელი გენების კუმულაცია. საილუსტრაციოდ განვიხილოთ მოდელური პოპულაცია, რომელსაც 100 გაუმყარებელი (არაფიქსირებული) პოლიალელური ლოკუსი მოეპოვება. დავუშვათ, რომ ცალკეული ალელი უმნიშვნელოდ მოქმედებს შეგუებულობაზე. გარემოსა და სხვა ფაქტორთა გავლენით ალელი შეიძლება ფლობდეს დადებით ან უარყოფით სელექციურ ღირებულებას. რეკომბინოგენური ცვალებადობისა და გარემოს კონკრეტული ფაქტორების გავლენით ინდივიდებს განსხვავებული რაოდენობით მოეპოვებათ, როგორც სასარგებლო ისე საზიანო ლოკუსები. იმ ინდივიდებს, რომელთაც მეტი დადებითი ალელები აქვთ, წარმატებული გამრავლებისა და სქესმწიფე შთამომავლობის დატოვების მეტი შანსი გააჩნიათ. პოპულაციის ის წევრები, რომელთაც ძირითადად უარყოფითი ალელები (დაბალი ადაპტური ღირებულება) გააჩნიათ იღუპებიან. პოპულაციის ყოველ მომდევნო თაობაში უარყოფითი გენების სიხშირე კლებულობს, ხოლო დადებითი გენების კონცენტრაცია მატულობს.
ბუნებრივი გადარჩევის მოქმედებას თან ახლავს საზიანო ვარიაციათა მქონე ინდივიდების ელიმინაცია, ჯ. ჰოლდეინის ხატოვანი გამოთქმით, საზღაური გადარჩევისათვის. რამდენიმე გენის შეცვლის საფუძველზე ჩატარებული მათემატიკური ანალიზით გამოირკვა, რომ ეს სიდიდე საკმაოდ მაღალია. მიღებული შედეგის საფუძველზე გამოტანილია პარადოქსული დასკვნა. მრავალი გენის შეცვლის საფუძველზე შეუძლებელია სწრაფი ევოლუციური ცვლილება. ეს პარადოქსი ადვილად იხსნება ვინაიდან გადარჩევას ექვემდებარება ფენოტიპი და არა გენები.
ბუნებრივი გადარჩევის მოქმედების სფეროს შეფასებისას საჭიროა იმის გათვალისწინება, რომ ნიშან-თვისებები, რომელზედაც გადარჩევის ზეწოლა მოქმედებს, უშუალოდაა დაკავშირებული რეპროდუქციის პროცესთან და წარმატებული გამრავლების შესაძლო ზრდასთან. ზოგჯერ ევოლუციის პროცესში „ფასეულობათა გადაფასება“ მიმდინარეობს. კერძოდ შესაძლებელია ნეიტრალური ან საზიანო ნიშანი სხვა კონკრეტულ გარემოში ადაპტური აღმოჩნდეს. ამდაგვარი ადაპტური ნიშანი პოპულაციაში ვრცელდება და მყარდება.
გადარჩევის მოქმედების სფეროს ერთი შეზღუდვა მოეპოვება. ბუნებრივ გადარჩევას არ შეუძლია კონკრეტული სახეობის ორგანიზაციის შეცვლა იმგვარად, რომ მფლობელისთვის უსარგებლო აღმოჩნდეს, ხოლო სარგებლობა მხოლოდ სხვა სახეობისათვის ჰქონდეს. ამ სახის ცვლილებას თვით ბუნებრივი გადარჩევის პრინციპი „კრძალავს“. იგი მოქმედებს იმ ნიშან-თვისებათა სასარგებლოდ, რომელიც ხელს უწყობს ორგანიზმის წარმატებულ გამრავლებას.
ბუნებრივი გადარჩევა ხშირად იწვევს ისეთი ნიშნების ჩამოყალიბებასა და გამყარებას პოპულაციაში, რომელიც ცალკეული ინდივიდისათვის საზიანოა, მაგრამ სარგებლობა მოაქვს პოპულაციისა და მთლიანად სახეობისათვის (მაგალითად, ალტრუისტული ქცევა, რომელიც მოეპოვება სოციალურ ცხოველებსა და ადამიანს; ორაგულისებრთა ზოგიერთ სახეობაში ტოფობის შემდეგ ინდივიდების სიკვდილი და მისთ.).
ევოლუციის პროცესში სულ უფრო იზრდება სელექციური მექანიზმების როლი, რომელიც ვლინდება ერთი სახეობის ან სხვადასხვა სახეობის ინდივიდთა შორის. სახეობათაშორისი ურთიერთობანი უფრო მეტად კვებითი კავშირების ხარჯზე ყალიბდება, მაშინ როცა შიდა სახეობრივ ურთიერთობაში წამყვანი როლი გამრავლების ფუნქციას ენიჭება. ჯგუფური გადარჩევის კონტროლით სახეობის შიგნით სხვადასხვა ჯგუფური ადაპტაციები (სქესობრივი, შთამომავლობაზე ზრუნვასთან დაკავშირებული ადაპტაციები და მისთ.) ფორმირდება. მათ დიდი მნიშვნელობა არა აქვთ ცალკეული ინდივიდებისათვის, გაცილებით მნიშვნელოვანია ჯგუფისათვის. ასეთ ადაპტაციებს ს. სევერცოვმა (1951) კონგრუენციები უწოდა.
გადარჩევა არის ფაქტორი, რომელიც პოპულაციის გენეტიკური სტრუქტურის სწრაფ შეცვლას იწვევს. რა მექანიზმების საშუალებით ხდება გენოტიპურად განსხვავებული კლასების შეგუებითი ღირებულების ჩამოყალიბება?
ადაპტური ღირებულების მახასიათებლად, პირველ რიგში, გადარჩენადობა და გამრავლების უნარი წარმოგვიდგება. გადარჩენადობა განისაზღვრება განვითარების სისწრაფით, სხვადასხვა დაავადების მიმართ გამძლეობით, არასასურველი ზემოქმედების გადატანისა და გარემოს მავნე ფაქტორების თავიდან აცილების უნარიანობით, საკვების მოპოვების უნარით და საერთოდ სასიცოცხლო აქტივობით და სხვ. გამრავლების კვლავწარმოების რეპროდუქციული უნარი დამოკიდებულია სქესობრივი მომწიფების სისწრაფეზე, კონკურენციის გამძლეობაზე, ფერტილობაზე, სქესობრივ აქტივობასა და ქცევაზე. როგორც ყველა ჩამოთვლილი თვისება ისე დანარჩენი ნიშან-თვისება გენოტიპის კონტროლით ყალიბდება. ონტოგენეზში მემკვიდრული ინფორმაციის რეალიზაცია გარემოს გავლენით გარკვეული მოდიფიცირებით (რეაქციის ნორმის ფარგლებში) ხორციელდება.
რაც უფრო მაღალ ევოლუციურ საფეხურზე დგას ორგანიზმი, მით მეტ ხვედრით წონას იძენს ადაპტაციის სხვადასხვა ონტოგენეზური მექანიზმები. უმაღლეს მცენარეებს, ფართო მოდიფიკაციური ცვალებადობის უნარი, საშუალებას აძლევს სხვადასხვა საარსებო პირობებში განსახლდნენ და დაიკავონ სხვადასხვა ეკოლოგიური ნიში. ცხოველებში ნატიფი ადაპტაცია ონტოგენეზში გარემოს პირობებთან ძირითადად მიიღწევა ქცევითი რეაქციების პლასტიკურობით. ი. შმალგაუზენის (1969) მიხედვით, გრძნობათა ორგანოების განვითარება, ნერვული სისტემის ცენტრალიზაცია და უმაღლესი საკოორდინაციო ცენტრების განვითარება ისეთი თვისებების განვითარებას ნიშნავს, რომლებიც დიდ უპირატესობას ანიჭებენ ორგანიზმებს არსებობისათვის ბრძოლაში და თავის მნიშვნელობას ინარჩუნებენ ნებისმიერ პირობებში, ნებისმიერ გარემოში და მის ნებისმიერ ცვლილებისას.
|
|
10.2.3 8.6. ბუნებრივი გადარჩევის დამამტკიცებელი არგუმენტები |
▲ზევით დაბრუნება |
XIX საუკუნის ბოლოს და XX ს-ში მრავალი ექსპერიმენტული გამოკვლევა ჩატარდა ჩ. დარვინისა და ა. უოლესის მიერ მოწოდებული ბუნებრივი გადარჩევის ჰიპოთეზის დასასაბუთებლად. თვალსაჩინო არგუმენტები მხოლოდ XXს-ის შუა პერიოდში იქნა მოპოვებული. კერძოდ, დასაბუთდა, რომ ბუნებრივი გადარჩევა მთავარი და ერთადერთი მიმართული ევოლუციური ფაქტორია, რომელიც გადამწყვეტ როლს ასრულებს ორგანული სამყაროს ისტორიულ განვითარებაში. ყოველივე ზემოთ აღნიშნულმა განაპირობა ამ ჰიპოთეზის ბუნებრივი გადარჩევის თეორიად ჩამოყალიბება.

სურ. 8.2. არყის ხის მზომელას პეპლების ნორმალური და მელანისტური შეფერილობა. ა - მღიერებიან ღია შეფერილობის ქერქის ფონზე. ბ - გაჭვარტლულ ხის ქერქის ფონზე (ნ. იორდანსკი, 2002).
კლასიკური მაგალითია ინდუსტრიული მელანიზმის მოვლენა. XIX-XXსს. განვითარებულ ინდუსტრიულ რაიონებში ქარხნების გამონაბოლქვმა გარემოს დაბინძურება გამოიწვია. მცენარეული საფარი გაიჭვარტლა. ასეთ რაიონებში წარმოიქმნენ მუქი შეფერილობის მწერები. მათ პოპულაციიდან განდევნეს ღია ფორმები. ევროპაში პეპლების 80-ზე მეტ სახეობას შეფერილობა შეეცვალა. მათი შემადგენელი პოპულაციები ძირითადად მუქი შეფერილობისაა. ეს მოვლენა კარგადაა შესწავლილი არყის ხის მზომელას პეპლებში (Biston betularia) (სურ. 8.2). ბირმინგემის მიდამოებში მუქი შეფერილობის პეპელა (მელანისტური ფორმა) პირველად 1848 წელს ნახეს, 1953 წელს კი მთელი პოპულაციის 90%25 მუქი შეფერილობის ინდივიდებისაგან შედგებოდა. საწყისი შეფერილობის პეპლებს აქვთ ღია მონაცრისფრო-კრემისფერი, შავ წინწკლებიანი ფრთები (ფორმა ტყპიცალ). ისინი ღამის პეპლებია, დღისით უძრავად სხედან არყის ხის ქერქზე. მათი შეფერილობა ეხამება არყის ხეზე არსებულ მღიერებს. ბუნებაში გვხვდება მუქი რუხი (f. insularia) და მურა შავ ფრთებიანი (f.carbonaria) ინდივიდები. ჩატარებული გენეტიკური ანალიზით დადგინდა, რომ მურა შავი შეფერილობის განმსაზღვრელი ალელი (Cc) დომინირებს მუქ ნაცრისფერისაზე (Ci), ორივე მათგანი კი ღია შეფერილობისაზე (c). ე.ი. Cc>Ci>c. ასი წლის განმავლობაში ბირმინგემის მიდამოებში საწარმოო ნარჩენებით ტყეები იმდენად შეიჭვარტლა, რომ არყის ხეზე მღიერები მოისპო, ხოლო ქერქი შავი შეფერილობის გახდა. ამ ფონზე მუქ ფორმებს მფარველობითი შეფერილობა აღენიშნათ. ისინი ნაკლებ შესამჩნევნი არიან. ბუნებრივი გადარჩევა ფრინველთა მეშვეობით ხდება: ადვილად ამჩნევენ და ჭამენ ღია შეფერილობის პეპლებს. არასამრეწველო რაიონებში საპირისპირო მოვლენა შეინიშნება. ინგლისის ძლიერ დაბინძურებულ რაიონებში გვხვდება - carbonaria, შედარებით ნაკლებ დაბინძურებულში - insularia, ხოლო არაინდუსტრიულ რაიონებში, სადაც ტყეები დაუბინძურებელია - typical.
ბ. ქეთლუელმა (1958) შეისწავლა გადარჩევის გავლენა პეპლების მელანისტური ფორმების გავრცელებაზე. მან ბირმინგემის მახლობლად ძლიერ გაჭვარტლულ ტყეში გაუშვა საღებავით მონიშნული სამივე ფორმის პეპლები, შემდეგ მათ სატყუარათი იჭერდა და აღრიცხავდა გადარჩენილი ინდივიდების რაოდენობას. გამოირკვა, რომ ფრინველები ღია შეფერილობის პეპლებს ორჯერ მეტს ანადგურებდნენ, ვიდრე მუქ ფორმებს. ანალოგიური ცდა მან ჩაატარა დორსეტში, სადაც ტყეები გაჭვარტლული არაა. არყის ხის თეთრ ფონზე ფრინველები ადვილად ამჩნევდნენ მუქ პეპლებს და ისინი ორჯერ მეტი ნადგურდებოდა, ვიდრე ღია ფორმები. მას შემდეგ, რაც გარემოს დაცვას გაჭუჭყიანებისაგან დიდი ყურადღება მიექცა, კერძოდ ქვანახშირის გამოყენება საწვავად მკვეთრად შემცირდა, ადგილი აქვს უკუპროცესს.
მღრღნელებში და მწერებში შხამქიმიკატებისადმი, ხოლო პათოგენურ მიკროორგანიზმებში ფარმაკოლოგიური პრეპარატებისადმი რეზისტენტულობის ამაღლება გადარჩევის წამყვანი როლის დამადასტურებელი მნიშვნელოვანი არგუმენტია. 1944 წელს დაავადების გადამტან და სასოფლო-სამეურნეო მავნებლების წინააღმდეგ ფართო მაშტაბით გამოიყენეს ინსტექტიციდი დდტ. 1960 წლისთვის უკვე გამოავლინეს ამ შხამქიმიკატისადმი რეზისტენტული 120 სახეობის მწერი, მათ შორის 60 სახეობა მედეგი აღმოჩნდა რამდენიმე ინსექტიციდის მიმართ. ორფრთიანებში ჩატარებული გამოკვლევებით გამოირკვა, რომ დდტ-ს ზემოქმედებით 4-7 თაობის განმავლობაში ამ შხამქიმიკატისადმი მედეგობა 100-ჯერ იზრდება. ისინი უძლებენ ისეთ დოზებსაც კი, რომელიც საწყისი ფორმების 100%25-ით სიკვდილს იწვევდა. შხამქიმიკატის ზემოქმედების შედეგად გადარჩებიან ერთეული რეზისტენტული ინდივიდები. გამრავლების პოტენციალის რეალიზაციის შედეგად ყალიბდება პოპულაცია, რომელიც მხოლოდ შხამქიმიკატისადმი რეზისტენტული ფორმებისაგან შედგება. ბუნებრივი გადარჩევის გზით მწერების პოპულაციაში მიმდინარეობს ისეთი ალელების ფიქსაცია, რომელიც შეგუების ფართო სპექტრს აპირობებს. გენების მოქმედებით, ერთი მხრივ, ყალიბდება შხამქიმიკატების დამშლელი ეფექტური ფერმენტული სისტემა, მეორე მხრივ, იზრდება მათი შეუღწევადობა ორგანიზმში, ასევე უყალიბდებათ რთული ქცევები, რითაც თავს არიდებენ შხამიან გარემოს. მიკროორგანიზმებში გამოვლენილია სამკურნალო პრეპარატებისადმი (ანტიბიოტიკები, სულფამინამიდური წამლები და მისთ.) მდგრადი შტამები. ჩატარებული გენეტიკური ანალიზით გაირკვა, რომ მუტანტური გენები ლოკალიზებულია R-პლაზმიდაში.
სანიმუშო მაგალითია ნამგლისებრუჯრედიანი ანემიის გავრცელება ადამიანის ზოგიერთ პოპულაციაში. რეცესიული ჰომოზიგოტი (HBS/HBS) ინდივიდები ადრეულ ასაკში (იშვიათად ცოცხლობენ ორ წლამდე) იღუპებიან. ამ გენოტიპის შეგუებულობა თითქმის ნულის ტოლია. მიუხედავად ამისა, ლეტალი (HBS) ალელის სიხშირე ზოგიერთ პოპულაციაში საკმაოდ მაღალია, კერძოდ იმ რაიონებში, სადაც დაავადება მალარიას იწვევს პარაზიტი Plasmodium falciparium-ია. ალისონის გამოკვლევებით აფრიკის აბორიგენების 20%25, ინდოეთის ზოგიერთ რეგიონში მოსახლეობის 10-15%25 ჰეტეროზიგოტებია (HBA/HBB). პოპულაციაში ლეტალური ალელის მაღალი სიხშირით ფიქსაცია იმითაა გამოწვეული, რომ ჰეტეროზიგოტები გაცილებით რეზისტენტულებია მალარიის დაავადებისადმი, ვიდრე ნორმალური ჰომოზიგოტები (HBA/HBA). იმ რეგიონში, სადაც მალარიაა გავრცელებული, ჰეტეროზიგოტებს აღენიშნებათ სელექტიური უპირატესობა ჰომოზიგოტებთან შედარებით: HBB/HBB გენოტიპის ინდივიდები ანემიით, ხოლო HBA/HBA გენოტიპის ინდივიდები მალარიით იღუპებიან. ბუნებრივი გადარჩევის ზეწოლის შედეგად ხდება ჰომოზიგოტების ნაწილობრივი ან მთლიანი ელიმინაცია.
ნამგლისებრუჯრედიანი ანემიის გეოგრაფიული გავრცელება კიდევ ერთხელ ადასტურებს, რომ გენოტიპთა შეგუებადობა გარემო ფაქტორებზეა დამოკიდებული. იმ რეგიონებში, სადაც მალარია არ გვხვდება ან ლიკვიდირებულია, დომინანტი ჰომოზიგოტებისა და ჰეტეროზიგოტების შეგუებადობა ერთნაირია. ამასთანავე, იცვლება ბუნებრივი გადარჩევის მიმართულება. იგი ხელს აღარ უწყობს ჰეტეროზიგოტების შენარჩუნებას პოპულაციაში, რეცესიული ჰომოზიგოტების (HBS/HBS) წინააღმდეგაა მიმართული და მის ელიმინაციას იწვევს, რის შედეგადაც ლეტალი გენის კონცენტრაცია გენოფონდში თანდათანობით ქვეითდება. აშშ-სა და ბრაზილიის ზანგ მოსახლეობაში ჰეტეროზიგოტ ინდივიდთა რაოდენობა გაცილებით ნაკლებია (9-10%25), ვიდრე იმ აფრიკელებში, საიდანაც ისინი იღებენ სათავეს.
მიკროევოლუციის პროცესი შესწავლილია ჩრდილო ამერიკის ერის ტბის სანაპიროზე და კუნძულებზე გავრცელებულ წყლის ანკარას (erodia sipedon) პოპულაციებში (სურ. 8.3). შეფერილობის მიხედვით ანკარები ოთხ ჯგუფად: A,B,C,D იყოფა. A ჯგუფი რუხი შეფერილობის თითქმის უზოლოა, D ჯგუფი მუქი, კარგად გამოხატული ზოლებით, ხოლო B და C ჯგუფის ცხოველებს შუალედი ადგილი უჭირავს ამ ორ კიდურა ჯგუფებს შორის. კუნძულებზე მხოლოდ A,B და C ჯგუფი გვხდება, მაშინ როდესაც ტბის სანაპიროზე (ძირითადად აკრავს ჭაობები) შენიშნულია D ჯგუფის ინდივიდები. კუნძულთა კლდეები, ხევები და სანაპირო ზოლი კირქვებისაგან შედგება (ფონი მოთეთროა). შეისწავლეს კუნძულზე დაჭერილი განაყოფიერებული მდედრებისაგან მიღებული შთამომავლობა. გამოჩეკილ ინდივიდებში ოთხივე სახის შეფერილობა გვხვდება. ანალოგიური მოვლენა შეინიშნება კუნძულზე ახლად გამოჩეკილ ინდივიდებში. ზრდასრული ინდივიდები კი მხოლოდ A და B ჯგუფებისაგან შედგება. ამ მოვლენას პოსტემბრიონულ პერიოდში ინდივიდთა ამორჩევითი ელიმინაცია იწვევს. რუხი შეფერილობა მფარველობითია. კუნძულებზე ფრინველები ადვილად არჩევენ მუქ ფორმებს და ანადგურებენ. საპირისპირო მოვლენას ვხვდებით ტბის ნაპირზე გავრცელებულ ცხოველებში, სადაც დამცველობით როლს ზოლიანი შეფერილობა ასრულებს.
სურ. 8.3. ერის ტბის კუნძულზე გავრცელებულ ანკარებში მიმდინარე ბუნებრივი გადარჩევა (სქემა) (ა. პარამონოვი, 1978).
კონგისა და ლოუსენის (1995) მიერ დადგენილია, რომ ზოლიანობას განსაზღვრავს ერთი ვარიაბილური დომინანტური გენი. რეცესიული უზოლობის განმსაზღვრელი ალელის სიხშირე კუნძულებზე გავრცელებულ პოპულაციებში საშუალოდ 0,73-ის ტოლია. ხელოვნურ პირობებში გამრავლებულ და კუნძულებზე გაშვებულ ინდივიდებში გამოთვლილია გადარჩევის კოეფიციენტი, რომელიც სხვადასხვა კუნძულზე განსხვავებულია და ის S=0,11-დან S=0,28-მდე ვარირებს. მიუხედავად ბუნებრივი გადარჩევის ძლიერი ზეწოლისა, კუნძულებზე მაინც გვხვდება ზოლიანი ფორმები. ეს, ძირითადად, გენთა ნაკადითაა გამოწვეული, რომელიც, დაახლოებით, 10%25 -ს შეადგენს.
|
|
10.2.4 8.7. ბუნებრივი გადარჩევის ეფექტურობა და სიჩქარე |
▲ზევით დაბრუნება |
გადარჩევის, როგორც პოპულაციის გენეტიკურ დინამიკაზე მოქმედი ფაქტორის მოქმედების კანონზომიერებათა გასაგებად აუცილებელია ელემენტარული ევოლუციური მოვლენების შესწავლა, კერძოდ გადარჩევის მოქმედების ეფექტურობის შეფასება მონოგენურ პანმიქტურ პოპულაციაში გენური სიხშირეების ცვლილების მიხედვით. გადარჩევის მოქმედების ეფექტურობა ე.ი. გენოფონდის გარდაქმნის სიჩქარე (რომელიც შეიცავს ადაპტურ ალელებს) დამოკიდებულია მრავალ პარამეტრზე, რომელთაგან უნდა გამოიყოს რამდენიმე.
გენოტიპურად განსხვავებული კლასების ადაპტური ღირებულების გადამწყვეტ კრიტერიუმს წარმოადგენს მათ მიერ ყოველ თაობაში წარმოქმნილ შთამომავალთა შეფარდებითი რაოდენობა, რომელთაც ნორმალური ცხოველმყოფელობა ახასიათებთ. გენოტიპის უნარი გადარჩენისა და პოპულაციებში შთამომავლობის კვლავ წარმოქმნისა სხვა გენოტიპთან შედარებით გენოტიპის ადაპტურ ღირებულებას წარმოადგენს. იგი გენოტიპის შეფარდებითი შეგუებულობის ხარისხის მახასიათებელია და აღინიშნება ასო W-თი, გენოტიპის ადაპტური ღირებულება მერყეობს 0-დან 1-მდე. როცა W=0 ინდივიდის მიერ გენეტიკური ინფორმაციის გადაცემა შემდეგ თაობაზე არ ხდება, ხოლო W=1 დროს მოცემული - მემკვიდრული ნიშნით გამეტათა მაქსიმალური რაოდენობის წარმოქმნა ხდება.
გენოტიპის ადაპტური ღირებულება განისაზღვრება გენთა მთელი კომპლექსით. ამდენად გენოტიპები რომლებიც ჰომოზიგოტურები არიან ერთი და იმავე გენის მიხედვით, შეიძლება განსხვავებული ადაპტური ღირებულებით ხასიათდებოდნენ. ადაპტური ღირებულების მთავარ მახასიათებლებს წარმოადგენენ გადარჩენადობა და გამრავლებადობის - კვლავწარმოქმნის უნარი, გადარჩენადობა განისაზღვრება განვითარების სიჩქარით, სხვადასხვა დაავადების მიმართ გამძლეობით, სხვადასხვა არახელსაყრელი ფაქტორების მოქმედების თავიდან აცილებით, სასიცოცხლო აქტივობით და სხვა. გამრავლებადობის უნარი - შთამომავლობის წარმოქმნის უნარი დამოკიდებულია სქესობრივი მომწიფების სისწრაფეზე, სქესობრივი ქცევის თავისებურებებზე, სქესობრივ აქტივობაზე, ფერტილობაზე, სახეობრივი ადაპტაციების განვითარების ხარისხზე, შთამომავლობის ზრუნვაზე და სხვა. ყოველი ეს თავისებურება კონტროლირდება გენოტიპით, მაგრამ მათი რეალიზაცია ონტოგენეზში განპირობებულია შესაძლებლობების ფარგლებში - რეაქციის ნორმის ფარგლებში.
ადაპტურ ღირებულებას განსაზღვრავენ ალელთა სიხშირის აღრიცხვით გადარჩევამდე და გადარჩევის შემდეგ, ე.ი. მათი სიხშირის აღრიცხვით თაობებში, რაც თავის მხრივ გადარჩევის კოეფიციენტზეა დამოკიდებული.
გადარჩევის კოეფიციენტი ახასიათებს ელიმინაციის ინტენსივობას ან საწყის ფორმასთან შედარებით მუტანტური ალელის წარმოქმნის დაქვეითებას. იგი აღინიშნება ასოთი S და წარმოადგენს გენოტიპის ადაპტური ღირებულების საწინააღმდეგო სიდიდეს. თუ W=1 მაშინ S=0, თუ 100 გაჩენილი ინდივიდიდან გარკვეული ნიშნით გამრავლებამდე ყველა იცოცხლებს, ხოლო სხვა ნიშნით - მხოლოდ 99 ინდივიდი, მაშინ შეიძლება ითქვას, რომ პირველ შემთხვევაში გადარჩევის კოეფიციენტი 0-ია; ხოლო მეორე შემთხვევაში - 0,01, ბუნებრივ პირობებში კოეფიციენტი ჩვეულებრივ არ აღემატება 0,10 - 0,20, ხშირად და ამიტომ ნაკლები მნიშვნელობა აქვს. რასაკვირველია ლეტალური მუტაციების შემთხვევაში გადარჩევის კოეფიციენტი შეიძლება იყოს 1,0.
ბუნებაში გენოტიპის ადაპტური ღირებულების შეფასება ცალკეული ნიშნების მიხედვით იშვიათად ხდება, ჩვეულებრივ გადარჩევა წარმოებს ნიშანთა კომპლექსის მიხედვით, პოპულაციაში ყველა გენოტიპის ადაპტური ღირებულების დასახასიათებლად განსაზღვრავენ საშუალო შეგუებადობას, „W”. იგი ტოლია გენოტიპის სიხშირის ჯამის გადარჩევის შემდეგ და განისაზღვრება ჰარდი-ვაინბერგის ფორმულით:
W=P2WO+2PQW1+Q2W2
პირველად ღებულობენ დომინანტური ჰომოზიგოტის საშუალო შეგუებადობას, ხოლო სხვა გენოტიპების შეგუებადობას მასთან აფარდებენ.
AA-W0=W0/W0=1
AA - W1=W1/W0=1-S1
AA-W2=W2/W0=1-S2
ამ ფორმულის გამოყენებით და გენოტიპების შედარებით გადარჩევამდე და გადარჩევის შემდეგ, შეიძლება გადარჩევის კოეფიციენტის განსაზღვრა (იხ. ცხრ. 8.2).
რაც უფრო მაღალი მნიშვნელობისაა გადარჩევის კოეფიციენტი, მით უფრო ინტენსიურია გადარჩევა რომელიმე გენოტიპის მიმართ, ე.ი. გადარჩევის ზეწოლა მაღალია, გადარჩევის ზეწოლა გარკვეულ მონაკვეთში განსაზღვრული დადებით შედეგს იძლევა (გენური სიხშირეების ცვლილება), ეს არის გადარჩევის ეფექტურობა და ის კოეფიციენტზეა დამოკიდებული.
ცხრილი 8.2.
გენოტიპის გადარჩევის კოეფიციენტის განსაზღვრა
მაჩვენებლები |
გენოტიპები |
||
|
|
A |
AA |
სიხშირე |
0,25 |
0,50 |
0,25 |
სიხშირე |
0,35 |
0,48 |
0,17 |
შეფარდეით |
W0=0,35/0,25=
W0/W0= |
W1=0,48/0,50=
W1/W0= |
W2=0,17/0,25=
W2/W0=0,68/ |
გადარჩევის |
1,0-1,0=0 |
1,0-0,7=0,3 |
1,0-0,4=0,6 |
ბუნებრივი გადარჩევის სისწრაფე ამა თუ იმ ალელის მიხედვით დამოკიდებულია ალელის სელექციურ უპირატესობაზე. ალელის კონცენტრაციის ცვლილების სისწრაფე გადარჩევის კოეფიციენტის პროპორციულია. გადარჩევის სისწრაფე განისაზღვრება, აგრეთვე, მოცემულ თაობაში ალელის კონცენტრაციით. ამიტომ პოპულაციის გარდაქმნის სისწრაფე თაობიდან თაობამდე კანონზომიერად იცვლება.
ბუნებრივი გადარჩევის მოქმედების არეში ექცევა არა კონკრეტული ალელით განსაზღვრული ფენი, არამედ მათი ერთობლიობა - ფენოტიპი (ბუნებრივია გენოტიპი). ეს უკანასკნელი პოპულაციასთან ერთად ევოლუირებს. ინდივიდის ფენოტიპის ევოლუცია არის იმავდროულად პოპულაციის ევოლუირების შედეგი. ამის გამო აუცილებელია ბუნებრივი გადარჩევის მოქმედების შედეგად პოპულაციის გენოფონდის გარდაქმნის პროცესის განხილვა. შედარებით მუდმივ ერთნაირ პირობებში, რაც უფრო დიდია კონკრეტული ალელის სელექციური უპირატესობა, მით უფრო სწრაფად მიმდინარეობს გადარჩევის პროცესი. სელექციური უპირატესობა გადარჩევის კოეფიციენტით გამოიხატება.
S=P1-P/P-PP1
სადაც P - არის ალელის სიხშირე პირველ თაობაში, P1 - კი ალელის სიხშირე მეორე თაობაში. როგორც ზემოთ აღვნიშნეთ თეორიულად გადარჩევის კოეფიციენტი 0-დან 1-ის ფარგლებში ვარირებს. ალელის კონცენტრაციის ცვლილება (W0/W0၄P) არის გადარჩევის კოეფიციენტის პროპორციული. როდესაც შ-ის მნიშვნელობა მცირეა, მაშინ იყენებენ შემდეგ ფორმულას: ၄P=S(1-P)P. ზოგადი ფორმულა კი შემდეგია
၄P=P(1-P)S/1PS
ითვლება, რომ ბუნებაში გადარჩევის კოეფიციენტი დაახლოებით 0,001-0,01-ია. გადარჩევის ასეთი კოეფიციენტისას ევოლუცია ძალიან ნელა მიმდინარეობს. ჯ. ჰოლდეინის გამოთვლით S=0,01, დომინანტური ალელის კონცენტრაციის ასამაღლებლად P=0,0000001-დან P=0,000002-მდე აუცილებელია 11739 თაობა, ხოლო რეცესიული ალელისათვის 321 444 თაობა. აქედან გამომდინარეობს კიდევ ერთი დასკვნა, რომ პოპულაციის გარდაქმნა სხვა თანაბარ პირობებში მით უფრო სწრაფად ხდება, რაც უფრო სწრაფია თაობათა ცვლა.
და ბოლოს, პოპულაციის გარდაქმნის სისწრაფე დამოკიდებულია პოპულაციის სიდიდეზე. რაც უფრო მცირერიცხოვანია პოპულაცია, მით უფრო სწრაფად შეიცვლება იგი მოცემული ალელის მიხედვით, მაგრამ მცირე პოპულაციაში როგორც ვიცით გადარჩევას ხელს უშლის გენთა დრეიფი. გენთა დრეიფს შეუძლია გადარჩევის გარეშე პოპულაციის გენოფონდიდან ამა თუ იმ ალელის გაქრობა ან კონცენტრაციის გაზრდა და ნორმად ქცევა, მაგრამ როგორც კი ალელი ფენოტიპურად ვლინდება, მასზე უკვე მოქმედებს გადარჩევა, ამიტომ გენთა დრეიფი უპირატესად რეცესიულ ალელებსა და ნეიტრალურ ნიშნებზე მოქმედებს და შესაბამისად მნიშვნელოვან გავლენას არ ახდენს ადაპტურ ევოლუციაზე. მას შეუძლია იმოქმედოს ევოლუციის მასალაზე და არა მის მიმართულებაზე.
მცირე პოპულაციებში გარდაქმნის სიჩქარე დამოკიდებულია გარკვეული ხარისხის მუტირების სიჩქარეზე. დიდ პოპულაციებში სიტუაცია შებრუნებულია. მათში იზრდება თაობათა რიცხვი, რომლებიც საჭიროა იმისათვის, რომ მოცემული ალელი პოპულაციის ყველა ინდივიდში გავრცელდეს. გადარჩევა მეტად ეფექტურია იმ შემთხვევაში, როცა სახეობა არსებობს არა ძალიან მსხვილი ნაკლებად იზოლირებული, პოპულაციების სისტემის სახით. ასეთი სტრუქტურის არსებობისას ყოველი პოპულაციის შეფარდებით მცირე ზომები ადაპტური ალელების ჰომოზიგოტიზაციას აიოლებენ, ხოლო ინდივიდთა მიგრაცია უზრუნველყოფს ყოველ პოპულაციაში ახალი ალელების ნაკადის უზრუნველყოფას, ე.ი. ამაღლებს მის გენეტიკურ ცვალებადობას.
სახეობის სტრუქტურის განხილვისას ამ პოზიციიდან ნათელი ხდება, რომ სახეობის შიდა ჯგუფების რთული იერარქიული სისტემა ოპტიმალურია ევოლუციისათვის, რადგან იგი უზრუნველყოფს ახალი ალელების სწრაფ გამოვლენას ნათესაურ ჯგუფებში ინბრიდინგის შედეგად, ადაპტური ვარიანტების სწრაფ გავრცელებას ოჯახური და ჯგუფური ელიმინაციის ხარჯზე და ცვალებადობის მაღალ დონეს ჯგუფებს შორის გენთა გაცვლის გამო.
ამრიგად, პოპულაციის გენოფონდი გადარჩევის გავლენით იცვლება არაადაპტური ვარიანტების ელიმინაციისა და ადაპტური ვარიანტების გავრცელების ხარჯზე. გადარჩევის ტემპზე მოქმედებენ შემდეგი ფაქტორები: 1) გადარჩევის კოეფიციენტი, ე.ი. კონკრეტული ალელის სელექციური უპირატესობა, 2) მოცემულ თაობაში მოცემული ალელის კონცენტრაცია და მისი დომინანტობის ხარისხი. 3) თაობათა ცვლის სიხშირე 4) რიცხოვნობის მერყეობის ხარისხი 5) პოპულაციის ზომა, მაგრამ ნებისმიერი პოპულაციის ევოლუციის რეალური ტემპი, პირველ რიგში, განისაზღვრება გარემოს ცვალებადობის სიჩქარით. გარემოს ცვლილების ტემპზე ევოლუციის ტემპების ჩამორჩენისას პოპულაცია იღუპება: ამრიგად, გადარჩევის კოეფიციენტი დამოკიდებულია არსებობისათვის ბრძოლის სიმძლავრეზე.
ბუნებრივი გადარჩევა როგორც უკვე აღვნიშნეთ წარმოადგენს პოპულაციაში ინდივიდთა ნაწილის დიფერენციალური გადარჩენისა და შემდგომ თაობებში გადარჩენილ ინდივიდთა გენოტიპის დიფერენციალური კვლავწარმოქმნის პროცესს. გენოტიპების დიფერენციალური კვლავწარმოქმნა პოპულაციაში ადაპტური ნიშნების გავრცელებას იწვევს, ამ გზით ხდება პოპულაციის გენოფონდის ევოლუციური ცვლილება. ეუკარიოტებში ადაპტური გენური კომპლექსები წარმოიქმნება არა უშუალოდ მუტაციური პროცესის შედეგად, არამედ გენომთა კომბინაციების გადარჩევის შედეგად. გადარჩევა ხდება ფენოტიპური ნიშნების მიხედვით, მაგრამ გადაირჩევიან ერთიანი გენოტიპები, ამის გამო ევოლუირებს ონტოგენეზი.
აუცილებელია ხაზი გაესვას იმას, რომ ბუნებრივი გადარჩევა მიმართული, ვექტორიზებული პროცესია და როგორც ყოველ ვექტორს სამი პარამეტრი გააჩნია: 1. სათავე-ნიშანი, რომლის მიხედვითაც მიმდინარეობს გადარჩევა; 2. სიდიდე, რომელიც გადარჩევის კოეფიციენტით გამოიხატება; 3. მიმართულება, მისი განმსაზღვრელი არის არსებობისათვის ბრძოლა.
|
|
10.2.5 8.8. ბუნებრივი გადარჩევის ფორმები |
▲ზევით დაბრუნება |
თანამედროვე ევოლუციურ თეორიაში საკვანძოა საკითხი ბუნებრივი გადარჩევის ფორმების შესახებ. მკვლევართა მიერ გადარჩევის 30-ზე მეტი ფორმაა გამოვლენილი და გაანალიზებული. თითოეული მათგანი ზოგადი, ერთიანი ბუნებრივი გადარჩევის მოქმედების სპეციფიკურ, კერძო მხარეს წარმოადგენს. პოპულაციურ დონეზე გადარჩევის სამ ძირითად ფორმას გამოყოფენ: მასტაბილიზებელს, მამოძრავებელსა და დიზრუპტულს. მასტაბილიზებელი და მამოძრავებელი გადარჩევის ფორმა პირველად ამერიკელმა მეცნიერმა რ. ფიშერმა (1930) გამოყო და მისი მოქმედების მექანიზმი მათემატიკურად დაასაბუთა. გადარჩევის დიზრუპტული ფორმა გამოყო და მისი მოქმედების მექანიზმი თეორიულად დაასაბუთა ჯ.სიმპსონმა (1944). ტერმინები „მასტაბილიზებელი და მამოძრავებელი“ გადარჩევა შემოიტანა ს.შმალგაუზენმა (1946).
|
|
10.2.5.1 მასტაბილიზებელი გადარჩევა |
▲ზევით დაბრუნება |
მასტაბილიზებელი გადარჩევა არის ბუნებრივი გადარჩევის ისეთი ფორმა, რომელიც მიმართულია პოპულაციაში ნიშნის საშუალო, ადრე ჩამოყალიბებული მნიშვნელობის შენარჩუნებისა და მდგრადობის ამაღლების რეალიზაციისაკენ. იგი მიმდინარეობს ნორმიდან ნებისმიერი გადახრის ელიმინაციის გზით (სურ. 8.4).
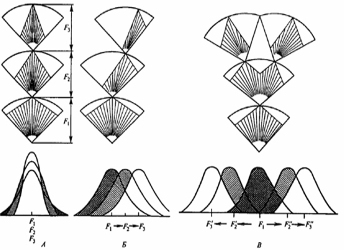
სურ. 8.4. ბუნებრივი გადარჩევის სამი ძირითადი ფორმა: მასტაბილიზებელი (A), მამოძრავებელი (B) და დიზრუპტული (C). მოცემულია გადარჩევის მოქმედების სქემა (ნ. ტიმოფეევ-რესოფსკი და სხვ., 1977).
მასტაბილიზებელი გადარჩევის მოქმედებისას უპირატესად მრავლდებიან და შთამომავლობას ტოვებენ ის ინდივიდები, რომელთაც პოპულაციისათვის დამახასიათებელი ნიშნის საშუალო მაჩვენებელი აქვთ. ინდივიდები, რომელთაც საშუალო ნორმიდან ნებისმიერი მიმართულებით ფენოტიპური გადახრა აქვთ, ელიმინირდებიან. მასტაბილიზებელი გადარჩევა ინახავს და აძლიერებს ადრე ჩამოყალიბებულ ადაპტაციურ ნიშნებს. ამასთანავე, იგი ასტაბილიზებს ორგანიზმების ურთიერთობას გარემოსთან. მასტაბილიზებელი გადარჩევა მოქმედებს მეტ-ნაკლებად უცვლელ გარემო პირობებში (ე.ი. როდესაც გარემო პირობები საშუალო მაჩვენებლის ფარგლებში ფლუქტუირებს). ბუნებრივია, ამ პირობებში მოქმედებას აგრძელებს მუტაციური პროცესი, არსებობისათვის ბრძოლა და ბუნებრივი გადარჩევა, მაგრამ ინდივიდებში ადრე ჩამოყალიბებული ადაპტაციური ნიშნები არ იცვლება. წარმოქმნილი მუტაციები მავნეა, ვინაიდან არღვევენ გარემოსთან იმ შეგუებულობას, რომელიც ჩამოყალიბდა პოპულაციის ევოლუციის პროცესში, ამიტომაც ახალი ადაპტაციები აღარ წარმოიქმნება. ინახება მხოლოდ უკვე მიღწეული შეგუებითი ნორმა. ამრიგად, მასტაბილიზებელი გადარჩევის ყველაზე თვალსაჩინო შედეგია უკვე არსებული ადაპტაციური ნიშნებისა და მათთვის დამახასიათებელი რეაქციის ნორმის შენარჩუნება და სტაბილიზაცია.
მასტაბილიზებელი გადარჩევის მოქმედების საილუსტრაციოდ მოვიყვანთ რამდენიმე მაგალითს. ამერიკელმა ორნითოლოგმა გ. ბამპერსმა 1898 წლის ზამთარში მანჰეტენის ქუჩებში (ნიუ-იორკის უბანი) შეაგროვა ძლიერი ქარბუქისა და ყინვისაგან გათოშილი და ცოცხალმკვდარი 136 სახლის ბეღურა (asser domesticus) ლაბორატორიაში ისინი გაათბეს და ჩაუტარეს ბიომეტრიული ანალიზი. გადარჩნენ ის ბეღურები (მხოლოდ 72), რომელთა ნიშნები პოპულაციის საშუალო მაჩვენებლებს უახლოვდებოდა. ნებისმიერი ნიშნის მიხედვით (ფრთის, ნისკარტის და სხეულის სიგრძე, სხეულის მასა და სხვა) ნორმიდან გადახრილი ფრინველები დაიღუპნენ.
მ. კარნომ და ლ.ჰენრუზმა (1952) ადამიანში გამოავლინეს კავშირი ახალშობილის წონასა და მათ სიკვდილიანობას შორის. რაც უფრო მეტი იყო გადახრა საშუალო მაჩვენებლიდან 3,6კგ (როგორც კლებადობით, ისე მატებით) მით უფრო იზრდებოდა სიკვდილიანობა. გადარჩევა „იღვწის“ საშუალო მაჩვენებლის შენარჩუნებისთვის.
ენტემოფილურ მცენარეებში ყვავილის ფორმა და ზომა შედარებით უფრო სტაბილურია, ვიდრე ანემოფილურებში. ყვავილისა და მათი დამტვერავი მწერების ზომები ერთიმეორეს მიესადაგება. ყვავილის აგებულების მდგრადობა მასტაბილიზებელი გადარჩევით შენარჩუნდება. მდგრადობა მრავალი მილიონი წლის მანძილზე მიმდინარე ენტემოფილური მცენარეებისა და მათი დამტვერავი მწერების ერთობლივი (შეუღლებული) ევოლუციითაა განპირობებული. ხდებოდა შეცვლილი ყვავილის მქონე მცენარეების „წუნდება“. ვინაიდან მწერები მათ ვერ მტვერავდნენ, თესლს მხოლოდ „სტანდარტული“ ყვავილის მქონე მცენარეები იკეთებენ.
ცოცხალი სისტემის ორგანიზაციის ნებისმიერ დონეზე ნიშნების შენარჩუნება მასტაბილიზებელი გადარჩევის შედეგია. ორგანიზაციის მოლეკულურ დონეზე მისი მოქმედებით იქმნება სიცოცხლის ბიოქიმიური ერთიანობა და უნივერსალობა, კერძოდ გენეტიკური კოდის უნივერსალობა (ვირუსებში, პროკარიოტებსა და ეუკარიოტებში ერთნაირი გენეტიკური კოდი ფუნქცირებს), ცილის მსგავსი ამინომჟავური შედგენილობა (შენებაში 20 სახის ამინომჟავა მონაწილეობს), ხერხემლიან ცხოველებში ჰემის მოლეკულის ერთნაირი აგებულება და სხვა. ორგანიზაციის უჯრედულ დონეზე მასტაბილიზებელი გადარჩევის მოქმედებით შენარჩუნებულია ორგანოიდების ერთნაირი სტრუქტურა და ფუნქცია (ეუკარიოტებში), მიტოზური და მეიოზური გაყოფის ერთნაირი მექანიზმი და სხვ. სახეობის დონეზე კი პერსისტენტული ფორმების არსებობა. მასტაბილიზებელი გადარჩევის ხანგრძლივი მოქმედების შედეგად ზოგიერთი სახეობა (ე.წ. პერსისტენტული ფორმები) მრავალი ათასი და მილიონი თაობის განმავლობაში თითქმის უცვლელი დარჩა. ამგვარი „ცოცხალი ნამარხი“ სახეობებია: მოლუსკი neopilina eopilina galatheae (დაახლოებით 600 მილიონი წელია, რაც არსებობს), მოლუსკი nautilusi autilus pompilis (დაახლოებით 500 მილიონი წელი), მახვილკუდიანები iphosura (დაახლოებით 400 მილიონი წელია რაც არსებობს), tuataria (hateria) Sphenodon punctatus (დაახლოებით 230 მილიონი წელია რაც არსებობს), gingko ingkgo biloba (დაახლოებით 240 მილიონი წელია რაც არსებობს), ციანობაქტერიის ზოგიერთი სახეობა (არანაკლებ 1,5-2 მილიარდი წელია, რაც არსებობს) და სხვა (სურ. 11.3).
|
|
10.2.5.2 მამოძრავებელი გადარჩევა |
▲ზევით დაბრუნება |
მამოძრავებელი გადარჩევა ბუნებრივი გადარჩევის ისეთი ფორმაა, რომელიც ნიშნის ან თვისების საშუალო მნიშვნელობის ცვლილებას განსაზღვრული მიმართულებით წარმართავს. ზოგჯერ მას დარვინულ გადარჩევასაც უწოდებენ. იგი მასტაბილიზებელი გადარჩევის ალტერნატიული ფორმაა, რომელიც გარემოს შეცვლისას ვლინდება და ახალი ადაპტაციური ნიშნების ჩამოყალიბებას იწვევს. მამოძრავებელი გადარჩევის მეშვეობით ახალ გარემოში გამოუსადეგარი ძველი ნორმა ახლით იცვლება და მისი განმტკიცება ხდება. ამა თუ იმ ნიშნის გამოვლენის ხარისხი შეიძლება შესუსტდეს ან გაძლიერდეს. მამოძრავებელი გადარჩევის დროს ერთვება ევოლუციის პროცესში შეცვლილი გარემო პირობების შესაბამისი მემკვიდრული ცვლილებები, ე.წ. სამობილიზაციო რეზერვი. მუტაციები, რომელიც ადრე საზიანო ან ნეიტრალური იყო, ახლა შეიძლება სასარგებლო აღმოჩნდეს. ის ინდივიდები, რომელთაც ახალ გარემოში უპირატესობა არა აქვთ, ელიმინირდება. პოპულაციაში თანდათანობით ვრცელდება სასარგებლო გადახრა და ყალიბდება ახალ-ახალი ადაპტაციები (სურ. 8.4).
სურ. 8.5. სხვადასხვა წლებში ინგლისში ვარფარინისადმი მდგრადი რუხი ვირთაგვას (Rattus norvegicus) პოპულაციის გამოვლენილი კერები (დ. დრუმონდი, 1970).
ევოლუციური თეორიის განვითარების საწყის ეტაპზე მოწოდებული ბუნებრივი გადარჩევის დამამტკიცებელი პირდაპირი და არაპირდაპირი არგუმენტების დიდი ნაწილი გადარჩევის ამ ფორმას მიეკუთვნება და დარვინისეული, კლასიკური ბუნებრივი გადარჩევის არსის გამომხატველია.
მამოძრავებელი გადარჩევის საილუსტრაციოდ რამდენიმე მაგალითს მოვიყვანთ. კლასიკურადაა აღიარებული ინგლისელი ზოოლოგის ვ.უელდონის (1877) მიერ ჯერ კიდევ ჩ.დარვინის სიცოცხლეში კიბორჩხალაზე (Carcinus maenas) ჩატარებული გამოკვლევები. ხუთი წლის განმავლობაში იგი აკვირდებოდა ცხოველებს. ქ. პლიმუტის (ინგლისი) ნავსადგურში ახალი ჯებირის აშენებამ ფართო სალაყუჩე ნაპრალის მქონე ცხოველების ელიმინაცია გამოიწვია. გადარჩნენ ვიწრო სალაყუჩე ნაპრალის მქონე კიბორჩხალები. გამოირკვა, რომ ჯებირმა წყლის ცირკულაციის დარღვევა გამოიწვია. ორი მდინარის მიერ ჩამოტანილი მინარევებით წყალი იმღვრეოდა. წყალში შეტივნარებული კირქვის მინარევები ილექებოდა ფართო ნაპრალიანი კიბორჩხალების ლაყუჩებში და ისინი იღუპებოდნენ. დასკვნა დადასტურებულ იქნა ჩატარებული ექსპერიმენტით: აკვარიუმში მოათავსეს ორივე ფორმის 248 კიბორჩხალა და განუწყვეტლივ ამღვრევდნენ წყალს. სწრაფად დაიღუპა 154 ცხოველი, ყველა მათგანს ფართო სალაყუჩე ნაპრალი აღმოაჩნდათ.
1947 წელს ინგლისში ვირთაგვების წინააღმდეგ გამოიყენეს ანტიკოაგულანტი-ვარფარინი. სატყუარათი ნივთიერების მიღების შემდეგ ცხოველები ჩხუბში ან სხვა გზით წარმოქმნილი ნაკაწრებიდან სისხლდენის შედეგად იღუპებოდნენ. 1958 წელს შოტლანდიაში პირველად აღმოაჩინეს - ვარფარინისადმი მედეგი (ე.წ. „სუპერვირთაგვები“) ფორმები. 1960 წ. კი ინგლისის მრავალ მხარეში გამოავლინეს რეზისტენტული პოპულაციები, რომლებიც მთელი ალბიონის კუნძულზე განსახლდნენ. გამოირკვა, რომ რეზისტენტულობას იწვევდა ერთი მუტანტური დომინანტი გენი (სურ. 8.5).
ფუნქციური უვარგისობის შემთხვევაში მამოძრავებელი გადარჩევა ნიშნის დაკარგვას იწვევს. ზოგიერთ მწერებსა (ოკეანიის კუნძულებზე) და ფრინველებში (მაგ., კივი, სირაქლემა, პინგვინი) ფრთების, გველებში - კიდურების, ჩლიქოსნებში - თითების, მღვიმის ცხოველებში - თვალების, პარაზიტ მცენარეებში - ფოთლებისა და ფესვების დაკარგვა მამოძრავებელი გადარჩევით გამოწვეული ორგანოთა რედუქციის ნათელი მაგალითებია.
ბუნებაში ვიწროქალა მემინდვრიაზე (Microtus gregalis) დაკვირვებით დადასტურებულია მამოძრავებელი გადარჩევის მოქმედება (შვარცი, 1966). ვიწროქალა მემინდვრია დიდი ხანი არ არის, რაც პოლარულ ურალში განსახლდა და ქვესახეობად ჩამოყალიბდა. იგი სამხრეთში გავრცელებული ქვესახეობისაგან მორფოფიზიოლოგიურად განსხვავდება. კერძოდ, მას აქვს დიდი ზომის სხეული (რითაც სხეულის ტემპერატურის გაცემა დაქვეითებულია), ღვიძლი და გული (ენერგეტიკული ცვლა გაზრდილი აქვს), ემბრიონების რიცხვი გაზრდილია.
ბუნებაში გადარჩევის ორივე (მასტაბილიზებელი და მამოძრავებელი) ფორმა მუდმივად მოქმედებს. შესაძლებელია დროის ერთ მონაკვეთში ერთი უფრო მეტად მოქმედებდეს, ვიდრე მეორე. მასტაბილიზებელი გადარჩევა კონსერვატორულ როლს ასრულებს, ხოლო მამოძრავებელი კი - პროგრესულს. ისინი კომპლემენტურ მოქმედებას ავლენენ. მასტაბილიზებელი ინახავს და ამაგრებს ევოლუციურ გარდაქმნებს, რომელიც მამოძრავებელი გადარჩევის შედეგად მიიღება.
|
|
10.2.5.3 დიზრუპტული გადარჩევა |
▲ზევით დაბრუნება |
დიზრუპტული გადარჩევა ბუნებრივი გადარჩევის ისეთი ფორმაა, რომელიც იწვევს თვისებრივად განსხვავებული რამდენიმე ფორმის (ფენოტიპური კლასის) არსებობას და მოქმედებს შუალედური ნიშნის (საშუალო მნიშვნელობის) საწინააღმდეგოდ. დიზრუპტული გადარჩევის თეორია დაამუშავა ჯ.სიმპსონმა (1944). მას ხშირად დამანაწევრებელ გადარჩევასაც უწოდებენ. გადარჩევის ეს ფორმა გაცილებით იშვიათად გვხვდება, მაგრამ ევოლუციურ გარდაქმნაში მნიშვნელოვანი როლის შესრულება შეუძლია. იგი მოქმედებს მაშინ, როცა ერთ ტერიტორიაზე გარკვეულ საარსებო პირობებში არსებობისათვის ბრძოლაში გენოტიპთა ვერცერთი ჯგუფი ვერ აღწევს აბსოლუტურ უპირატესობას. გარემო პირობების მერყეობა (მაგ., სეზონური ცვლილებები, კლიმატური ფაქტორები) ხელს უწყობს პოპულაციაში რამდენიმე განსხვავებული ფენოტიპის არსებობას. ამ დროს ერთ პირობებში მიმდინარეობს ერთგვარი ნიშნების, სხვა პირობებში - სხვაგვარი ნიშნების გადარჩევა. პოპულაციაში გადარჩევის ზეწოლამ შეიძლება გამოიწვიოს მისთვის დამახასიათებელი საშუალო მაჩვენებლიდან ფენოტიპების გადახრა სხვადასხვა მიმართულებით. შუალედი ფორმების ელიმინაციის შედეგად პოპულაცია რამდენიმე ჯგუფად-სუბპოპულაციად ნაწევრდება. თუ სუბპოპულაციებს შორის გენთა ნაკადი გარკვეული მიზეზით შეწყდება, თითოეულმა შეიძლება დასაბამი მისცეს ახალ სახეობას. რიგ შემთხვევაში გადარჩევის ეს ფორმა იწვევს ერთ პოპულაციაში რამდენიმე განსხვავებული ფენოტიპის წარმოქმნას (ე.ი. პოლიმორფიზმის წარმოშობას). პოლიმორფიზმის ყველა შემთხვევა დიზრუპტული გადარჩევის მოქმედების შედეგია. მისი საბოლოო შედეგი შეიძლება წარმოვიდგინოთ როგორც მამოძრავებელი და მასტაბილიზებელი გადარჩევის ერთდროული და მონაცვლეობითი მოქმედება. დიზრუპტული გადარჩევა მამოძრავებელი გადარჩევის მსგავსად პროგრესულ ფუნქციას ასრულებს.
საილუსტრაციოდ მხოლოდ ერთ კლასიკურ მაგალითს მოვიყვანთ. ბაღის ლოკოკინას ჩეპაეა ნემორალის-ის ინდივიდები ნიჟარის შეფერილობისა და ზოლების რაოდენობით განსხვავდებიან. გვხვდება ყავისფერ, ვარდისფერ და ყვითელნიჟარიანი მოლუსკები. ნიჟარის ნაირგვარი შეფერილობა მფარველობითია, რაც იცავს მოლუსკებს ფრინველებისაგან. ბაღის ლოკოკინას ერთიანი პოპულაცია რამდენიმე ფორმად (მორფა) იყოფა და არცერთ მათგანს არა აქვს უპირატესობა სხვების მიმართ. დიზრუპტული გადარჩევის მოქმედებით ლოკოკინაში რამდენიმე ფენოტიპური კლასია ჩამოყალიბებული (დაწვრილებით იხ. §9.1).
|
|
10.2.5.4 სქესობრივი გადარჩევა |
▲ზევით დაბრუნება |
ბუნებრივი გადარჩევის, იმ ფორმას რომელიც ცალსქესიან ინდივიდებში მხოლოდ ერთი სქესის ინდივიდებს ეხება, სქესობრივი გადარჩევა ეწოდება. სქესობრივ გადარჩევას ყურადღება მიაქცია ჩ.დარვინმა „სახეობათა წარმოშობაში“. იგი აღნიშნავს: „გადარჩევის ეს ფორმა განისაზღვრება არსებობისათვის ბრძოლით არა სხვა ორგანიზმების ან გარემო პირობების წინააღმდეგ, არამედ ერთი და იმავე სქესის ინდივიდთა, უპირატესად მამრებს შორის მეორე სქესის ინდივიდთა დაუფლებისათვის. ამის შედეგად წარუმატებელი მამრი კი არ კვდება, არამედ ხდება მისი შთამომავლობის შეზღუდვა ან სრული შეწყვეტა“.
სქესობრივი გადარჩევა ბუნებრივი გადარჩევის კერძო შემთხვევაა, რომელიც მიმდინარეობს პოპულაციის ერთი სქესის ინდივიდებს შორის გამრავლებისა და შთამომავლობის დატოვებისათვის. ჩ. დარვინი გამოყოფდა სქესობრივი გადარჩევის ორ ფორმას. ერთ შემთხვევაში მდედრი პასიური რჩება და ბრძოლა მიმდინარეობს მამრებს შორის მდედრის დასაუფლებლად. ბრძოლაში გამარჯვებული ძლიერი, ჯანსაღი, კარგად განვითარებული მამრი ამ თვისებებს გადასცემს შთამომავლობას.

სურ. 8.6. ხარირემის ბრძოლა ფურების დაუფლებისათვის (ი. შმალგაუზენი, 1990).
ამჟამად გადარჩევის ამ ფორმას შიდასქესობრივი გადარჩევა ეწოდება. პირდაპირი მეტოქეობა, ძირითადად, ორი ფორმით იჩენს ხოლმე თავს. ერთ-ერთია ე.წ. „ქორის სტრატეგია“, რომლის დროსაც მეტოქეები მძიმედ იჭრებიან. ორთაბრძოლის შედეგად რომელიმე მათგანი იღუპება კიდეც. მაგალითად, ჩუბუკს (თოვლის ცხვარი, Ovis canadensis) აქვს მასიური რქები და ძლიერ გასქელებული შუბლის ძვალი. იგი სქესობრივი გადარჩევის შედეგად ჩამოყალიბდა. ვაცების მეტოქეობა იმდენად დაუნდობელია, რომ ბრძოლისას სასიკვდილო ჭრილობას აყენებენ ერთმანეთს (გრანტი, 1991). პირდაპირი მეტოქეობა მკაფიოდ არის გამოხატული კეთილშობილ ირემში (ჩერვუს ელაპჰუს). ხარირემს ყოველწლიურად ეცვლება რქები. შემოდგომით მყვირალობისას მას ბრძოლისათვის სრულყოფილად განვითარებული რქები აქვს. ხარირმები ბღავილით ერთიმეორეს უხმობენ. მათ შორის ორთაბრძოლა იმართება ფურებისათვის. ბრძოლა, მეტწილად, დაუნდობელ და მძაფრ ხასიათს ატარებს (იხ.: სურ. 8.6). დაახლოებით 25%25 შემთხვევაში ხარირმები მძიმედ იჭრებიან. გამარჯვებული ხარირემი ფურირმების ჯოგს ეუფლება. ფურირმებს რქები არა აქვთ. პარტნიორის ამორჩევა მათზე არ არის დამოკიდებული. გამარჯვებული ხარირემი სათავეში უდგას თავის ჰარამხანას, ამასთანავე, ის ფურირემთა ჯოგს იცავს უცხო პოტენციური მეტოქისაგან (მაკ-ფარლენდი, 1988).
მეტოქეობის მეორე ფორმაა ე.წ. „მტრედის სტრატეგია“. ამ შემთხვევაში მეტოქეები ემოციების დემონსტრაციით ან მუქარის პოზებით იფარგლებიან. ზოგჯერ კომფლიქტიც მოსდით, მაგრამ ცდილობენ ერთმანეთი არ დააზიანონ. მაგალითად, ანტილოპა ორიქსს (ryx gazella) აქვს 120 სმ-იანი გრძელი, სწორი და ბასრი რქები. ამ „იარაღით“ მამრი მარჯვედ იცავს თავს ძლიერი და მრისხანე მტრებისაგან. თუმცა, მამრები კონფლიქტში რქებს დიდი სიფრთხილით რიტუალურად იყენებენ და ერთმანეთს არ აზიანებენ. ისინი ძალის დემონსტრაციით იფარგლებიან. ჭიდილში დამარცხებული მამრი ტოვებს ტერიტორიას.
მეორე შემთხვევაში მამრები ერთმანეთს კონკურენციას უწევენ მდედრის მოხიბვლაში. ამ დროს მდედრი აქტიურია და ირჩევს მამრს. ამ შემთხვევაში ძლიერდება მეორეული სასქესო ნიშნები (მამრის მოკაზმულობა, კაშკაშა შეფერილობა და მისთ.). გადარჩევის ამ ფორმას სქესთაშორისი გადარჩევა ეწოდება. გამარჯვებული მამრის შემდგომი წვლილი გენოფონდის ფორმირებაში წარმატებულია. ამდაგვარი მამრის ნიშან-თვისებები და ქცევის თავისებურებები მომდევნო თაობებს გადაეცემა. მაგალითად, მამალი ხოხობი (hasianus colchicus) კაშკაშა მიმზიდველი შეფერილობით, გრძელი კუდითა და შესაბამისი საქორწილო რიტუალური ქცევით თავს აწონებს მურა-მორუხოდ შეფერილ მდედრს.
ზოგჯერ მკვეთრი შეფერილობა, გალობა, სპეციფიკური სუნი და სხვა ისეთი გენოტიპის გადარჩევას იწვევს, რომლებიც გამრავლებას უზრუნველყოფენ. მეორე მხრივ, ეს ნიშნები არღვევენ შენიღბვას და აქვეითებენ გადარჩენადობას. ამ უარყოფით თვისებებს ზოგჯერ დადებითი მნიშვნელობა აქვს მთელი სახეობისათვის, ვინაიდან სასარგებლო ნიშნების მქონე მამრები ტოვებენ შთამომავლობას და ამით თავისი წვლილი შეაქვთ პოპულაციის გენოფონდის ფორმირებაში. სქესობრივი გადარჩევის შედეგია სქესობრივი დიმორფიზმი.
ამრიგად სქესობრივი გადარჩევა არ არის ევოლუციის დამოუკიდებელი ფაქტორი. ის შიდასახეობრივი ბუნებრივი გადარჩევის კერძო შემთხვევას წარმოადგენს.
|
|
10.2.5.5 ბუნებრივი გადარჩევის სხვა ფორმები |
▲ზევით დაბრუნება |
ამჟამად მკვლევართა მიერ გადარჩევის სხვადასხვა ფორმაა გამოვლენილი, რომლებიც „დარვინული“ ანუ ზოგადი გადარჩევის კერძო ფორმას წარმოადგენს. განვიხილოთ გარკვეული ნაწილი. გარემოს ზოგიერთი ფაქტორი სელექციურ როლს ასრულებს. პოპულაციის სიმჭიდროვის ზრდის კვალობაზე მისი მოქმედების ეფექტურობაც ძლიერდება. მაშასადამე, მოქმედებას აძლიერებს სიმჭიდროვეზე დამოკიდებული ბუნებრივი გადარჩევა. ასე მაგალითად, პოპულაციაში სიმჭიდროვის ზრდის შესაბამისად დაავადებულთა რიცხვი და სიკვდილიანობა მატულობს. სიმჭიდროვის გარკვეული დონის მიღწევისას შესაძლებელია ის ეპიდემიაში გადაიზარდოს. როდესაც პოპულაციის სიმჭიდროვე ქვეითდება, დაავადების გამომწვევი ფაქტორი ნაკლებ აქტიური ხდება. პოპულაციის ძლიერი დისპერგირებისას ამ ფაქტორმა შეიძლება მოქმედება შეწყვიტოს. ამ პროცესს თან ახლავს პოპულაციაში დაავადებისადმი რეზისტენტული ფორმების ჩამოყალიბება (იხ.: §5.3).
კონკურენცია სიმჭიდროვით განპირობებულ ფაქტორს წარმოადგენს (იხ.: §8.3). მცენარეთა პოპულაციაზე ფიტოფაგი ცხოველების მოქმედება, ასევე, მტაცებლისა და მსხვერპლის ურთიერთმიმართება სიმჭიდროვეზე დამოკიდებული გადარჩევის დამადასტურებელი არგუმენეტებია. წესისამებრ, სიმჭიდროვეზე დამოკიდებული გადარჩევა პოპულაციაში სიმჭიდროვესა და რიცხოვნობას ასტაბილიზებს. ის მოქმედებას მაშინ იწყებს, როცა პოპულაციის სიმჭიდროვე გარკვეულ ზღვარს მიაღწევს ან მას სცილდება.
მოვიყვანთ კიდევ ერთ სანიმუშო მაგალითს: ბუნებაში შეინიშნება გამრავლების ორი სახის სტრატეგია: ე.წ. r-სტრატეგია, რომელიც г-გადარჩევით მიიღწევა. ამ ფორმისათვის ნიშანდობლივია ხანმოკლე სასიცოცხლო ციკლი, ნაადრევი სქესობრივი მომწიფება, დიდი რაოდენობით შთამომავლობის მოცემის უნარი (ენერგიის დიდი ნაწილი დიდი რაოდენობით შთამომავლობის წარმოქმნას ხმარდება). ამდაგვარი სტრატეგიით ცოცხალი სისტემა ცვლად საარსებო პირობებს ეგუება. გამრავლების სტრატეგიის მეორე ფორმა К-სტრატეგიაა, რომელიც K-გადარჩევით ყალიბდება. ამ სტრატეგიას ხანგრძლივი სასიცოცხლო ციკლი ახასიათებს. ინდივიდები სქესობრივ სიმწიფეს გვიან აღწევენ და მცირერიცხოვან შთამომავლობას წარმოქმნიან. პოპულაციის ენერგიის დიდი ნაწილი მცირერიცხოვანი შთამომავლობის გამოზრდას ხმარდება. სტრატეგიის ამ ფორმას ცოცხალი სისტემა შედარებით სტაბილურ საარსებო გარემოში იყენებს.
ზოგჯერ პოპულაციაში იშვიათი გენოტიპის მფლობელ ინდივიდს პოპულაციის სხვა წევრებთან შედარებით, სელექციური უპირატესობა აქვს. როცა იშვიათი გენოტიპების კონცეტრაცია პოპულაციაში იზრდება, ინდივიდი კარგავს თავის უპირატესობას. უფრო მეტიც, იშვიათი გენოტიპი შეიძლება საზიანოც კი აღმოჩნდეს მფლობელი ინდივიდისათვის. ამ შემთხვევაში მოქმედებს სიხშირეზე დამოკიდებული ბუნებრივი გადარჩევა. სანიმუშოდ შეიძლება მიმიკრიის მოვლენა (იხ.: §10.1) დავასახელოთ. აპოსემიური შეფერილობის ინდივიდი-მოდელი გარემოში ბევრად უნდა ჭარბობდეს იმიტატორის რაოდენობას. პოტენციური მტერი - ოპერატორი, პირადი გამოცდილებით არკვევს მოდელის საკვებად ვარგისიანობას. საკვებისადმი უარყოფითი რეაქციის შედეგად (უსიამოვნო სუნი, ტოქსიკური მოქმედება და ა.შ.) ხდება დასწავლა. პოტენციური მტერი გარეგნობით ცნობს აპოსემიური შეფერილობის მოდელს (ასევე, მისგან ვერ ასხვავებს იმიტატორს), რომელმაც უარყოფითი რეაქცია გამოიწვია მასში, რის გამოც ამ უკანასკნელს ერიდება. მაშასადმე, იმიტატორის სელექციური მნიშვნელობა დიდია, როცა მისი სიხშირე გარემოში მცირეა. მაგრამ, თუ იმიტატორის რიცხოვნობამ მოიმატა, გაუტოლდა ან გდააჭარბა მოდელის რაოდენობას, მაშინ ოპერატორს (პოტენციურ მტერს) საკვებზე რეაქცია ეცვლება. ამ შემთხვევაში მიმიკრია თავის მნიშვნელობას კარგავს.
დროზოფილას ხელოვნურ პოპულაციაში ჩატარებული ექსპერიმენტით გამოირკვა, რომ მუტანტური და ნორმალური ფენოტიპის მქონე მამრების წარმატებული შეჯვარება მდედრებთან გარემოში მათ სიხშირეზეა დამოკიდებული. მდედრები უპირატესობას ანიჭებენ და პარტნიორად ირჩევენ ისეთ მამრებს, რომლებიც დაბალი სიხშირით გვხვდებიან პოპულაციაში.
გადარჩევის პროცესს, რომელიც მიმართულია კონკრეტული ინდივიდის ახლო ნათესავების ნიშან-თვისებების შენარჩუნებისაკენ ნათესაური გადარჩევა (in selection) ეწოდება. ტერმინი მოწოდებულია ჯ. მეინარდ სმიტის მიერ (1981). კონკრეტული ინდივიდისათვის ასეთი პირებია თავისი შთამომავლები, სიბსები, და სხვა ახლო ნათესავები. ნათესაური გადარჩევის შედეგადაა ჩამოყალიბებული საზოგადოებრივ მწერებში სტერილური კასტები. მაგალითად, ფუტკრის სკაში მუშა ფუტკრები (სტერილური მდედრები) ოჯახს უვლიან, იცავენ და სარჩოთი ამარაგებენ. მოვიყვანთ მეორე მაგალითს: ფრინველი, რომელიც კლავს აპოსემიური (გამაფრთხილებელი) შეფერილობის მწერს, გემოს გასინჯვის შემდეგ ადგენს, რომ იგი საკვებად გამოუსედაგარია. ფრინველი დასწავლით იმახსოვრებს ფერით სიგნალს. ამის შემდეგ ის აღარ ეკარება ამ სიგნალის მქონე მწერებს, რაც სასარგებლოა დაღუპული მწერის სიბსებისათვის, რომელნიც ანალოგიური შეფერილობის არიან.
გადარჩევის კერძო ფორმას წარმოადგენს მადესტაბილიზებელი გადარჩევა (ბელიაევი, 1975). გარეული ცხოველების მოშინაურებისას მასტაბილიზებელი გადარჩევის შესუსტება პოპულაციაში ცვალებადობის მკვეთრ გაძლიერებას იწვევს. გადარჩევის მადესტაბილიზებელი ფორმა არღვევს ორგანიზმში ადრე ჩამოყალიბებულ კორელაციურ ურთიერთდამოკიდებულებებს. მოშინაურების პროცეში მტაცებელ ცხოველებში ხელოვნური გადარჩევა, რომელიც აგრესიის დაქვეითებისაკენაა მიმართული, ასევე, გამრავლების ციკლის დესტაბილიზაციას იწვევს - ცხოველები მთელი წლის მანძილზე მრავლდებიან.
|
|
10.2.5.6 ინდივიდური და ჯგუფური გადარჩევა |
▲ზევით დაბრუნება |
ჩვეულებრივ, ბუნებრივი გადარჩევის ორ - ინდივიდურსა და ჯგუფურ - ფორმას გამოყოფენ. ინდივიდური გადარჩევა მიმართულია პოპულაციაში ცალკეული ინდივიდის დიფერენცირებული გამრავლებისაკენ. არსებობისათვის ბრძოლაში (შეჯიბრში) წარმატებულია ის ინდივიდი, რომელსაც მეტი წვლილი შეაქვს შემდგომი გენოფონდის ფორმირებაში. მაშასადამე, ევოლუციურად წარმატებული ინდივიდი დიდი რაოდენობით იძლევა შთამომავლობას, რომელიც სქესმწიფე ასაკს აღწევს და მრავლდება. ინდივიდური გადარჩევა პოპულაციის შიგნით მოქმედებს და ინდივიდთა შეჯიბრზეა დაფუძნებული. არსებობისათვის ბრძოლაში წარმატებული ინდივიდები შემდგომ თაობაში მემკვიდრეობით გადასცემენ თავიანთ ნიშან-თვისებათა თავისებურებებს (გენოტიპებს). ბუნებრივი გადარჩევის ზეწოლა იწვევს გენოფონდის გარკვეული მიმართულებით შეუქცევად შეცვლას. მემკვიდრული გადახრები, რომლითაც შეცვლილ გარემოში ინდივიდები უპირატესობით სარგებლობენ, თანდათანობით გროვდება და პოპულაციებში ფიქსირდება. ამ პროცესს თან ახლავს წინაპრებსა და შთამომავლებს შორის არსებითი განსხვავების ჩამოყალიბება. ამა თუ იმ სახეობის ან სხვა ტაქსონების ნებისმიერი ნიშანი თუ თვისება ბუნებრივი გადარჩევის მოქმედებით ინდივიდურ განსხვავებათა საფუძველზე ფორმირდება.
ჩ. დარვინი მიიჩნევდა, რომ ბუნებრივი გადარჩევა ყველაზე შეგუებულ ფორმათა გდარჩენაა, რომელიც მიმდინარეობს როგორც პოპულაციის შემადგენელ წევრთა შორის, ისე ჯგუფებს (დემები, პოპულაციები, ქვესახეობები და სხვ.) შორისაც, რადგანაც ჯგუფური გადარჩევა ჯგუფში გაერთიანებულ ინდივიდთა გადარჩევას ემყარება, ამიტომ ევოლუციაში წამყვანია ინდივიდური გადარჩევა. ჯგუფური გადარჩევა არის კონკრეტული ჯგუფის ინდივიდების წარმატებული გამრავლება სხვა ჯგუფებთან შედარებით. ჯგუფური გადარჩევის დროს შეიძლება მასში გამყარდეს ჯგუფისათვის სასარგებლო ნიშანი. ზოგჯერ ჯგუფისთვის სასარგებლო ნიშან-თვისება მასში გაერთიანებული ცალკეული წევრისთვის შეიძლება საზიანო აღმოჩნდეს. ჯგუფური გადარჩევის დროს სხვადასხვა ჯგუფებში გაერთიანებული ინდივიდები ერთმანეთს ეჯიბრებიან, ჩამოაყალიბონ და შეინარჩუნონ ზეორგანიზმული ერთიანი სრულყოფილი სისტემები.
ჯგუფური გადარჩევის ერთ-ერთი ფორმაა ნათესაური გადარჩევა. მისი მოქმედებით ჯგუფში ალტრუისტული ქცევის განმსაზღვრელი გენები მყარდება. მონათესავე ინდივიდების გენოტიპები გაცილებით უფრო მსგავსია, ვიდრე არამონათესავე ფორმებისა. თუ დემის შემადგენელი რომელიმე ინდივიდი თავს წირავს სიბსების გადასარჩენად, რითაც მათ პოტენციურ გამრავლებას საფრთხე აღარ ემუქრება, ამით ალტრუისტი ინდივიდი (არაპირდაპირი გზით) საკუთარი გენების სიხშირეს ზრდის მომავალ გენოფონდში. მიუხედავად იმისა, რომ ნიშანი უარყოფითია მფლობელისათვის, ნათესაური გადარჩევის მოქმედებით მისი ფიქსაცია მაინც ხორციელდება პოპულაციაში (ცაგარელი, 2004).
ჯგუფური გადარჩევის მოქმედების შედეგად შეიძლება არეალიდან ერთმა კონკრეტულმა ჯგუფმა მეორე გააძევოს (მოქმედებს ე.წ. კონკურენტული გამოთიშვის პრინციპი). არეალში იზღუდება ჯგუფური მრავალფეროვნება. ჯგუფური გადარჩევის მოქმედებით, წესისამებრ, სახეობაში ჯგუფური განსხვავებები ყალიბდება და მტკიცდება. სახეობა პოლიტიკურ სტრუქტურას იძენს. ამით ცალკეულ ჯგუფში ქვეითდება ბუნებრივი გადარჩევის ზეწოლის დონე. სამაგიეროდ იზრდება სიცოცხლის ზეწოლა ერთეულ ფართობზე. ასე მაგალითად, ნაძვნარში გავრცელებული ამერიკული ასპუჭაკას ხუთი სიმპატიური სახეობა კვებითი სპეციალიზაციით განსხვავდებიან. ისინი ხეების სხვადასხვა ნაწილებიდან, სხვადასხვა ხერხით მოიპოვებენ მწერებს (იხ.: სურ. 7.8).
ამჟამად ჯგუფური გადარჩევის სხვადასხვა ფორმაა გამოკვლეული. ყველა მათგანი შიდაპოპულაციურ ბუნებრივ გადარჩევას ეფუძნება, რადგანაც კონკურენცია მიმდინარეობს არა აბსტრაქტულ ტაქსონებს შორის, არამედ სხვადასხვა ტაქსონებში გაერთიანებულ პოპულაციის წევრებს შორის. ევოლუციის პროცესში ადაპტაციები ყალიბდება ინდივიდური გადარჩევის შედეგად. ჯგუფური გადარჩევა მზა ადაპტური სისტემებიდან რომელიმე მათგანს (ჯგუფისათვის სასარგებლოს) ირჩევს. ინდივიდურ და ჯგუფურ გადარჩევას შორის გარდამავალი ფორმებია - ნათესაური, დემების, პოპულაციური ჯგუფების მიხედვით მიმდინარე გადარჩევა.
სურ. 8.7. ამერიკული ასპუჭაკის (endroica) ხუთი სახეობის საკვების მოსაპოვებელი ადგილის ლოკალიზაცია ნაძვნარის ტყეში I-D. tigrina, II-D. fusca, III - D. virens, IV-D. castanea V-D. coronata (რ. მაკარტური, 1958).
|
|
10.2.6 8.9. ბუნებრივი გადარჩევის მნიშვნელობა |
▲ზევით დაბრუნება |
პოპულაციის განვითარებას მხოლოდ ბუნებრივი გადარჩევა წარმართავს. იგი ბიოლოგიური ევოლუციის ერთადერთი მაორგანიზებელი და წარმმართველი ფაქტორია. ამ პროცესში ბუნებრივი გადარჩევა იყენებს ელემენტარულ ევოლუციურ მასალას - მუტაციებს. იგი მოქმედების პროცესში იქვემდებარებს ევოლუციის დანარჩენ ელემენტარულ ფაქტორებს, კერძოდ მუტაციურ პროცესს, გენთა ნაკადს და გენთა დრეიფს. ამასთანავე, ბუნებრივი გადარჩევა აერთიანებს და ერთიმეორესთან ათანხმებს ყველა ელემენტარული ფაქტორის მოქმედებას. ამდენად იგი მიკროევოლუციის მაკონტროლებელი და მაორგანიზებელი ფაქტორია. პოპულაციაში ბუნებრივი გადარჩევის მოქმედება სამ შესაძლო შედეგს იწვევს, კერძოდ:
1. უცვლელი სახით ინახავს ადრე ჩამოყალიბებულ ნიშნებს, ვინაიდან იგი სასარგებლოა ინდივიდებისათვის კონკრეტულ საარსებო პირობებში. გადარჩევით ძველი, უკეთ შეგუებული ვარიანტების (გენოტიპების) შენარჩუნება ხდება.
2. ყალიბდება ახალი ადაპტაციური ნიშნები, რომლებიც ძველის (ახალ საარსებო პირობებში ადაპტაციურ ღირებულება დაკარგულის) ადგილს იკავებენ. გადარჩევით ახალი, უკეთ შეგუებული ვარიანტებით (გენოტიპებით) იცვლება ძველი.
3. ხდება ერთდროულად მკვეთრად განსხვავებული ნიშნის რამდენიმე ვარიანტის (გენოტიპის) შენარჩუნება.
ბუნებრივი გადარჩევის როლი მხოლოდ ნაკლებშეგუებულ ინდივიდთა ელიმინაციით როდი შემოიფარგლება. მისი მოქმედებით ყალიბდება ახალ-ახალი ადაპტაციები, რისი მეშვეობითაც ინდივიდები ეგუებიან შეცვლილ საარსებო გარემოს. ამდენად, ბუნებრივი გადარჩევა ერთადერთ შემოქმედებით ფაქტორს წარმოადგენს. იგი ერთი და იმავე მასალიდან (მემკვიდრული ცვალებადობა) საარსებო პირობებისა და გადარჩევის მიმართულების მიხედვით აყალიბებს ნაირგვარ ადაპტაციურ ნიშნებს. განსხვავებული გენოტიპების მქონე ინდივიდების დიფერენცირებული გამრავლება თვისებრივად ახალი ფორმების ჩამოყალიბებას იწვევს. ბუნებრივი გადარჩევის მოქმედება შეიძლება მოქანდაკის შემოქმედებით პროცესს შევადაროთ. უფორმო თიხის მასისაგან მოქანდაკე თანდათანობით საოცარ ქმნილებებს ძერწავს. ბუნებრივი გადარჩევის შედარება მოქანდაკის საქმიანობასთან ნაწილობრივ შეიძლება. მოქანდაკეს აქვს მომავალი ქმნილების გეგმა, ბუნებრივ გადარჩევას კი - არა. მისი მოქმედების მიმართულება სტიქიურად წარმოიქმნება. სამაგიეროდ ბუნებრივი გადარჩევის პროცესში დიდ როლს ასრულებს დროის ფაქტორი. იგი განუსაზღვრელი დროის (მილიონობით და მილიარდობით წელი) განმავლობაში მოქმედებს და განუსაზღვრელად დიდ მემკვიდრულ მასალას იყენებს. ბიოლოგიურ დროში (თაობათა მანძილზე) მეტად უმნიშვნელო ცვლილებების სუმაციით მნიშვნელოვანი ცვლილება ყალიბდება. ნაკლებ სრულყოფილი ფორმებიდან უფრო სრულყოფილი ფორმები მიიღება.
|
|
11 § 9. შიგაპოპულაციური (გენეტიკური) პოლიმორფიზმი |
▲ზევით დაბრუნება |
|
|
11.1 9.1 მორფოლოგიური პოლიმორფიზმი |
▲ზევით დაბრუნება |
ნებისმიერი პოპულაციისათვის ნიშანდობლივია გენეტიკური ცვალებადობა. მისი ერთ-ერთი მნიშვნელოვანი გამოხატულებაა პოპულაციური პოლიმორფიზმი. ეს კანონზომიერება 1940 წელს დაადგინა ინგლისელმა გენეტიკოსმა და ეკოლოგმა ე. ფორდმა. გენეტიკური პოლიმორფიზმი ეწოდება პოპულაციაში რეგულარულად და ერთდროულად ორი ან მეტი გენოტიპურად მკვეთრად განსხვავებული ფორმის არსებობას, ამასთანავე ყველაზე იშვიათი ფორმის სიხშირე იმდენად მაღალია, რომ მისი შენარჩუნება შეუძლებელია მხოლოდ მუტაციური ზეწოლით აიხსნას.
პოლიმორფიზმი ცოცხალ სამყაროში ფართოდ გავრცელებული მოვლენაა. იგი ბუნებრივ პოპულაციებში მნიშვნელოვან ადაპტურ როლს ასრულებს. პოლიმორფიზმი ბუნებრივი გადარჩევის მოქმედების შედეგია, რომელიც ხელს უწყობს პოპულაციაში განსხვავებული მორფების ჩამოყალიბებას და შენარჩუნებას. ორი ან მეტი ფენოტიპურად (ე.ი. გენეტიკურად) განსხვავებული მორფა პოპულაციაში თაობათა განმავლობაში (ბიოლოგიურ დროში ხანგრძლივად) გაწონასწორებულ მდგომარეობაში იმყოფება. პოპულაციაში ფენოტიპურად (ბუნებრივია გენოტიპურად) განსხვავებული მორფების არსებობა, მიუთითებს, რომ მოქმედებს ბუნებრივი გადარჩევის ზეწოლა, რომელიც ხელს უწყობს პოლიმორფიზმის შენარჩუნებას.
პოპულაციური პოლიმორფიზმის კრიტერიუმად მიღებულია, რომ ყველაზე იშვიათი ფორმის სიხშირე ≥5%25 უნდა შეადგენდეს. ზოგიერთ შემთხვევაში ამ ზღვარს≥1%25-მდე დაწევენ ხოლმე, რაც მოვლენის არსს არ ცვლის. ნებისმიერი გენოტიპის სიხშირემ მუტაციური ზეწოლით პოპულაციაში შეუძლებელია 1%25-იან ზღვარს გადააჭარბოს. ამისათვის აუცილებელია სხვა ევოლუციური ფაქტორის, კერძოდ ბუნებრივი გადარჩევის მონაწილეობა.
საჭიროა განიმარტოს, თუ რით განსხვავდება გენეტიკური ჰეტეროგენულობა პოპულაციის გენეტიკური პოლიმორფიზმისაგან. გენეტიკური ჰეტეროგენულობა არის პოპულაციაში ნებისმიერი, თუნდაც უმნიშვნელო გენეტიკური ნაირგვარობა, რომელიც მუტაციური ან რეკომბინაციული პროცესითაა გამოწვეული. პოპულაციური პოლიმორფიზმი კი, ზემოთ მოტანილი განსაზღვრებიდან გამომდინარე, არის პოპულაციაში ერთდროულად გენოტიპურად განსხვავებული მრავალ კლასთა ხანგრძლივად არსებობა. მათი მუდმივი შენარჩუნება ბუნებრივი გადარჩევით მიიღწევა.
ორგანულ სამყაროში პოლიმორფიზმის მრავალი ფორმა არის გამოვლენილი. სადღეისოდ პოპულაციური პოლიმორფიზმის ერთიანი, საყოველთაოდ მიღებული კლასიფიკაცია არ გაგვაჩნია, რაც ამ მოვლენის სირთულეზე მიუთითებს. წარმოშობისა და შენარჩუნების მექანიზმების მიხედვით პოლიმორფიზმის ყველა შემთხვევას ორ დიდ ჯგუფად ყოფენ: 1. ბალანსირებული (სტაბილური); 2. გარდამავალი (ტრანზიტული). ბალანსირებული პოლიმორფიზმის შემთხვევაში პოპულაციაში გენოტიპურად განსხვავებულ ფორმებს შორის მყარდება წონასწორული თანაფარდობა.
ბალანსირებულ პოლიმორფიზმში გამოყოფენ ჰეტეროზიგოტულ და ადაპტურ პოლიმორფიზმს.
როდესაც ბუნებრივი გადარჩევის ზეწოლა ჰეტეროზიგოტების სასარგებლოდ მიმდინარეობს, მაშინ ყალიბდება ჰეტეროზიგოტული პოლიმორფიზმი. ჰეტეროზიგოტული პოლიმორფიზმის კლასიკური მაგალითია ნამგლისებრუჯრედიანი ანემიის HBS ალელის ფიქსაცია ადამიანის იმ პოპულაციაში, სადაც მალარიის პლაზმოდიუმი Pl. falciparum-ია გავრცელებული. ჰეტეროზიგოტების HBA/HBS ერითროციტებში მალარიის პლაზმოდიუმი ვითარდება სწრაფად, გამეტოციტები ნაადრევად ყალიბდება და პარაზიტი უმოკლეს დროში ტოვებს სისხლს. ამიტომ ჰეტეროზიგოტებში ავადმყოფობა ხანმოკლეა. ყოველივე აღნიშნულის გამო ჰეტეროზიგოტები დაავადებისადმი რეზისტენტულებია (იხ. §8.6). ჰეტეროზიგოტები ორივე ჰომოზიგოტური კლასის მიმართ უპირატესობას ინარჩუნებენ. რეცესიული HBS/HBS ჰომოზიგოტები ანემიის მძიმე ფორმით არიან დაავადებული. ისინი ფლობენ ჰემოგლობინის შ ვარიანტს (იხ. სურ. 14.2). მასში ၢ-ჯაჭვის მე-6 პოზიციაში უარყოფითი მუხტის მქონე გლუტამინის მჟავა შეცვლილია ნეიტრალური ვალინით. ჰიპოქსიის დროს შ ჰემოგლობინი წარმოქმნის კრისტალის მსგავს სტრუქტურებს, რის შედეგადაც ერითროციტს ეცვლება მორფოლოგია. იგი ნამგლისებრ ფორმას იძენს. დომინანტი ჰომოზიგოტები HBA/HBA ფლობენ ჰემოგლობინის ნორმალურ A ვარიანტს. პლაზმოდიუმი ერითროციტებში ნორმალურად ვითარდება. ასეთ პირებში მალარიის დაავადება მძიმედ მიმდინარეობს.
შეისწავლეს დროზოფილას ხელოვნური გაწონასწორებული პოპულაცია, რომელიც რუხი და შავი (იწვევს რეცესიული გენი ებონი) შეფერილობის ინდივიდებისაგან შედგებოდა. თითოეული კლასის სიხშირე საწყის პოპულაციაში 50%25-ს შეადგენდა (ე.ი. Q=0,5). რამდენიმე თაობის შემდეგ შავსხეულიანი ინდივიდების რაოდენობა მკვეთრად (10%25-მდე) შემცირდა. მომდევნო თაობებში კი ალელთა კონცენტრაცია პოპულაციაში სტაბილური გახდა. გამოირკვა, რომ ჰეტეროზიგოტები გაცილებით მაღალი სიცოცხლისუნარიანობით გამოირჩევიან, ვიდრე დომინანტური და რეცესიული ალელების მფლობელი ჰომოზიგოტები. ჰეტეროზიგოტების სელექციურ უპირატესობას ადგილი აქვს ზედომინანტობის (მონოგენური ჰეტეროზისი) დროს.
სურ. 9.1. ჰეტეროსტილია. ერთი პოპულაციის სხვადასხვა მცენარეს მტვრიანები და სვეტი სხვადასხვა დონეზე აქვს განლაგებული: ზოგიერთ მცენარეს აქვს გრძელი სვეტი და მტვრიანები ქვემოთაა (2), ზოგსაც - მოკლე სვეტი და მტვრიანები ზემოთაა განლაგებული (1).
მონოგენური ჰეტეროზისი არის ჰეტეროზიგოტული პოლიმორფიზმის უკიდურესი გამოვლენა. ამ მოვლენის დროს ჰეტეროზიგოტულ მდგომარეობაში გენის დომინანტური ალელი გაცილებით ძლიერ ფუნქციონირებს და ავლენს თავის მოქმედებას, ვიდრე ჰომოზიგოტურ მდგომარეობაში. პოლიმორფიზმის ეს ფორმა პირველად 1934 წელს გამოავლინა რ.ნებუსმა და ლ.კლინგმა. მათ დაადგინეს, რომ კუტკალიას (Apotettix eurycephalus) რეცესიული ლეტალური მუტაციის მფლობელი ჰეტეროზიგოტები ჰომოზიგოტებთან შედარებით მეტი სიცოცხლისუნარიანობით გამოირჩეოდნენ.
მონოგენური ჰეტეროზისი სასოფლო სამეურნეო კულტურებში (სიმინდი, ქერი) გამოავლინეს გამოჩენილმა გენეტიკოსებმა ჯ.სტადლერმა, ა.გუსტაფსონმა და გ.შტუბემ. მათ მიერ ინდუცირებული მუტაციების მქონე მონოჰეტეროზიგოტები მეტ მოსავალს იძლევა ვიდრე ჰომოზიგოტები. რუსმა გენეტიკოსმა დ.ბელიაევმა დაადგინა, რომ მოშინაურებულ წაულებში ალეუტური (მოცისფრო რუხი) და მოვერცხლისფრო ლურჯი შეფერილობის განმსაზღვრელი მუტირებული ალელები ჰეტეროზიგოტებში განაპირობებდნენ მაღალნაყოფიერებასა და სიცოცხლის უნარიან შთამომავლობას.
ზოგჯერ პოლიმორფიზმისთვის ნიშანდობლივია მკვეთრი გენეტიკური დეტერმინაცია. ენტომოფილურ ყვავილოვან მცენარეებში (ცოცხმაგარა, მატიტელა, ფურისულა, კესანე და მისთ.) შეინიშნება ჰეტეროსტილია. ზოგიერთ ეგზემპლარს მოეპოვება გრძელსვეტიანი ყვავილები, ხოლო სამტვერეები დინგსდაბლაა განლაგებული, ზოგიერთს კი პირუკუ. მათ ობლიგატურად აღენიშნებათ დათიშვა ორ ფენოტიპურ კლასად: ჰეტეროზიგოტებს (Bb) გააჩნიათ გრძელი სვეტი, ხოლო რეცესიულ ჰიმოზიგოტებს (bb) მოკლე სვეტი (იხ. სურ. 9.1). ბუნებაში შეჯვარდებიან მხოლოდ განსხვავებული გენოტიპის მცენარეები, რითაც თავიდან იცილებენ თვითდამტვერვასა და ინბრიდინგს. პოლიმორფიზმის გამოვლენაა ასევე დიქოგამია (ბუტკოსა და მტვრიანების არაერთდროული მომწიფება) ენტემოფილურ მცენარეებს აღენიშნებათ ორი განსხვავებული ფორმით: პროტანდრია (მამრობითი ორგანო მდედრობითზე ადრე მწიფდება) და პროტეროგინეა (მდედრობითი ორგანო მამრობითზე ადრე მწიფდება). ამდაგვარი ადაპტაცია ხელს უწყობს ჯვარედინ დამტვერვას, და შესაბამისად პოპულაციაში იზრდება კომბინაციური ცვალებადობა.
სქესობრივი დიმორფიზმი ცალსქესიანი სახეობის არსებობის აუცილებელი ფორმაა. ერთ-ერთი სქესის ობლიგატური ჰეტეროზიგოტულობით უზრუნველყოფილია დათიშვა თანაფარდობით 1:1 (სქესის პირველადი თანაფარდობა). მდედრებსა და მამრების განსხვავებული ცხოველმყოფელობა ცვლის სქესის მეორად და მესამეულ თანაფარდობას. ეს მაჩვენებლები ადაპტური მნიშვნელობისაა და ბუნებრივი გადარჩევით რეგულირდება.
გენეტიკურად განსხვავებული ფორმები, როდესაც განსხვავებულ ეკოლოგიურ პირობებში ექვემდებარებიან გადარჩევას, ყალიბდება ადაპტური პოლიმორფიზმი. საილუსტრაციოდ მოვიყვანოთ რამდენიმე მაგალითი.
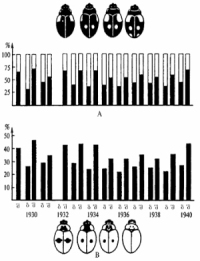
სურ. 9.2. მორფოლოგიური პოლიმორფიზმი ორწერტილიან ჭიამაიას (Adalia bipunctata) ბერლინის პოპულაციაში. A - შავი და წითელი ფორმების პროცენტული თანაფარდობა, გაზაფხულზე (G) და შემოდგომაზე (S). B - შავი შეფერილობის განმსაზღვრელი დომინანტური ალელის სიხშირე (ნ. ტიმოფეევ-რესოვსკი და სხვ., 1977).
გამოჩენილი რუსი გენეტიკოსი და ევოლუციონისტი ნ. ტიმოფეევ-რესოვსკი თერთმეტი წლის (1930-1940) განმავლობაში იკვლევდა ბერლინის გარეუბანში გავრცელებულ ორწერტილიანი ჭიამაიების (Adalia bipunctata) პოპულაციებში წინა ფრთების (elitra) შეფერილობის პოლიმორფულობას. მორფათა ერთ ნაწილს გააჩნდა წითელ ფონზე შავი ლაქები (ე.წ. წითელი ფორმები), ხოლო, მეორე ნაწილს წითელი ლაქები შავ ფონზე (ე.წ. შავი ფორმები). შავი შეფერილობა დომინანტობს წითელზე, ხოლო ლაქების ფორმას მრავლობითი ალელები განსაზღვრავენ. წელიწადში ორჯერ გაზაფხულსა და შემოდგომაზე ისაზღვრებოდა წითელი და შავი ფორმების სიხშირე პოპულაციაში. შემოდგომით - გამოზამთრებამდე პოპულაციაში ჭარბობდა შავი ფორმების სიხშირე. ჭიამაიები იკრიბებიან ჯგუფებად, ქვათა შორის ღრმა ნაპრალებში ძვრებიან, სადაც იზამთრებენ. გაზაფხულზე ნაპრალებიდან გამომძვრალ გამოზამთრებულ ჭიამაიებში სიხშირით ჭარბობდა წითელი ფორმები. შავი ფორმების რაოდენობა 30-45%25-ს არ აღემატებოდა (იხ. სურ. 9.2) ორი ფორმის პოპულაციაში დინამიკა გამოწვეულია იმით, რომ წითელი ფორმები კარგად იტანენ ზამთრის სუსხს, ხოლო შავი ფორმები მეტი ინტენსივობით მრავლდებიან ზაფხულში. ბუნებრივი გადარჩევა იწვევს წითელი ფორმების შენარჩუნებას ზამთარში, ხოლო შავი შეფერილობის ფორმებისას ზაფხულში. განსხვავებული მიმართულებით მომქმედი გადარჩევის ზეწოლით პოპულაციაში ხანგრძლივი დროის განმავლობაში ყველა მორფაა შენარჩუნებული. გადარჩევის ამგვარი სტრატეგია, საზოგადოდ მთელი პოპულაციისათვისაა სასარგებლო.
ჭიამაიების ბერლინის პოპულაციებში გამოვლენილი პროცესები არ აღმოჩნდა უნივერსალური და არ ასახავს ზოგად კანონზომიერებებს. რუსმა გენეტიკოსმა ი. ზახაროვმა შეისწავლა (1961-1981წწ.) პეტერბურგსა და მის გარეუბნებში გავრცელებულ ორწერიტილიან ჭიამაიებში პოლიმორფიზმი. განსხვავებული მორფების წარმოშობა ინდუსტრიული მელანიზმით იყო გამოწვეული. ქარხნების გამონაბოლქვით დაბინძურებულ რაიონებში ჭიამაიების პოპულაციებში ჭარბობენ შავი ფორმები, ხოლო ნაკლებ ან დაუბინძურებელი რაიონების პოპულაციებში წითელი ფორმები. ამრიგად, განსხვავებულ სიტუაციაში ერთი და იგივე პოლიმორფული სისტემა განსხვავებული სტრატეგიით ვლინდება. გარემოს კონკრეტულ პირობებში იცვლება გადარჩევის ზეწოლის მიმართულება და ხასიათი.
ადაპტური პოლიმორფიზმის კლასიკური ნიმუში გამოავლინა პ. შეპარდმა თანაავტორებთან ერთად ბაღის ლოკოკინაში (Cepaea nemralis). იგი ფართოდაა გავრცელებული დასავლეთ ევროპაში. ლოკოკინები განსხვავდებიან ნიჟარის შეფერილობით. მათ აქვთ ყვითელი (როდესაც მასში ცოცხალი ცხოველია, მწვანედ გამოიყურება), ყავისფერი (ნაირგვარი ელფერით), ვარდისფერი, ნარინჯისფერი ან წითელი ნიჟარა. ლოკოკინები გასხვავდებიან ნიჟარის ბაგის (პერისტომი) შეფერილობით, რომელიც თეთრი, ვარდისფერი ან მუქი ყავისფერია. ნიჟარა უზოლო ან ზოლიანია (სიგრძივად გასდევს 1-5 მუქი ზოლი). ყველა ჩამოთვლილი ნიშანი გენეტიკურად არის დეტერმინირებული. ნიჟარის შეფერილობას Y გენის მრავლობითი ალელები განსაზღვრავენ. ყავისფერის განმსაზღვრელი YB ალელი დომინანტობს ვარდისფერზე - YR, ორივე კი ყვითელისაზე Y, ე.ი. YB>YR>Y; უზოლობა დომინანტური ნიშანია (U), ზოლიანობა კი რეცესიული (U). ნიჟარის შეფერილობისა და ზოლიანობის განმსაზღვრელი გენები ძლიერ შეჭიდულია, და ქმნიან სუპერგენს. იგი მოქმედებს და მემკვიდრეობს, როგორც ერთიანი გენეტიკური ერთეული (იხ. სურ. 9.3). დამოუკიდებელი გენები განსაზღვრავენ ბაგისა და ზოლის შეფერილობას, ასევე ზოლების რაოდენობას.
სურ. 9.3. ნიჟარის შეფერილობა და ზოლიანობა ბაღის ლოკოკინაში (Cepaea nemoralis) A - ყვითელი უზოლო ნიჟარა; B - ვარდისფერი ნიჟარა; C - ყავისფერი ნიჟარა; D - ყავისფერი ორზოლიანი ნიჟარა E - ყავისფერი ხუთზოლიანი ნიჟარა (ფ. ჰედრიკი, 2004).
ლოკოკინებით ფრინველები (ძირითადად შაშვი) იკვებებიან. ისინი ქვაზე ამტვრევენ ნიჟარას და გამოაქვთ შიგთავსი. ამდაგვარ „სამტეხლოებზე“ დამსხვრეული ნიჟარების ანალიზით, ამავე გარემოში გავრცელებული ლოკოკინების შესწავლით, გამოირკვა რომ მოქმედებს სელექციური ფაქტორები. მეტწილად ნადგურდებიან ფონისაგან განსხვავებული ინდივიდები. სადაც გარემო ფონი ერთგვაროვანია (მაგ. ბალახი, ტყის საფარი) სელექციური უპირატესობა ყვითელ და ყავისფერ უზოლო ფორმებს აქვთ ვინაიდან „სამტეხლოებზე“ ამდაგვარი ნიჟარები საკმაოდ მცირე რაოდენობით აღმოჩნდა. ამავე გარემოში ჭრელ ფონზე (მაგ., საძოვრები, სხლტეები) მუქ ზოლიანი ფორმები იძენენ სელექციურ უპირატესობას. ლოკოკინას მრავალრიცხოვანი პოლიმორფული პოპულაცია საკმაოდ დიდ ტერიტორიზეა გავრცელებული, რომელთაც ფონით (მცენარული საფარი, რელიეფი და მისი შეფერილობა...) განსხვავებული ადგილი უჭირავთ. პოპულაციის საარსებო გარემოს ფონი იცვლება წლიური სეზონების მიხედვით. მართალია, ფრინველების მიერ ფონისაგან განსხვავებული ინდივიდები განუწყვეტლივ ნადგურდება ვინაიდან არცერთ მათგანს არ გააჩნია სელექციური უპირატესობა, წლიდან წლამდე განსხვავებულ ფორმათა თანაფარდობა პოპულაციაში უცვლელია.
ბაღის ლოკოკინაში შეფერილობის პოლიმორფიზმი მხოლოდ იმით ვერ აიხსნება, რომ ფრინველები მათ შერჩევით ანადგურებენ. ზოგიერთ არეალში გარემო პირობების ჰეტეროგენულობის მიუხედავად სხვადასხვა შეფერილობის ვარიანტთა შეხვედრის სიხშირე ერთნაირია. სხვა არეალში სადაც გარემო პირობები ერთნაირია ნიჟარის შეფერილობაში კლინური ცვალებადობა შეინიშნება. ამ მოვლენას არეალის ეფექტი ეწოდება. იგი სხვადასხვა შეფერილობის ლოკოკინებზე ჰეტეროგენული გარემოს და კლიმატური ფაქტორების სხვადასხვაგვარი ზემოქმედების შედეგია. გამოთქმულია მოსაზრება, რომ ნიჟარის შეფერილობის პოლიმორფიზმის შენარჩუნება გარემო პირობებისა და გენეტიკური ფაქტორების რთული ურთიერთქმედებით არის გამოწვეული. მათი შეფარდებითი მნიშვნელობა არეალის შესაბამისად ვარირებს (ჰედერიკი, 2003).
ბალანსირებული პოლიმორფიზმის სანიმუშო მაგალითია, სისხლის ABO ჯგუფის ალელთა გავრცელება ადამიანის სხვადასხვა პოპულაციაში. სისხლის A, B, AB, 0 ჯგუფის სიხშირე ადამიანთა სხვადასხვა პოპულაციაში შეიძლება დიდ ფარგლებში ვარირებდეს, მაგრამ კონკრეტულ პოპულაციაში მათი თანაფარდობა უცვლელი რჩება, ვინაიდან არცერთი მათგანისათვის არაა ნიშანდობლივი სელექციური უპირატესობა.
გარდამავალი პოლიმორფიზმი ყალიბდება იმ შემთხვევაში, როდესაც პოპულაციის შემადგენელი განსხვავებული მორფები, ბუნებრივი გადარჩევის ძლიერ ზეწოლას ექვემდებარებიან. საილუსტრაციოდ ჩვენ მიერ განხილული არყის ხის მზომელას პეპლებში (Biston betularia) ინდუსტრიული მელანიზმის მოვლენა შეიძლება დავასახელოთ (იხ. §9.2).
პოლიმორფიზმის საილუსტრაციოდ მოყვანილი მაგალითები ცხადყოფენ, რომ პოპულაციის შემადგენელი გენეტიკური კლასები მორფოლოგიურად მკაფიოდ განსხვავდებიან და მათ შორის განაწილებულია ფუნქციები. ცალკეული გენეტიკური კლასი მისთვის ნიშანდობლივ ფუნქციას ასრულებს. გარემოს ცვლადი პირობებისადმი მორფები სივრცესა და დროში სპეციფიკურად არიან სპეციალიზებული. ამდაგვარი განსხვავების შედეგი მათ სეზონურ ციკლურობასა და სივრცობრივ დიფერენცირებაში ვლინდება. მორფებად დიფერენცირებით იზრდება პოპულაციის პლასტიურობა და ერთიანობა.
|
|
11.2 9.2. ქრომოსომული პოლიმორფიზმი |
▲ზევით დაბრუნება |
ქრომოსომულ პოლმორფიზმს სპონტანურად ინდუცირებული ქრომოსომული და გენომური მუტაციები განაპირობებენ. იგი ადაპტურ როლს ასრულებს პოპულაციაში. გამოყოფენ ქრომოსომული პოლიმორფიზმის შემდეგ ფორმებს:
1. ინვერსიული პოლიმორფიზმი.
2. რობერტსონული ტრანსლოკაციების პოლიმორფიზმი.
3. დამატებითი ქრომოსომების ფორმისა და რაოდენობის პოლიმორფიზმი.
4. სასქესო ქრომოსომების ფორმისა და რაოდენობის პოლიმორფიზმი.
5. ქრომოსომაში ჰეტეროქრომატინის რაოდენობისა და განაწილების პოლიმორფიზმი
|
|
11.2.1 ინვერსიული პოლიმორფიზმი |
▲ზევით დაბრუნება |
ინვერსიული პოლიმორფიზმი 1937 წ. ერთმანეთისგან დამოუკიდებლად პირველად გამოავლინეს და შეისწავლეს რუსმა გენეტიკოსმა ნ.დუბინინმა და ამერიკელმა ევოლუციონისტმა ფ.დობჟანსკიმ თანაავტორებთან ერთად. მათ დაადგინეს, რომ ქრომოსომული ცვლილებების (ინვერსიების) შენარჩუნება პოპულაციებში ბუნებრივი გადარჩევით ხორციელდება. პოპულაციაში ინვერსირების სიხშირის ცვლილება ეკოლოგიური ფაქტორების ზემოქმედებითაა გამოწვეული. სახეობის არეალში ინვერსიების მფლობელი ინდივიდები მეტწილად კონკრეტულ ტერიტორიაზე გვხვდება ზოგჯერ კი მთელ ტერიტორიას მოიცავს. არცერთი ინვერსიული ცვლილება ჰომოზიგოტებისათვის არ არის ლეტალური. ნ. დუბინინმა თანამშრომლებთან ერთად გამოავლინა, რომ დროზოფილას (D. ფუნელბის) ქალაქის (მოსკოვი, თბილისი, ბაქო) პოპულაციებს მოეპოვებოდა მეტი ინვერსიები, ვიდრე სოფლის ფორმებს. ასე მაგალითად, მოსკოვის პოპულაციებში ინვერსიების ჯამური სიხშირე 89%25-ს შეადგენდა, ხოლო გარეუბნებში 1-4%25-ს არ აღემატებოდა.
დუბინინმა და მისმა თანამშრომლებმა ლაბორატორიაში ინტროდუცირებული ხელოვნური პოპულაცია, რომელიც ფლობდა ქალაქის პოპულაციებისათვის ნიშანდობლივ ინვერსიებს ზაფხულის დასაწყისში სოფელში (მოსკოვიდან 105კმ-ის დაშორებით 105 ჰომოზიგოტი ინდივიდი) გაუშვეს. სამი წლის განმავლობაში იმეორებდნენ ცდას და მწერებში აღრიცხავდნენ ინვერსიების რაოდენობას. ყოველწლიურად ერთი და იგივე კანონზომიერება მეორდებოდა. ივნისში ინვერსიების სიხშირე დაახლოებით 50%25-მდე მცირდება, აგვისტოში 20%25-მდე, ხოლო ოქტომბერში 5-10%25-მდე ქვეითდება. გამოზამთრების შემდეგ ინვერსიების მფლობელი ჰეტეროზიგოტი ინდივიდების რაოდენობა პოპულაციაში 2-4%25-ს შეადგენდა ე.ი. ქრომოსომული პოლიმორფიზმი სოფლის რასებისათვის ნიშნულ დონეს უბრუნდებოდა. სადღეისოდ დაუდგენელია, კონკრეტულად რომელი ეკოლოგიური ფაქტორი განსაზღვრავს ქალაქისა და სოფლის რასების ამდაგვარ განსხვავებას.

სურ. 9.4. ქრომოსომული მუტაციების (ინვერსიები: ST, AR, PP,CH და SL) სიხშირე drozofilas D. pseudoobscura სხვადასხვა პოპულაციაში (ფ. დობჟანსკი, 1975).
ფ. დობჟანსკი და თანაავტორები ამერიკის საკმაოდ ვრცელ ტერიტორიაზე გავრცელებულ დროზოფილაში (D. pseudoobscura) იკვლევდნენ მე-3 ქრომოსომაში ლოკალიზებულ სხვადასხვა სახის პარაცენტრული ინვერსიების პოლიმორფიზმს. მათ მიერ გამოვლენილია მკაფიოდ გამოხატული გეოგრაფიული გრადიენტი (კლინური ცვალებადობა). შთ ინვერსია (გენთა სტანდარტული განლაგება) მაღალი სიხშირით - 50%25, აღენიშნებოდა წყნარი ოკეანის სანაპირო ზოლში (კალიფორნია) გავრცელებულ ფორმებს. მათი სიხშირე თანდათანობით მცირდებოდა სამხრეთ-აღმოსავლეთით. შთ ინვერსიებს მინიმალური სიხშირით ნიუ-მეხიკოსა და ტეხასში გავრცელებული პოპულაციები ფლობდნენ (იხ. სურ. 9.4).
AR ტიპის ინვერსიები მაღალი სიხშირით აღენიშნებოდა არიზონასა და ნიუ-მექსიკოს რასებში. ამ შტატებიდან როგორც აღმოსავლეთით, ისე დასავლეთით გავრცელებულ პოპულაციებში AR ინვერსიის სიხშირე მკვეთრად ქვეითდება. სრულიად განსხვავებული სივრცობრივი გავრცელება ახასიათებდა PP ტიპის ინვერსიებს. იგი საერთოდ არ მოეპოვებათ დასავლეთ შტატებში გავრცელებულ პოპულაციებს, ხოლო სამხრეთ შტატებში გავრცელებულ რასებში მათი სიხშირე 70-80%25-ს აღწევდა. ორი სხვა სახის CH და TL ინვერსიას კლინარული ცვალებადობა არ ახასიათებდა. შესწავლილ ტერიტორიაზე მეტ-ნაკლებად ერთნაირი, დაბალი სიხშირით იყო გავრცელებული. მიღებული შედეგები გენომის ცალკეული ნაწილის (კონკრეტულ შემთხვევაში მე-3 ქრომოსომის ინვერსირებული უბნების) ნატიფ დიფერენცირებულობასა და კონკრეტულ საარსებო პირობებში მათ განსხვავებულ სელექციურ ღირებულებაზე მიუთითებს.
დროზოფილას პოპულაციებში ინვერსიათა სიხშირე ვარირებდა როგორც ჰორიზონტალური, ისე ვერტიკალური (ზღვის დონიდან სხვადასხვა სიმაღლეზე) ზონალობის მიხედვით. სანიმუშო მაგალითად სახელდება სიერა-ნევადას მთიანეთში (კალიფორნია, აშშ) გავრცელებულ დროზოფილას პოპულაციაში ინვერსირების სიხშირის შესწავლის შედეგები. გამოირკვა, რომ სიმაღლის მატების შესაბამისად შთ ინვერსიების კონცენტრაცია პოპულაციაში მცირდებოდა 46%25-დან (250მ. ზღვის დონიდან) 10%25-მდე (3000 მ. ზღვის დონიდან), ხოლო AC ინვერსიების კონცენტრაცია მატულობდა 25%25-დან - 50%25-მდე. მაშინ როდესაც CH ინვერსიების სიხშირე უმნიშვნელოდ 16-20%25-ის ფარგლებში ფლუქტუირდება (იხ. სურ. 9.5). ინვერსიების ვარიაბილობის გამომწვევ ძირითად მიზეზს ეკოლოგიური ფაქტორის - ტემპერატურის მკვეთრი მერყეობა წარმოადგენს.

სურ. 9.5. დროზოფილაში ინვესიების (შთ, AC, CN) სიხშირის დამოკიდებულება ზღვის დონიდან სხვადასხვა სიმაღლეზე (სან-ჯასინტო, სიერა-ნევადა, აშშ) (ფ. დობჟანსკი, 1951). კვადრატით აღნიშნული ინვესიების შეფარდებითი სიხშირე: დაშტრიხულით - ST, წერტილებიანი - AR, შავით - CN.
დროზოფილას პოპულაციებში ინვერსიების სიხშირე იცვლებოდა არა მარტო სივრცულად, არამედ დროშიც (სეზონების მიხედვით). საილუსტრაციოდ მოვიყვანთ ერთ მაგალითს. მოსკოვის დროზოფილას პოპულაციაში ინვერსიების სიხშირე მარტიდან მოყოლებული სექტემბრამდე თანდათანობით 13%25-დან 51%25-მდე იზრდებოდა. გამოზამთრების შემდეგ ინვერსიების სიხშირე პოპულაციაში ნიშნულ დონეს უბრუნდებოდა. გაზაფხულიდან ციკლი კვლავ მეორდებოდა. დროზოფილაში ინვერსიების ციკლური მერყეობა იმავე კანონზომიერებას ემყარება, რაც გამოვლენილია ჩვენ მიერ განხილულ ჭიამაიების ბერლინის პოპულაციებში, კერძოდ სხვადასხვა გენეოტიპური კლასების განსხვავებული ადაპტური ღირებულება. პოპულაციის შემადგენელი განსხვავებული გენოტიპური კლასები ცვლადი ეკოლოგიური ფაქტორებისადმი (ტემპერატურა, ტენიანობა და მისთ) სპეციფიკურად არიან სპეციალიზირებული.
ამრიგად, ბუნებრივი გადარჩევის მოქმედებით პოპულაციაში მიმდინარეობს გენთა ერთიანი ბლოკების შემცველი სპეციფიკური ინვერსიების ფიქსაცია და გავრცელება. ამ ტიპის ინვერსიები ფლობენ გაცილებით მაღალ ადაპტურ ღირებულებას სტანდარტულთან შედარებით. გამოირკვა, რომ კონკრეტულ გარემო პირობებში ადაპტური ღირებულება კონკრეტულ ჰომოკარიოტიპს (ინვერსიების მიხედვით ჰომოზიგოტი) აქვს. ამასთანავე ჰეტეროკარიოტიპები (ინვერსიების მიხედვით ჰეტეროზიგოტები) გაცილებით მაღალი ადაპტური ღირებულებით გამოირჩევიან ნებისმიერი სახის ჰომოკარიოტიპებთან შედარებით. ბუნებრივი გადარჩევის ზეწოლა განაპირობებს ინვერსიული პოლიმორფიზმის განმტკიცებასა და შენარჩუნებას პოპულაციაში.
პირველად დროზოფილას ხელოვნურ პოპულაციაზე ჩატარებული გამოკვლევებით დადასტურდა, რომ გადარჩევა მიმდინარეობს ჰეტეროკარიოტიპების სასარგებლოდ, რითაც შენარჩუნებულია ინვერსიული პოლიმორფიზმი. კლასიკური ნიმუში გამოავლინა ფ. დობჟანსკიმ თანაავტორებთან ერთად. დროზოფილას საწყის ხელოვნურ პოპულაციაში AR ინვერსიული გენოტიპური კლასის რაოდენობა 4-ჯერ (80%25) ჭარბობდა CH (20%25) კლასის რაოდენობას. თერთმეტი თაობის მანძილზე ბუნებრივი გადარჩევის ზეწოლით მიმდინარეობდა ინვერსიული გენოტიპური კლასების სიხშირის ცვლილება. კერძოდ, AR გენოტიპური კლასის სიხშირე თანდათანობით ქვეითდებოდა, ხოლო CH კლასის კონცენტრაცია მატულობდა. მას შემდეგ რაც ამ ორ გენოკარიოტულ კლასს შორის რაოდენობრივი თანაფარდობა 2:1 გაუტოლდა, დამყარდა სტაბილური მდგომარეობა, რომელიც მომდევნო 40 თაობის მანძილზე შენარჩუნდა. ჩამოყალიბებულ პოპულაციაში ჰეტეროკარიოტიპების სიხშირე ჭარბობდა ჰომოკარიოტიპებისას. სპეციალური გამოკვლევებით დადასტურდა, რომ პოპულაციაში კლასთა შორის წონასწორობის დამყარება ბუნებრივი გადარჩევით იყო გამოწვეული. ამასთანავე ჰეტეროკარიოტების გადარჩენადობა მნიშვნელოვნად აღემატებოდა დანარჩენი კლასების ანალოგიურ მაჩვენებელს. შედარებითი ადაპტური ღირებულების კოეფიციენტი სამივე გენოტიპური კლასისათვის შემდეგი სახის აღმოჩნდა:
AR/AR |
AR/CH |
CU/CU |
ამრიგად, ქრომოსომული ინვერსიებით განსხვავებულ კლასებს შორის ჩამოყალიბდა ბალანსირებული პოლიმორფიზმი. ინვერსიული პოლიმორფიზმის ადაპტური როლი დადასტურებულია მრავალი მკვლევარის მიერ. მიღებული შედეგების საფუძველზე ფ. დობჟანსკიმ წამოაყენა მოსაზრება, რომ ბუნებრივი გადარჩევის პროცესში პოპულაციის შემადგენელი გენეტიკური ელემენტებით იქმნება გენთა კოადაპტური ახალი კომპლექსი. კოადაპტაცია არის გენეტიკურ სტრუქტურათა ჩამოყალიბების პროცესი, რომელშიც გენეტიკური ელემენტები ოპტიმალურად არიან ერთიმეორესთან შეთანაწყობილი. მაშასადამე გენთა კოადაპტურ კომპლექსში ალელები იმგვარად არის შერჩეული და განლაგებული, რომ მათი ურთიერთქმედებით ჰეტეროზისის ეფექტი მაქსიმალურად გამოვლინდეს.
ექსპერიმენტებით დადასტურებულია, რომ ერთსა და იმავე საკვები რესურსების ათვისებისას პოლიმორფული პოპულაციები წარმოქმნიან დიდ ბიომასას და მეტ რიცხოვნობას, ვიდრე მონომორფულები, ე.ი. ისინი გარემო რესურსების ეკონომიური გამოყენებით გამოირჩევიან.
ამჟამად ინვერსიული პოლიმორფიზმი დროზოფილას ასზე მეტ სახეობაშია გამოვლენილი. ამასთანავე ცალკეულ სახეობაზე საშუალოდ 6 სხვადასხვა სახის ინვერსია მოდის. მხოლოდ რამდენიმე სახეობა (D. simulans, D. virilis, D. repleta) აღმოჩნდა მონომორფული. ასეთი განსხვავების მიზეზი სადღეისოდ უცნობია. მაშასადამე, დიდი მნიშვნელობის მიუხედავად ეს მოვლენა თვით, Drosophilidae-ს ოჯახისთვისაც კი არ აღმოჩნდა უნივერსალური ხასიათის. ასევე ინვერსიული პოლიმორფიზმი ორფრთიანთა რიგის სხვა ოჯახებსაც (Simuliidae, Chironomidae, Culicidae) აღენიშნებათ.
სადღეისოდ ინვერსიული პოლიმორფიზმი გამოვლენილია ხერხემლიან ცხოველებშიც, კერძოდ, პერიცენტრული პოპულაციური პოლიმორფიზმი ფართოდაა გავრცელებული ზაზუნებსა და მექვიშიებში. კოლორადოში (აშშ) გავრცელებულ ზაზუნას (Peromyscus maniculatus) მე-4 ქრომოსომაში ლოკალიზებული პერიცენტრული ინვერსიის სიხშირე ვარირებდა ვერტიკალური ზონალობის შესაბამისად. კერძოდ, ზღვის დონიდან სიმაღლის მატების კვალობაზე იზრდებოდა ინვერსიების კონცენტრაცია პოპულაციაში.
იაპონიაში გავრცელებულ შავ ვირთაგვას (Rattus rattus) პოპულაციებში გამოვლენილია ინვერსიათა კლინური ცვალებადობა. მათ კარიოტიპის (2ნ=42) პირველი წყვილი ქრომოსომა პოლიმორფული აღმოაჩნდათ. იგი შესაძლოა იყოს აკროცენტრული (A) ან სუბტელოცენტრული (C). ეს უკანასკნელი აკროცენტრული ქრომოსომის პერიცენტრული ინვერსიის შედეგადაა ჩამოყალიბებული. იაპონიის ჩრდილო-აღმოსავლეთ ნაწილში გავრცელებული ვირთაგვები კარიოტიპული კლასების (A/A; A/C; C/C) სიხშირით მკვეთრად განსხვავდებიან სამხრეთ-დასავლეთით გავრცელებული ფორმებისგან (იხ. სურ. 9.6). პოპულაციები სივრცობრივი ბარიერით (თოვლის საფარი) არიან გამიჯნულნი, რის გამოც გენთა ნაკადი (მიგრაციები) არ მიმდინარეობს.
სურ. 9.6. იაპონიის კუნძულზე გავრცელებულ შავი ვირთაგვას (R. rattus) პოპულაციებში სამი კარიომორფის გავრცელების სიხშირე (ლ. კაიდანოვი, 1996).
პერიცენტრული ქრომოსომული პოლიმორფიზმი (1,8 და 13 წყვილი აუტოსომა) გამოვლენილია აზიაში (იაპონია, ჰონკონგი, ფილიპინები, ტაილანდი) გავრცელებულ რუხ ვირთაგვაში (R. norvegicus). ამავე ტიპის ინვერსიული პოლიმორფიზმი (3-6 აუტოსომა) რეგისტრირებულია რუსეთსა (ციმბირი, ალტაი) და ყაზახისტანში გავრცელებულ ცოკორში (Myospalax myospalax).
|
|
11.2.2 რობერტსონული ტრანსლოკაციების პოლიმორფიზმი |
▲ზევით დაბრუნება |
XX ს. 60-იან წლებამდე სახეობისთვის ნიშანდობლივი ქრომოსომათა რაოდენობა და მორფოლოგია (კარიოტიპი) უცვლელ ნიშნად განიხილებოდა. სახეობის ინდენტიფიკაციაში კარიოტიპი სახეობის არსებით ტაქსონად იყო მიჩნეული. მოგვიანებით დაგროვდა ფაქტები, რომელიც მიუთითებდა, რომ ამ ნიშნით განსხვდებიან არა მარტო სხვადასხვა პოპულაციები და გეოგრაფიული რასები, არამედ ერთი და იმავე პოპულაციის შემადგენელი წევრები.
ამჟამად ცნობილია, რომ მხოლოდ ძუძუმწოვრებში ქრომოსომული პოლიმორფიზმის მქონე სახეობის წილი 10,9%25-ს შეადგენს. ხერხემლიან ცხოველთა თითქმის ყველა კლასში გვხვდება რობერტსონული ტრასლოკაციების მიხედვით პოლიმორფული პოპულაციები. რობერტსონული ტრანსლოკაციისას, ორი აკროცენტრული ქრომოსომის ურთიერთდაკავშირებით ერთი მეტაცენტრული ქრომოსომა ყალიბდება, ან პირუკუ მეტაცენტრული ქრომოსომა დისოცირდება ორ აკროცენტრულ ქრომოსომად. ეს მოვლენა პირველად 1916წ. აღმოაჩინა უ.რობერტსონმა სწორფრთებიანებში. ამ ფენომენის ფართომასშტაბიანი კვლევა მხოლოდ ნახევარი საუკუნის შემდეგ დაიწყეს.
იაპონელმა ციტოგენეტიკოსმა ს. ონომ თანაავტორებთან ერთად პირველად თევზებში გამოავლინა რობერტსონული ტრანსლოკაციებით განპირობებული პოლიმორფიზმი. კერძოდ, ფოლადთავა ორაგულის (შალმო გაირდნერი) პოპულაციის წევრებში კარიოტიპი (2N) 58-დან 64-მდე ქრომოსომის ფარგლებში მერყეობდა. ამასთანავე ყველა მათგანში ქრომოსომთა ფუნდამენტური რაოდენობა (NF=104) მუდმივი იყო. პოლიმორფიზმს კარიოტიპში წარმოდგენილი 40 მეტა - და სუბმეტაცენტრული ინვარიანტული ქრომოსომის დანარჩენებთან რობერტსონული დაკავშირება იწვევდა. რობერტსონული პერეტრუბაციის გამო პოპულაციაში განსხვავებული დიპლოიდური კომპლექტის მფლობელი კარიომორფები ყალიბდებოდა.
წყნარ ოკეანეში გავრცელებულ ორაგულის ორ სახეობაში ნერკასა (Oncorhynchus nerka) და კიჟუჩას (O. kisutsch) პოლიმორფიზმი ვიწრო ფარგლებში აღენიშნებათ. კარიომორფა (2N) შესაბამისად 56-58 და 58-60 ფარგლებში ვარირებს (NF=104).
მკვეთრად გამოხატული რობერტსონული ტიპის პოლიმორფულობა ნიშანდობლივია თაგვისებრი მღრღნელებისათვის. ზოგიერთ მათგანს აღენიშნება ე.წ. რობერტსონული „მარაოს“ ფენომენი. კერძოდ კარიოტიპში წარმოდგენილი ყველა ან თითქმის ყველა აკროცენტრული ქრომოსომა დაწყვილებულია.
პირველად ეს კანონზომიერება გამოავლინა გამოჩენილმა შვეიცარიელმა კარიოლოგმა რ. მატთეიმ აფრიკულ ჩია თაგვებში (ქვეგვარი Leggada, კომპლექსური სახეობა minutoides/musculoideს). პოპულაციის შემადგენელ წევრებს გააჩნდათ 18-დან 34-მდე ქრომოსომის შემცველი კარიომორფები (NF=36). კარიომორფებში ყველა აკროცენტრული ქრომოსომა წყვილებად იყო ასოცირებული. კარიომორფების ფართო დიფერენციაცია (რობერტსონული „მარაო“) გამოვლენილია ალპებსა (შვეიცარია) და აპენინებში (იტალია) გავრცელებულ სახლის თაგვში (Mus musculus). პოპულაციებში ქრომოსომთა დიპლოიდური კომპლექტი (2N) 22-დან 40-მდე ვარირებდა, თითოეულ კარიომორფაში ფუნდამენტური რაოდენობა (NF=40) უცვლელი აღმოჩნდა. ერთი და იგივე აკროცენტრული ქრომოსომა ნებისმიერ სხვა აკროცენტრულთან არის ასოცირებული.
მიწისმთხრელ მღრღნელ ბრუცუნაში (Wutala, Ellobius talpinus) უნიკალური რობერტსონული „მარაო“ გამოავლინეს პროფესორებმა ნ. ვორონცოვმა და ე. ლიაპუნოვამ. ამ ცხოველს საკმაოდ დიდი არეალი უკავია. იგი გავრცელებულია უკრაინიდან მოყოლებული მანჯურიის ჩათვლით. მთელ ტერიტორიაზე მხოლოდ ორი სტაბილური კარიომორფა გვხვდება, გამონაკლისია პამირ-ალტაის მცირე ტერიტორია, კერძოდ, მდინარე ვახშ-სურხობის ველი, სადაც პოპულაციაში კარიომორფები სრულად გამოხატული რობერტსონული მარაოთი არის წარმოდგენილი. მასში 24 სახის კარიომორფა ყველა შესაძლო დიპლოიდურ კომპლექტს (2N=31-დან ვიდრე 2N=54) ფლობს. ამასთანავე თითოეულ მათგანს სტაბილურად შენარჩუნებული აქვთ ქრომოსომთა ფუნდამენტური რაოდენობა (NF=56). კარიომორფათა უმეტესობა ჰიბრიდული წარმომავლობის აღმოჩნდა, რომლებიც ამ ნიშნით განსხვავებული მცირერიცხოვანი ჯგუფებიდან იღებს სათავეს. რობერტსონული მარაო ნიშანდობლივია კავკასიაში გავრცელებულ ბუჩქნარის მემინდვრიას გვარში (Pitymus) გაერთიანებული სახეობებისათვის. რობერტსონული ტრანსლოკაციების პოლიმორფიზმი მიწის მთხრელ მღრღნელებში (ბრუცა, ბრუცუნა, გოფერი, ცოკორი, ტუკო-ტუკო) შესწავლილ სახეობების პოპულაციებში 40-50%25-ს შეადგენს. პოლიმორფიზმის ეს ფორმა დამახასიათებელია ჩლიქოსნებისათვის (ირმები, ანტილოპები, გარეული ღორები, რქაფიწალები). ტრანსლოკაციური პოლიმორფიზმის ჩამოყალიბებისა და პოპულაციებში მისი შენარჩუნების ფაქტი, მის ადაპტურ მნიშვნელობაზე მიუთითებს.
|
|
11.2.3 B - ქრომოსომების პოლიმორფიზმი |
▲ზევით დაბრუნება |
მცენარეთა და ცხოველთა მრავალ სახეობაში გამოვლენილია B - ქრომოსომის პოლიმორფიზმი. ძირითადი კომპლექტის შემადგენელ ქრომოსომების (A ქრომოსომები) გარდა კარიოტიპს ზოგჯერ მოეპოვება დამატებითი ანუ ზეკომპლექტური ე.წ. B - ქრომოსომები. ისინი ძირითადად ჰეტეროქრომატიზირებულია. ამ სახის გენომური პოლიმორფიზმი გვხვდება მწერებში. მაგალითად 19 სხვადასხვა სახეობის ქინქლას (Simulidae) პოპულაციებში რეგულარულად მნიშვნელოვანი სიხშირით (10 - 15%25) გვხვდება B - ქრომოსომების მფლობელი ინდივიდები. ქინქლა ადაგმიაში (Adagmia ornata) B - ქრომოსომების რაოდენობა 1-დან 8-მდე მერყეობს. ამ ქრომოსომის სიხშირე პოპულაციაში გარემოს გაჭუჭყიანების ხარისხის მიხედვით ვარირებს, რაც მის ადაპტურ მნიშვნელობაზე მიუთითებს.
B - ქრომოსომების პოლიმორფიზმი გამოვლენილია უმდაბლეს ხერხემლიანთა სხვადასხვა ჯგუფებში, კერძოდ, თევზებში (ძირითად სიგანაირებში) და ხვლიკებში. ისინი ფართოდ არის წარმოდგენილი ფრინველთა კარიოტიპში. ძუძუმწოვრების 20-ზე მეტი სახეობა ამ ნიშნით პოლიმორფულია. მათ კარიოტიპებში სხვადასხვა რაოდენობით მოეპოვებათ B - ქრომოსომები. ამასთანავე ქრომოსომათა რაოდენობა ვარიაბილურია როგორც პოპულაციის სხვადასხვა წევრებში, ისე ერთი და იმავე ინდივიდის ცალკეულ უჯრედებშიც (მოზაიციზმი).
აღმოსავლეთ აზიაში გავრცელებულ ტყის თაგვსა (Apodemus peninsulae) და მელას (Vulpes vulpes) თითქმის ყოველ ინდივიდს გააჩნია B - ქრომოსომა. ტყის თაგვში მათი რაოდენობა 0-დან 24-მდე ვარირებს, მელაში კი 1-დან 8-მდე. ამ უკანასკნელში პოლიმორფიზმი ზოგადი ხასიათისაა და ნიშანდობლივია მთელ არეალზე გავრცელებული პოპულაციებისთვის. თითქმის ყველა ინდივიდში შეინიშნება მოზაიციზმი (სხვადასხვა უჯრედი განსხვავებული რაოდენობით დამატებით ქრომოსომებს ფლობს).
ფარულთესლოვან მცენარეთა 750 სახეობაში გამოვლენილია B - ქრომოსომები. მათი რაოდენობა 1-35-მდე ფარგლებში ვარირებს. გენომური პოლიმორფიზმის ამ ფორმას მცენარეებში ადაპტური მნიშვნელობა გააჩნია. რასაც შემდეგი მაგალითიც ადასტურებს. მდელოს წივანას (Festuca pratensis) გენომური პოლიმორფიზმის მფლობელი პოპულაციები მძიმე თიხნარ ნიადაგზე სახლობენ, ხოლო ღიღილოს (Centaurea scabiosa) პოპულაციები მშრალ ადგილებში. ნაძვის (Picea glauca) B - ქრომოსომის მქონე თესლები გაცილებით სწრაფად ღივდებიან და აღმოცენდებიან, მათთან შედარებით, რომელთაც იგი არ მოეპოვებათ.
|
|
11.2.4 სასქესო ქრომოსომების პოლიმორფიზმი |
▲ზევით დაბრუნება |
ცალსქესიან ცხოველებში გამოვლენილია გონადოსომების (სასქესო ქრომოსომები) ფორმისა და რაოდენობის პოლიმორფიზმი. სქესის განსაზღვრის ტიპური XX-XY ფორმის გარდა გვხვდება მრავლობითი სასქესო ქრომოსომების ტიპი. მაგალითად, თევზებში გამოვლენილია მრავლობითი სასქესო ქრომოსომებით სქესის განსაზღვის 10 განსხვავებული ფორმა. ზოგიერთ ძუძუმწოვარში გვხვდება ზემოთ აღნიშნული სქესის განსაზღვრის ტიპი. იგი ყალიბდება გონადოსომის აუტოსომასთან ან მის ფრაგმენტთან ტრანსლოკაციური შეერთებით. X - გონადოსომაზე აუტოსომის ტრანსლოკაცია იწვევს მამრებში დამატებით არაჰომოლოგიურ გონადოსომის: XY1Y2 ჩამოყალიბებას. ტრანსლოცირებული Y2 ქრომოსომა სქესის განსაზღვრაში არ მონაწილეობს. მსგავსი მოვლენა აღწერილია ძუძუმწოვრების 6 რიგში გაერთიანებულ 40 სახეობაში.
როდესაც აუტოსომა Y გონადოსომაზეა ტრანსლოკალიზებული მაშინ მამრს სამი სასქესო ქრომოსომა: X1X2 გააჩნია. X1 - ტიპური გონადოსომაა, ხოლო X2 - გონადოსომა აუტოსომისა და Y ქრომოსომის შეერთების შედეგად არის ჩამოყალიბებული. ამდაგვარი კარიოტიპის მფლობელი მამრები გამოვლენილია ძუძუმწოვართა 18 სახეობაში, რომლებიც 6 სხვადასხვა რიგს მიეკუთვნება.
სქესის განსაზღვრის სრულიად ახალი ტიპი გამოვლენილია ლემინგებში (Dicrostonyx torquatus). მდედრი სამი კარიომორფით: XmXm; XmXf; Xf0 გვხვდება, მამრები მხოლოდ ერთით - Xm0 (Y გონადოსომას ისინი არ ფლობენ). ყველა ზემოთ განხილულ შემთხვევაში სასქესო ქრომოსომების პოლიმორფულობის მიუხედავად სქესთა თანაფარდობა არის დახლოებით 1:1.
|
|
11.2.5 ჰეტეროქრომატინის პოლიმორფიზმი |
▲ზევით დაბრუნება |
ქრომოსომების დიფერენცირებული შეღებვის მეთოდების გამოყენებით შესაძლებელი გახდა სომატურ და სასქესო ქრომოსომებში ჰეტეროქრომატინის შეფარდებითი ცვალებადობის გამოვლენა და შესწავლა. ამერიკულ ზაზუნებში (Peromyscus) C - ქრომატინის (ლოკალიზებულია ცენტრომერასთან) მოკლე მხრის ზომა ვარირებს როგორც სახეობის ისე პოპულაციის დონეზე. შავ ვირთაგვაში C - ჰეტეროქრომატინის სპეციფიკური განლაგება ჰიბრიდულ შთამომავლებს მენდელისეული კანონზომიერებით გადაეცემოდა. მღრღნელებს, ჩლიქიანებს, ვეშაპისნაირებს და რიგ სხვა ძუძუმწოვრებს, როგორც ცენტრომერიასთან, ისე ტელომერულ უბანთან განლაგებული ჰეტეროქრომატინი ფორმითა და ზომით პოლიმორფული აღმოაჩნდათ. ყველაზე სრულყოფილად ჰეტეროქრომატინული პოლიმორფიზმი შესწავლილია ადამიანში. ამ ნიშნით მნიშვნელოვანი ვარიაბილობა აღენიშნებათ 1, 9, 16 აუტოსომურ და Y - სასქესო ქრომოსომას. C - და Q ჰეტეროქრომატინის შემცველი ბლოკების განლაგება ადამიანის ქრომოსომებში მკაფიოდ ინდივიდურობას ავლენს.
გამოჩენილი ციტოგენეტიკოსის ა. პროკოფიევა-ბელგოვსკაიას შეხედულებით ქრომოსომის ჰეტეროქრომატინულ უბნებში ჰეტეროქრომატინის ცვლილება სახეობისათვის ადაპტური მნიშვნელობისაა. მუტაციური პროცესისაგან განსხვავებით იგი უზრუნველყოფს შეცვლილი გარემოსადმი ორგანიზმის სწრაფ ადაპტაციას. ქრომოსომის სტრუქტურაში ჰეტეროქრომატინული უბნების ფიქსაცია მისმა მრავალმა სასარგებლო თვისებამ განაპირობა, რომლის რეალიზაცია ხორციელდება როგორც ორგანიზმის ისე პოპულაციის დონეზე.
|
|
11.3 9.3. ბიოქიმიური პოლიმორფიზმი |
▲ზევით დაბრუნება |
გასული საუკუნის 60-იან წლებში პულს-ელექტროფორეზის მეთოდის გამოყენებით დაიწყეს გენეტიკურად კონტროლირებადი ცილების პოლიმორფიზმის შესწავლა. პოპულაციის გენეტიკაში ამ მეთოდის დანერგვამ ძირეული გარდატეხა მოახდინა. მკვეთრად გაიზარდა პოპულაციაში გენეტიკური ჰეტეროგენულობის გამოვლენის შესაძლებლობა. გენეტიკურ მარკერებად წესისამებრ ალოზიმებს იყენებენ. მათ სინთეზს კონკრეტული ალელები აკონტროლებენ. პოპულაციის გენეტიკაში შემოიტანეს ახალი პარამეტრი: საშუალო ჰეტეროზიგოტულობა და პოლიმორფულობა (იხ. §4.).
|
|
11.3.1 დიდ ტაქსონებში ცილათა პოლიმორფიზმის დონეები |
▲ზევით დაბრუნება |
ჩატარებული გამოკვლევებით გაირკვა, რომ დიდ ტაქსონებში გაერთიანებულ სახეობათა ცილების ცვალებადობის დონე საკმაოდ მაღალია. მცენარეთა და ცხოველთა სამეფოსთვის განსაზღვრულია ცილების საშუალო პოლიმორფულობა და ჰეტეროზიგოტულობა (იხ. ცხრ. 9.1). მათში სტრუქტურული გენების დაახლოებით 30%25 პოლიმორფულია ე.ი. წარმოდგენილია ერთზე მეტი ალელით. კონკრეტულ ინდივიდს საშუალოდ გენების 7%25 ჰეტეროზიგოტულ მდგომარეობაშია. სხვადასხვა დიდი ტაქსონომური ჯგუფები საკმაოდ განსხვავდებიან ცილების პოლიმორფიზმით. ცილების მაკონტროლებელი ლოკუსების მაღალი ვარიაბილობა აღენიშნება უხერხემლო ცხოველებს. მათთან შედარებით ხერხემლიანებში ლოკუსთა ცვალებადობა 1.5-2-ჯერ დაბალია. თვითმტვერია მცენარეებთან შედარებით ჯვარედინამტვრიანებს 2-3-ჯერ მაღალი ლოკუსთა ცვალებადობა აღენიშნებათ. საშუალო ჰეტეროზიგოტულობა საკმაოდ მაღალია ადამიანის პოპულაციებში, რომელიც 6.7%25-ს შეადგენს.
|
|
11.3.2 პოპულაციები: მსგავსება და განსხვავება. კლინური ცვალებადობა |
▲ზევით დაბრუნება |
თავდაპირველად 1966 წელს ინგლისში გ. ჰარისონის და თანაავტორების მიერ ადამიანში, ხოლო ამერიკაში რ. ლევონტინისა და მისი თანამშრომლების მიერ დროზოფილაში ჩატარებული გამოკვლევებით თვალნათლივ დადასტურდა, რომ პოპულაციები ფლობენ ცილების პოლიმორფიზმის მაღალ დონეს. ადამიანის 10 შემთხვევით შერჩეული ფერმენტიდან 3 პოლიმორფული აღმოჩნდა.
ცხრილი. 9.1.
ცილების მაკონტროლებელ ლოკუსთა საშუალო
ჰეტეროზიგოტულობა (H) და პოლიმორფიზმი (P)
დიდ ტაქსონებში (ე.ნევო და თანაავტ., 1984)
ტაქსონები |
სახეობათა |
H |
სახეობათა |
P |
ხერხემლიანები |
551 |
0.054 |
596 |
0.226 |
დროზოფილას (D. pseudoobscura) ბუნებრივი პოპულაციის ლარვების ჰემოლიმფაში არსებული 8 ფერმენტული და 10 არაფერმენტული ცილიდან 1/3 პოლიმორფული იყო.
განსხვავებული პოპულაციები ცილების პოლიმორფიზმის შენარჩუნების უნარით ერთიმეორეს ემსგავსებიან. ეს ფენომენი პირველად ლეონტინმა და მისმა თანამშრომლებმა დაადგინეს. კალიფორნიის, ტეხასისა და კოლორადოს დროზოფილას განცალკევებულ პოპულაციებში შესწავლილი 20 ლოკუსიდან მხოლოდ 11 აღმოჩნდა მონომორფული. გეოგრაფიულად და ეკოლოგიურად იზოლირებული პოპულაციები პოლიმორფული ალელების სიხშირით ერთმანეთისგან უმნიშვნელოდ განსხვავდებოდნენ.
მსგავსი ფენომენი გამოვლენილია დროზოფილას სხვა სახეობაში (D. willistoni). კარიბის ზღვის კუნძულებსა და კონტინენტზე (ცენტრალური და სამხრეთ ამერიკა) გავრცელებულ იზოლირებულ პოპულაციებში შესწავლილი 20 გენიდან მხოლოდ 4 აღმოჩნდა მონომორფული. საკმაოდ ვრცელ განსხვავებულ არეალზე გავრცელებულ იზოლირებულ პოპულაციებში 14-ს პოლიმორფული ალელის სიხშირე მსგავსი აღმოაჩნდათ (აიალა, 1984).
დედამიწის ხუთ კონტინენტზე გავრცელებულ დროზოფილას (D. melanogaster) პოპულაციებში შესწავლილია 26 სტრუქტურული გენით კოდირებული ალოზიმების პოლიმორფულობა. გაანალიზებულ 90-ვე პოპულაციას ერთი და იგივე ორი ლოკუსი მონომორფული გააჩნდათ. ვრცელ ტერიტორიაზე გავრცელებულ პოპულაციებში 12 პოლიმორფული ლოკუსის სიხშირე ერთნაირი იყო. დანარჩენი ლოკუსების სიხშირე კლიმატურ ფაქტორებზე იყო დამოკიდებული. ერთი ალელი (ADH-F) კლინურ ცვალებადობას ავლენდა. კერძოდ, სამხრეთ ამერიკის აღმოსავლეთ სანაპიროზე გავრცელებულ პოპულაციებში სამხრეთიდან ჩრდილოეთის მიმართულებით პოლიმორფულ ალელთა სიხშირე წრფივად მატულობდა. დანარჩენ კონტინენტზე რთული სურათი შეინიშნებოდა. რაოდენობრივად განსხვავდებოდა ტროპიკები და სუბტროპიკები ცივი რეგიონებისგან.
კლინური ცვალებადობის სანიმუშო მაგალითი გამოვლენილია თევზ კლარკში (Catostomus clarki), რომელიც მდინარე კოლორადოს აუზში ბინადრობს. მასში შეისწავლეს ცილის შრატის ესთერაზას მაკონტროლირებელი ალელების სიხშირე. 800 კმ-ის სიგრძეზე ჩრდილოეთიდან სამხრეთის მიმართულებით ეს მაჩვენებელი კანონზომიერად იცვლებოდა. მდინარის აუზის ჩრდილოეთ ნაწილში ბ ალელის სიხშირე საკმაოდ მაღალია. ალელის კონცენტრაცია პოპულაციებში 0.82-ის ტოლია. სამხრეთისაკენ ეს მაჩვენებელი თანდათანობით - წრფივად კლებულობს და სამხრეთის პოპულაციებში მისი სიხშირე ნულს უტოლდება. სამხრეთის პოპულაციებს მეორე ა ალელი მოეპოვება. ამრიგად სამხრეთის პოპულაციები ფლობენ A/A ჰომოზიგოტ კლასებს აუზის შუა წელში გავრცელებულ პოპულაციებში გვხვდება A/B ჰეტეროზიგოტი ფორმები. ჩრდილოეთ ნაწილში პოპულაციები ძირითადად B/B ჰომოზიგოტი გენოტიპური კლასებითაა წარმოდგენილი. გამოირკვა, რომ მაღალ ტემპერატურაზე (370ჩ) A/A გენოტიპის თევზებში ესთერიაზას აქტიობა 10-ჯერ მეტია ვიდრე B/B გენოტიპის მფლობელებში.
სურ. 9.7. აშშ-ს სამხრეთ-აღმოსავლეთ რეგიონში გავრცელებულ თეთრფეხა ზაზუნას (Peromyscus polionotus) პოპულაციების საშუალო ჰეტეროგენულობა (ლ. კაიდანოვი, 1996). 1-30 პოპულაციის ნომერი; წრეში შავი სექტორი - საშუალო ჰეტეროგენულობა ინდივიდზე.
ცილების პოლიმორფიზმის კლინური ცვალებადობა გამოვლენილია ძუძუმწოვრებშიც. საილუსტრაციოდ მხოლოდ ერთ მაგალითს მოვიყვანთ. მექსიკის ყურის სანაპიროზე (აშშ) გავრცელებულ თეთრფეხა ზაზუნას (Peromyscus polionotus) 30 პოპულაციაში შეისწავლეს 32 სტრუქტურული გენის სიხშირე. საშუალო ჰეტეროზიგოტულობის მიხედვით პოპულაციები განსხვავდებოდნენ. ნახევარკუნძულ ფლორიდაზე გავრცელებული პოპულაციები H (საშუალო ჰეტერზიგოტულობა) მაჩვენებლით 75%25 ჭარბობს კონტინენტის არეალზე გავრცელებულ ფორმებს. დაბალი ჰეტეროზიგოტულობით გამოირჩეოდნენ დასავლეთ კუნძულებზე ბინადარი პოპულაციები, ხოლო კონტინენტის სანაპირო ზოლში გავრცელებული ფორმების ალელთა სიხშირე ფართო ფარგლებში ვარირებს (იხ. სურ. 9.7).
|
|
11.3.3 სუბსტრატისადმი სპეციფიკური ცილების პოლიმორფიზმი |
▲ზევით დაბრუნება |
სხვადასხვა ბიოლოგიურ ობიექტებში ჩატარებული გამოკვლევებით დადგენილია, რომ ცილების პოლიმორფიზმი მნიშვნელოვნადაა დამოკიდებული ცილების სტრუქტურულ-ფუნქციურ თავისებურებებზე. ფართო სუბსტრატული სპეციფიკურობის ფერმენტები ჩვეულებრივ გაცილებით ცვალებადია ვიდრე ის ფერმენტები, რომლების ენდოგენური ბუნების სპეციფიკურ სუბსტრატს აკატალიზებენ. პირველი ჯგუფის ფერმენტები გარდაქმნიან გარემოდან მიღებულ სუბსტრატს. მეორე ჯგუფის ფერმენტები უჯრედში მიმდინარე მეტაბოლიზმის პროცესის რეგულაციაში მონაწილეობენ. ამ უკანასკნელს პირობითად ორ ქვეჯგუფად: მარეგულირებელ და არამარეგულირებელ ფერმენტებად ყოფენ. მარეგულირებელი ფერმენტები წესისამებრ მეტაბოლიზმის პროცესის საწყის ან საბოლოო ეტაპის რეაქციებში მონაწილეობენ. სტაბილურად ფუნქციონირებადი არამარეგულირებელი ფერმენტები მეტაბოლიზმის გზის საკვანძო ეტაპებში იღებენ მონაწილეობას. ამდენად, მათი საშუალო პოლიმორფულობა საკმაოდ დაბალია. დადგენილია დადებითი კორელაციური დამოკიდებულება სტრუქტურული გენების გენეტიკურ ცვალებადობასა და მათ მიერ კოდირებულ ცილის სივრცით კონფორმაციას შორის. გენეტიკური ცვალებადობა მკვეთრად მცირდება ცილის აგრეგატებში (მეოთხეულ სტრუქტურაში) სუბერთეულების რაოდენობის მომატების შესაბამისად. უაღრესად დაბალი სიხშირით გენეტიკური ცვლილებები ინდუცირდება და ფიქსირდება ისეთ ცილებში (კოვარიონში), რომლებიც უმნიშვნელოვანეს საბაზისო ფუნქციებს ანხორციელებენ. უაღრესად ნელა იცვლება, მიოგლობინი (კუნთის ცილები), ჰემოგლობინი, ჰისტონები, ასევე მემბრანებთან დაკავშირებული ცილები. ეს ცილები მათთვის ნიშანდობლივ კონკრეტულ მაგრამ სასიცოცხლო თვალთახედვით უმნიშვნელოვანეს ფუნქციას ასრულებენ. მაგალითად ჰისტონების სივრცითი კონფორმაციის უმნიშვნელო ცვლილებითაც კი ირღვევა ქრომატინის ნუკლეოსომური ორგანიზაცია, ვეღარ სტაბილიზდება ზექრომატინული - სოლენოიდის სტრუქტურა, დნმ-ის კომპაქტიზაცია ატიპურად მიმდინარეობს. ამ ტიპის ცვლილებით მიტოზის შემდეგ ორივე შვილეული უჯრედი იღუპება.
|
|
11.3.4 ცილების პოლიმორფიზმის ადაპტური მნიშვნელობა |
▲ზევით დაბრუნება |
სხვადასხვა გენოტიპურ კლასებს განსხვავებული სელექციური მნიშვნელობა აღენიშნება. პოპულაციებში ჩატარებული გენეტიკური ანალიზით დადგენილია, რომ განსხვავებული გენოტიპური კლასები კონკრეტულ საარსებო გარემოში არაერთგვაროვან ადაპტურ ღირებულებას ფლობენ. ცილების პულს-ელექტროფორეზით გამოვლენილია განსხვავებული ელექტრომორფები. მათზე ბუნებრივი გადარჩევის ზეწოლა არაერთმნიშვნელოვანია. უპირატესად მრავლდებიან და შთამომავლობას ტოვებენ იმ მორფების მფლობელი ინდივიდები, რომლებიც კონკრეტულ საარსებო გარემოში ადაპტურ უპირატესობას ფლობენ.
წყნარი ოკეანის ორაგულებში (გვარი Oncorhynchus), კერძოდ, ნერკასა (O. nerka) და ღურკანაში (O. gorbuscha) ჩატარებული გამოკვლევებით დადგენილია, რომ პოპულაციაში განსხვავებულ გენოტიპებს, ნაირგვარი სელექციური მნიშვნელობა გააჩნიათ. ორივე მათგანი მონოციკლური სახეობაა. წელიწადში მხოლოდ ერთხელ მრავლდებიან და ტოფობის შემდეგ მალე იღუპებიან. წყნარი ოკეანის ორაგულები გამსვლელი თევზებია. სქესმწიფე ინდივიდები ზღვიდან ათეულ კილომეტრით მიგრირებენ და იმ მდინარეს ან ტბას უბრუნდებიან, სადაც ისინი დაიჩეკნენ (სახლის ინსტინქტი). ყალიბდება მრავალრიცხოვანი ინდივიდებისგან შემდგარი ლოკალური გუნდი (ე.წ. სუბპოპულაცია). ისინი ერთიმეორისგან არიან იზოლირებული. სუბპოპულაციებს შორის დაბალი სიხშირით მიმდინარეობს გენთა ნაკადი (ემიგრაცია-იმიგრაცია). მიუხედავად ამისა, ემიგრანტის წვლილი გენოფონდის ფორმირებაში საკმაოდ დიდია. ლიფსიტები 3-4 წლამდე მდინარეში რჩებიან. შემდეგ გუნდი ზღვისკენ ეშვება. ინდივიდები 3-4 წლის განმავლობაში ზღვაში ცხოვრობენ. გვხვდება მობინადრე ფორმები, რომლებიც მთელი სიცოცხლის მანძილზე მტკნარ წყალში (მდინარეში ან ტბაში) ატარებენ. ზღვის პოპულაციაში ნორმალური ფორმების გარდა გვხვდება პატარა ზომის მამრები. ისინი სქესობრივად სწრაფად მწიფდებიან. ზღვაში ერთ, იშვიათად ორ წელს რჩებიან. ტოფობისას სელექციური უპირატესობა დიდი ზომის 5-7 წლის სქესმწიფე მამრებს გააჩნიათ. მიგრაციის დროს მათ უჭირთ მდინარის თავთხელი მონაკვეთების გადალახვა ან წესისამებრ მტაცებელთა მსხვერპლი ხდებიან. იგი წარმოადგენს ერთგვარ „საზღაურს ევოლუციისათვის“.
ნერკას პოპულაციაში შესწავლილია ლაქტატდეჰიდროგენაზას (LDH-A, LDH-B) და ფოსფოგლუკომუტაზას (PGM-A, PGM-B) მარკერული ალელების შემცველი გენოტიპების სიხშირე. მიუხედავად იმისა, რომ სისტემა სტაბილური იყო, სუბპოპულაციები გენოტიპური კლასების სიხშირით ერთმანეთისგან მნიშვნელოვნად განსხვავდებოდნენ. გენოტიპური კლასების განსხვავებული სიხშირით ჩამოყალიბებას (ჰარდი-ვანბერგის კანონიდან გადახრა) ტემპერატურული ფაქტორი იწვევს. დაბალი ტემპერატურისადმი განსაკუთრებით მგრძნობიარეა ონტოგენეზური განვითარების ადრეული სტადიები. ტემპერატურისადმი მგრძნობელობას ჰომოზიგოტური კლასები ავლენენ. დიდი ზომის მამრები ძირითადად ჰომოზიგოტურებია. ჰეტეროზიგოტულ გენოტიპურ კლასებს პატარა ზომის მამრები ფლობენ. მათში ჰეტეროზიგოტულობა კორელაციურ დამოკიდებულებაშია ნაადრევ სქესობრივ მომწიფებასა და სიცივისადმი მედეგობასთან, ასევე ზრდის ინტენსიობასთან. ასევე ჰეტეროზიგოტული გენოტიპისაა პატარა ზომის მობინადრე ფორმები.
ზღვის ფსკერზე ბინადარ (იაპონიის ზღვა, პეტრე დიდის ყურე) უხერხემლო ცხოველებში (ხამანწკები, მიედიები, ზღვის ვარსკვლავები და მისთ.) შეისწავლეს 70 მარკერი ალეზიმური ცილის მაკონტროლირებელი გენების შემცველობა. განისაზღვრა პოპულაციებში გენოტიპური კლასების სიხშირე და სივრცობრივად ჰეტეროზიგოტების განაწილება. შესწავლილი პოპულაციები ალელებისა და გენოტიპების სიხშირით ერთმანეთის მსგავსი აღმოჩნდა, პოპულაციებში დიდ გენეტიკურ ნაირგვარობას ფლობენ, რაც ლარვული სტადიის არსებობისა და მაღალი ნაყოფიერების შედეგია.
ცილების პოლიმორფიზმის ადაპტური როლი დადასტურებულია ამფიბიებში. ვეფხვა-ამბისტომაში (Ambystoma tigrinum) გამოვლენილია დადებითი კორელაცია ლარვის სხეულის ზომასა და 7 პოლიმორფული ლოკუსის გასაშუალებულ ჰეტეროზიგოტულობასთან. გომბეშოში (Bufo boreas) თავკომბალას ნელი ან სწრაფი განვითარება დამოკიდებული აღმოჩნდა ხუთი ცილის მაკოდირებელ ალელთა და გენოტიპური კლასების სიხშირესთან.
ბაყაყის (Rana temporaria) ესთერაზული ლოკუსის მიხედვით განსხვავებული ინდივიდები ტემპერატურისადმი დიფერენცირებულ დამოკიდებულებას ავლენენ. ზაფხულის მაღალ ტემპერატურას ადვილად იტანენ ჰეტეროზიგოტი ფორმები. ასაკის ზრდასთან დაკავშირებით გენოტიპური კლასების სიხშირის ცვლილება მრავალ ძუძუმწოვარშია გამოვლენილი. მაგალითად, თეთრკუდა ირმის docoileus virginianus პოპულაციებში β - ჰემოგლობინის ლოკუსის ჰეტეროზიგოტულობის საშუალო დონე 3 წლის ასაკის ინდივიდებში მიიღწევა.
ცილების პოლიმორფიზმის ადაპტური როლი გამოვლენილია მცენარეებშიც. მრავალრიცხოვანი დამადასტურებელი არგუმენტიდან საილუსტრაციოდ მხოლოდ ერთ მაგალითს მოვიყვანთ. ველურ შვრიაში (Svriuka, Avena barbata) ხუთი გენის განსხვავებული ალელების კომბინაცია და მათი სიხშირე განაპირობებს განსხვავებულ საარსებო პირობებთან შეგუებას. ორ განსხვავებულ ეკოლოგიურ-გეოგრაფიულ ზონაში გავრცელებულია შვრიუკას სხვადასხვა პოპულაციები. თბილ მშრალ კლიმატურ ზონაში გვხვდება ე.წ. „ქსეროფიტული“, ხოლო ტენიან ზომიერ ზონაში „მეზოფილური“ პოპულაციები. შესწავლილი ხუთივე ლოკუსის მიხედვით ორივე ზონის პოპულაციები მონომორფულია, მაგრამ განსხვავებულ ალელებს შეიცავენ, რომლებიც სხვადასხვა სიხშირითაა წარმოდგენილი. გარდამავალ ცვალებად კლიმატურ პირობებში მხოლოდ ჰეტეროგენული ფორმები გვხვდება, ამასთანავე მათთვის ნიშანდობლივია მაღალი პოლიმორფულობის დონე.
პოპულაციონისტების შეხედულებით, მოლეკულურ და ორგანიზმის დონეზე ცვალებადობა ერთიანი ჯაჭვის ორი შემადგენელი რგოლია და მათი განცალკევებით განხილვა არ შეიძლება. ევოლუციის სინთეზური თეორიის პოზიციაზე მდგომ პოპულაციონისტების ამ შეხედულებას მკაცრად აკრიტიკებენ ნეიტრალური ევოლუციის თვალსაზრისზე მდგომი მეცნიერები.
|
|
11.3.5 ნუკლეოტიდური პოლიმორფიზმი |
▲ზევით დაბრუნება |
ცილების პოლიმორფიზმით მხოლოდ არაპირდაპირ შეიძლება გენების პოლიმორფულობაზე ვიმსჯელოთ. ბოლო პერიოდში საგრძნობლად დაიხვეწა და გასრულყოფილდა დნმ-ის სექვენირების მეთოდი. ეს პროცესი ავტომატიზირებულია (ამ მიზნით, მაღალი კლასის რობოტსექვინატორებს იყენებენ). გაშიფრულია 50-ზე მეტი სახეობის გენომი. ჩამოყალიბდა ახალი ბიოლოგიური დისციპლინა გენომიკა. ძირითადად სექვენირებულია ვირუსებისა და მარტივი გენეტიკური სტრუქტურის მქონე ბაქტერიების გენომი. 2003 წლის პირველ ნახევარში დასრულდა ადამიანის გენომის სექვენირება.
ადამიანის გარდა ეუკარიოტებიდან სექვენირებულია საფუარის (Saccharomyces cervisiae) გენომი. მისი დნმ 12 MB-ს შეადგენს (MB - მეგაბაზა, 1MB=106 ნ.წ.), რომელიც დაახლოებით 6340 გენს აკოდირებს, 1998 წელს დასრულდა ნემატოდას (Caenorhabditis elegans) გენომის გაშიფვრა. მისი გენომი 3-ჯერ ჭარბობს საფუარისას და 97 MB დნმ-ისგან შედგება, რაც 19100 გენს შეესაბამება. ადამიანის გენომი შეიცავს 3700 MB დნმ -ს, მაგრამ გენების რაოდენობით მხოლოდ 4-ჯერ აღემატება ნემატოდას (გააჩნია დაახლოებით 80000 გენი). გამოირკვა, რომ ევოლუციის პროცესში უმაღლეს ორგანიზმებში დნმ-ის მასა მეტი სიჩქარით იზრდებოდა ვიდრე გენების რაოდენობა. რაზედაც მიანიშნებს შემდეგი ფაქტი, რომ უმაღლეს ორგანიზმებს გაცილებით მეტი არაკოდირებადი დნმ მოეპოვება ვიდრე უმდაბლეს ეუკარიოტებს (ჰედრიკი, 2003).
დნმ-ის ანალიზი ნაირგვარი უახლესი მეთოდებით ხორციელდება. დნმ-ის მოლეკულის დაფრაგმენტება სხვადასხვა სახის ბაქტერიული რესტრიქციული ენდონუკლეაზების ანუ რესტრიქტაზებით ხორციელდება. ისინი დნმ-ის მოლეკულას სპეციფიკურ, მათთვის შესაბამის სამიზნე წერტილებში - საიტებში ხლეჩენ. საიტის ზომა საკმაოდ მცირეა და 4-6 ნ.წ.-გან შედგება. ფერმენტი დნმ-ის მოლეკულის მთელს სიგრძეზე ამოიცნობს საიტურ უბნებს და ჭრის ფრაგმენტებად. რესტრიქტაზები საიტსპეციფიკურობით ხასიათდებიან. ე.ი. კონკრეტული რესტრიქტაზა მხოლოდ მისთვის ნიშანდობლივ ნუკლეიტიდურ თანამიმდევრობაზე მოქმედებს. სხვადასხვა რესტრიქტაზების სამიზნე წერტილები - საიტები განსხვავებულია.
შესწავლილია მიტოქონდრიული დნმ-ის (მტ-დნმ) პოლიმორფულობა. მტ-დნმ მცირე ზომის წრიული სტრუქტურის მოლეკულაა. ადამიანის მტ-დნმ 16569 ნ.წ. შეიცავს, რომელშიც 16 გენია ლოკალიზებული, საფუარისაში კი 75000 ნ.წ. და შეიცავს 36 გენს (ჰედრიკი, 2003). ადამიანის მტ-დნმ-ში ფერმენტი EcoRI-სადმი (პირველად გამოყოფილია E. coli-დან) სპეციფიკური საიტების რაოდენობა 3-ია, ფუტკრისაში - 5, ხოლო საფუარისაში - 4. როგორც ცალკეული ინდივიდი ისე მთლიანად პოპულაცია განსხვავდება სხვადასხვა რესტრუქციული საიტების რაოდენობით. ეს უკანასკნელი საფუძვლად უდევს რესტრიქციული ფრაგმენტების სიგრძივ პოლიმორფიზმს. ვინაიდან დნმ-ის მოლეკულაში ახალი საიტების დაგროვება სწრაფად მიმდინარეობს, რესტრიქციული საიტების ანალიზით ახლო ნათესაურ ჯგუფებს შორის ურთიერთკავშირის შეფასებას ახდენენ. ამასთანავე ოთხ - ან ექვსწევრიანი თანამიმდევრობების მქონე საიტებზე ბუნებრივი გადარჩევა არ მოქმედებს. ამიტომაც ყოველი ზემოთ დასახელებული ზომის საიტი არ ევოლუირებს, უნიკალურია და ერთხელ ყალიბდება.
აშშ-ს სამხრეთ-აღმოსავლეთში (ალაბამა, ჟორჟია, ფლორიდა) გავრცელებულ ჩანთოსან ვირთაგვას (Geomys pinetis) პოპულაციებში შეისწავლეს მტ-დნმ-ში რესტრიქციის 6 განსხვავებული საიტი. გამოვლენილია 23 განსხვავებული ჰაპლოტიპი. გეოგრაფიულად მოსაზღვრე პოპულაციები ერთიმეორის მსგავსი აღმოჩნდა. ჰაპლოტიპები ერთმანეთისგან უმნიშვნელოდ (ერთი ან ორი საიტით) განსხვავდებოდნენ, მაშინ როდესაც დასავლეთსა და აღმოსავლეთში გავრცელებული პოპულაციები რესტრიქციულ საიტთა განაწილების მიხედვით საკმაოდ განსხვავდებოდნენ. თითოეული მათგანი 9 სხვადასხვა საიტს ფლობდა.
ბოლო პერიოდში დნმ-ის ტექნოლოგიაში მიღწეული შედეგების გამოყენებით შესაძლებელია ერთნუკლეოტიდიანი პოლიმორფიზმის შესწავლა. თავდაპირველად ეს ფენომენი დროზოფილას (D. melanogaster) ალკოჰოლდეჰიდროგენაზას მაკოდირებელ გენში (ADH) გამოიკვლიეს. გაანალიზებულია ADH თერთმეტი ლოკუსი. გამოვლენილ 49 ვარიაბილური ლოკუსიდან 43-ში ნუკლეოტიდთა თანამიმდევრობა აღმოჩნდა შეცვლილი, 6 საიტს კი ინსერცია და დელეცია მოეპოვებოდა.
სხვადასხვა სახეობის პოპულაციებში შესწავლილი ცილის მაკოდირებელი გენების დიდი უმრავლესობა დაბალ პოლიმორფულობას ავლენდა. მხოლოდ ზოგიერთ ლოკუსს აღენიშნებოდა პოლიმორფულობის მაღალი დონე. მათ რიცხვს მიეკუთვნება ხერხემლიან ცხოველებში გამოვლენილი ჰისტოთავსებადობის კომპლექსური MHC გენები. ადამიანში შესწავლილია MHC გენების (მათ HLA გენები ეწოდება) ვარიაბილობა. ეს გენები რეაგირებენ გადანერგილ უცხო ორგანოზე და იმუნურ შეუთავსებლობას იწვევენ. მეტად მნიშვნელოვანია HLA გენების როლი იმუნური დაავადებისა და პათოგენების ამოცნობაში. ამ გენების სექვენირებით გამოირკვა, რომ HLA-A, HLA-B და HLA-DC ლოკუსებს შესაბამისად 67, 149 და 179 ალელი მოეპოვებათ. HLA-A ლოკუსის ჰეტეროზიგოტულობა 0,064, HLA-B ლოკუსის კი 0,058-ს შეადგენს. HLA გენში მომხდარი ცვლილებები, კოდირებულ ცილის მოლეკულაში ისეთ საიტებს ცვლის რომლებიც თ - ლიმფოციტებში შესაბამისი იმუნური პასუხის გამომუშავებას განაპირობებს.
|
|
12 §10. ადაპტაცია - ბუნებრივი გადარჩევის შედეგი |
▲ზევით დაბრუნება |
ადაპტაცია არის ორგანიზმთა მორფოლოგიურ-ქცევითი, პოპულაციურ და სხვა ნებისმიერ თავისებურებათა ერთობლიობა, რომელიც ხელს უწყობს კონკრეტულ საარსებო გარემოში სახეობის არსებობას. ადაპტაცია ბუნებრივი გადარჩევის მოქმედების შედეგად ყალიბდება. ტერმინი „ადაპტაცია“ ორი მნიშვნელობით - შეგუების წარმოქმნის პროცესის (ადაპტაციოგენეზი) და თვით მისი შედეგის (ადაპტაციური ნიშნის) აღსანიშნავად იხმარება. ადაპტაცია ინდივიდის სასიცოცხლო ციკლის ნებისმიერ ეტაპზე ყალიბდება. ჩ.დარვინის მიერ ორგანული სამყაროს ევოლუციის მექანიზმების აღმოჩენამ დაადასტურა, რომ მთელი ევოლუცია არსებითად ადაპტაციაა. ამიტომაც ევოლუცია შეიძლება განხილულ იქნეს, როგორც ადაპტაციის წარმოქმნის, განვითარებისა და გარდაქმნის პროცესი. ვიწრო მნიშვნელობით ადაპტაცია არის კონკრეტული მორფოფიზიოლოგიური თვისებების ჩამოყალიბება, რითაც ორგანიზმი შესაბამის საარსებო გარემოს ეგუება და სიცოცხლის შენარჩუნებასა და შთამომავლობის დატოვებას ემსახურება.
|
|
12.1 10.1. ადაპტაციის მაგალითები |
▲ზევით დაბრუნება |
ცოცხალ ორგანიზმთა შეგუების დამამტკიცებელი არგუმენტები იმდენად მრავალფეროვანი და მრავალრიცხოვანია, რომ ყველა ფორმის სრულყოფილად აღწერა და გადმოცემა შეუძლებელია. ამდენად რამდენიმე კლასიკური ნიმუშით შემოვიფარგლებით. ადაპტაცია ნიშანდობლივია ცოცხალი სისტემის ორგანიზაციის ყველა დონისათვის. იგი ყველაზე მკაფიოდ პასიური თავდაცვის საშუალებათა გამომუშავებისას ვლინდება. პასიური თავდაცვის სამარჯვეებს წარმოადგენს ისეთი სტრუქტურები და თავისებურებები, რომელთა არსებობითაც მიიღწევა ინდივიდების მაღალი სიცოცხლისუნარიანობა, ფერტილურობა და კონკურენტუნარიანობა.
|
|
12.1.1 მფარველობითი (კრიპტული) შეფერილობა და ფორმა |
▲ზევით დაბრუნება |
ის ცხოველებში ერთ-ერთი მნიშვნელოვანი პასიური თავდაცვის საშუალებაა. იგი ფართოდ არის გავრცელებული ცხოველთა სამეფოში. მფარველობით შეფერილობაში იგულისხმება გარემო ფონთან ორგანიზმის შეფერილობის მსგავსება, რის გამოც ცხოველი ძნელად შესამჩნევი ხდება. იგი გამოიყენება მტაცებელთაგან თავდასაცავად. მაგრამ ამდაგვარი დაცვითი ფორმა ხშირად გვხვდება მტაცებლებშიც (მაგ., თეთრი დათვი, ლომი, ტყის კატა და მისთ.), რაც მათ ხელს უწყობს შეუმჩნევლად ჩაუსაფრდეს ან მიეპაროს მსხვერპლს.
მფარველობითი შეფერილობა ონტოგენეზის ნებისმიერ სტადიას ახასიათებს. ზღვის პლანქტონის მრავალი ცხოველის კვერცხები, ლარვები და ზრდასრული ფორმები გამჭვირვალე და მტრისთვის შეუმჩნეველია. კვერცხები, რომელსაც მწერები ფოთლებზე და ყლორტებზე დებენ მწვანეა, იქიდან გამოსული ლარვებიც მეტწილად მწვანე ფერისაა. ხეებისა და ბუჩქების ქერქზე პეპლების მიერ დადებული კვერცხები მეტწილად ღია რუხი ან მურა-რუხი ფერისაა. გამოჩეკილი მუხლუხებიც, ასევე ჩამოყალიბებული ჭუპრებიც რუხი ან მურა ფერისაა. ფრინველები, რომლებიც ბუდეს ფარულ ადგილას (სორო, ხის ფუღურო და მისთ.) იშენებენ, კვერცხებს თეთრს დებენ, ხოლო რომლებიც ღია ადგილებში იშენებენ ბუდეს, კვერცხის ნაჭუჭი ნაირგვარი შეფერილობისაა და ეხამება გარემო ფონს. როგორც ბუდე ისე კვერცხები მტაცებლებისთვის ნაკლებ შესამჩნევია (სურ. 11.1).

სურ. 10.1. A - პრანჭიას (Vanellus vanellus) ბუდე და კვერცხები ეხამება გარემო ფონს, იგი პოტენციური მტრისათვის ნაკლებ შესამჩნევია. B - სუსხური (Somateria mollissima) ბუდეზე (ა. იაბლოკოვი, ა. იუსუპოვი, 2004).
![]()
სურ. 10.2. ხავერდულა (Satyridea) მუხლუხი. მწვანე ბალახზე ბინადარი მწვანე მუხლუხს სიგრძივი ზოლები გასდევს. მწვანე ფონზე მუხლუხი ძნელად შესამჩნევია (ი. შმალგაუზენი)
ზრდასრული ცხოველების უმეტესობა ეხამება გარემო ფონს. მწვანე გარემოში მობინადრე ცხოველი უმეტესად მწვანე ფერისაა. მაგალითისათვის შეიძლება დავასახელოთ მწვანე კუტკალია (Tettigonia viridisima), მწვანე ბაღლინჯოები, ბუზები, მწვანე ჩოქელა (Mantis religiosa), ნემსიყლაპიები, მწვანე ბაყაყი (Rana esculenta), ვასაკა (Hyla arborea) მწვანე ხვლიკი (Lacerta virdis), ხის გველები და სხვ. ნექტონურ თევზებს ხშირად ვერცხლისფერი მუცელი და მურა მწვანე ზურგი აქვს, რის გამოც ზემოდან ზღვის წყლის ფონზე და ქვემოდან წყლის მოვერცხლისფრო (სხივების ნაკადის სრული არეკვლის გამო) ზედაპირზე ძნელად შესამჩნევია. თევზები (მაგ. ნემსთევზა და მისთ.), რომლებიც ზღვის წყალმცენარეებს აფარებენ თავს შესაბამისად მწვანე, წითელი ან მურა შეფერილობისაა. ის თევზები, რომლებიც კაშკაშა მარჯნის რიფებში ბინადრობენ, შეფერილობითაც კაშკაშაა.
ღამის ცხოველებს აქვთ ოდნავ შესამჩნევი რუხი შეფერილობა (ბუ, ღამურა, ვირთაგვა, თაგვი...). უდაბნოსა და ნახევარ უდაბნოს ცხოველები მოყვითალო ან მურა-მოყვითალოა (მაგალითად, უდაბნოს კალია, ვარანი, კობრა, ხვლიკი, ანტილოპა, აქლემი, ლომი და სხვ.). ზოგიერთ ცხოველს (კამბალა, აგამისებრი ხვლიკი, ქამელეონი...) კანში ან სხეულის სხვა საფარველში ლოკალიზებულ ქრომატოფორებში პიგმენტების სწრაფი გადანაწილებით შეფერილობა მალე ეცვლება გარემო ფონის მიხედვით. ჩრდილოეთში ბინადარ ზოგიერთ ცხოველს (სიასამური, ყარსაღი, თეთრი კურდღელი, თეთრი გნოლი და სხვ.) ახასიათებს შეფერილობის სეზონური ცვლა ე.წ. სეზონური დიმორფიზმი. ზაფხულის მუქი შეფერვა ზამთრისთვის თეთრით ეცვლება, რის გამოც ისინი თოვლის საფარზე მტრისათვის შეუმჩნეველია.
ცხოველთა საერთო შეფერილობას თან ერთვის მოხატულობა (წინწკლები, ზოლები, ხაზები) რის გამოც იგი ფონს მეტად ემსგავსება. მაგ., ღამის პეპლები ხის ქერქზე გარინდულად ყოფნისას მოხატულობით მღიერებს ან დაბზარულ ქერქს ემსგავსებიან და ფრინველებისათვის შეუმჩნეველია. პეპლის მწვანე მუხლუხებს ზოგჯერ სიგრძივი ზოლები გააჩნიათ, რის გამოც ბალახზე მათი გარჩევა ძნელია (იხ. სურ. 10.2).
|
|
12.1.2 დანაწევრებითი (დიზრუპტული) შეფერილობა |
▲ზევით დაბრუნება |
იგი მფარველობითი შეფერილობის კერძო შემთხვევას წარმოადგენს. ცხოველს აქვს კონტრასტული ზოლები ან ხალები, კერძოდ სხეულზე მუქი და ნათელი უბნები ერთიმეორეს ენაცვლება. მკვეთრი ხაზებით თითქოს შემოსაზღვრულია სრულიად განსხვავებული სხეული, რომელიც არაცოცხალ საგნებს ან მცენარეებს ჩამოგავს. სხეულის ერთიანი ფორმა გარემო ფონზე იკარგება (სურ. 10.3). ამდაგვარი შეფერილობა ბუნებაში ფართოდაა გავრცელებული. იგი ახასიათებს ზოგიერთ მწერს, ამფიბიას, ქვეწარმავალს, ფრინველსა და ძუძუმწოვარს (სურ. 10.4). მაგალითად ზებრა (Equus greveyi, E. bohme, E. zebra) და ვეფხვი (Panthera tigris) გაშლილ ადგილებშიც კი 50-70 მ-ის მანძილიდან ძნელი შესამჩნევია სხეულზე არსებული ზოლებისა და გარემოს შუქ-ჩრდილების თანხვედრის გამო. რიგ შემთხვევაში მარტო ცხოველის კონტურები კი არ ერწყმის გარემოს, არამედ გაძნელებულია ცხოველის სასიცოცხლო მნიშვნელობის ორგანოების გარჩევა. მაგალითად, მრავალ არამონათესავე ცხოველს თვალი მუქი ზოლებით ან ლაქებით მსგავსად აქვთ შენიღბული (სურ. 10-5).

სურ. 10.3. დიზრუპტული შეფერილობის მნიშვნელობა: 1. მთლიანი შეფერილობა. 2. დანაწევრებითი შეფერილობა. 3. დანაწევრებითი შეფერილობა შესაბამის ფონზე (ჰ. კოტი, 1950).

სურ. 10.4. დანაწევრებითი შეფერილობა ბაყაყში (Megalixalus fornasinii). მსვენებარე პოზაში ზოლები ერთმანეთს თანხვდება, ერთიანი ფორმა გარემო ფონზე იკარგება (ჰ. კოტი, 1950).
დიზრუპტული შეფერილობა თავდასაცავად ეფექტურია, საარსებო გარემოში ადეკვატური ფონის არსებობისას. კრიპტული შეფერილობის ეფექტი ძლიერდება მაშინ თუ იგი შეუღლებულია კრიპტულ ქცევასთან. საფრთხის შემთხვევაში ცხოველები (ობობები, მწერები, ამფიბიები, გველები, ფრიველები და სხვ.) ირინდებიან ე.წ. „მოსვენებით პოზას“ იღებენ. მაგალითად ლერწმიანში ბინადარი ყარაულა (წყლის ბუღა, Bოტაურუს სტელლარის) საშიშროების შემთხვევაში იჭიმება, კისერსა და თავს ვერტიკალურად მაღლა სწევს და ინაბება. შეფერილობით (მოწითურო მურაა, მუქი განივი და გასწვრივი ხალებით) გარემოსთან შეხამებული ფრინველის შემჩნევა ახლო მანძილიდანაც კი ძნელია. კრიპტული ქცევები ახასიათებთ იმ ცხოველებს, რომლებიც მტრისგან აქტიურად თავს ვერ იცავენ.
სურ. 10.5. თვალის მუქი ზოლით შენიღბვა: A - თევზი, B - გველი, C - ბაყაყი, D - ჩანთოსანი ჭიანჭველაჭამია (ჰ. კოტი, 1950).
|
|
12.1.3 გამაფრთხილებელი (აპოსემიური) და დემონსტრაციული შეფერილობა |
▲ზევით დაბრუნება |
ამ ტიპის შეფერილობა კრიპტული შეფერილობის საპირისპიროა. ცხოველს აქვს თვალშისაცემი კაშკაშა შეფერილობა. იგი მოეპოვება ზოგ ცხოველს და საკვების უვარგისობაზე წინასწარ მიანიშნებს. გამაფრთხილებელი შეფერილობა მფლობელს იცავს მხოლოდ იმ ცხოველებისაგან, რომელთაც კარგად განვითარებული მხედველობის ორგანოები გააჩნიათ და ფერთა გარჩევის უნარს ფლობენ. თავდამსხმელს კარგად განვითარებული ნერვული სისტემისა და რთული რეფლექსური მოქმედების მეოხებით ამა თუ იმ ფერზე პირობითი რეფლექსების გამომუშავების და შეკავების უნარი გააჩნიათ. გამაფრთხილებელი შეფერილობის ცხოველებს აქვთ მსუსხავი უჯრედები (მრავალი აქტინია), შხამიანი ჯირკვლები და ეკლები (მრავალფეხა, ზოგიერთი მწერი, თევზი, გველი და ხვლიკი), შხამიანი ლორწო ან სხეული (სალამანდრა, გომბეშო, ვასაკა და ზოგიერთი თევზი) არასასიამოვნო სუნის ან მწველი სითხის გამოყოფის უნარი (სხვადასხვა მყრალა, ზოგიერთი ხოჭო, ბაღლინჯო, პეპელა). თავდამსხმელი ცხოველი რამდენიმე სინჯის შემდეგ სწავლობს და სწრაფად იმახსოვრებს საკვების ვარგისიანობას. გამაფრთხილებელი შეფერილობის ცხოველები მტაცებელს გარკვეული თავდაცვითი ქცევებითაც (ე.წ. „მუქარის პოზები“) პასუხობენ. მრავალი მათგანი საშიშია თავდამსხმელთათვის. მაგალითისათვის შეიძლება დავასახელოთ კაშკაშად შეფერილი შხამიანი გველები. კერძოდ, აფრიკული კაშკაშა ასპიტების (Elaps) სხეული გარშემოვლებულია შავი და თეთრი შეფერილობის რგოლებით. ამერიკაში გავრცელებული მარჯნისფერი ასპიტების (Micruruს) წითელ სხეულზე განლაგებულია ვიწრო ყვითელი ან მწვანე ზოლებით შემოფარგლული შავი რგოლები.
კრიპტულ და აპოსემიურ შეფერილობას თუ ერთმანეთს შევადარებთ აშკარაა, რომ პირველი ახასიათებთ დაუცველ ცხოველებს. სასიცოცხლო მნიშვნელობის სტრატეგიაა, რომ ისინი პოტენციური მტრებისათვის შეუმჩნეველი და მოუხელთებელი იყვნენ. მეორე ფორმა, კი მოეპოვებათ კარგად დაცულ ცხოველებს. მისი არსი გამოიხატება თეზით: „ხელს ნუ მახლებ“. მართლაც იგი გამაფრთხილებელი შეფერვაა, რომელიც პოტენციურ მტერში სამარჯვეების მეშვეობით უსიამოვნო შეგრძნებებს, ტკივილს ან მოწამვლას იწვევს.
დემონსტრაციული შეფერილობა არის მფარველობითი შეფერილობის ერთ-ერთი ტიპი, რომელიც ხელს უწყობს ინდივიდის გადარჩენას. ცხოველს გააჩნია სპეციფიკური შეფერილობა, რომლითაც გარემოს ფონზე შესამჩნევი ხდება. ამ შეფერილობის დემონსტრირებას ცხოველი მხოლოდ საფრთხის შემთხვევაში ახდენს, რომელსაც ჩვეულებრივ თან ახლავს სპეციფიკური ხმის გამოცემა, რიტუალური თავდაცვითი ქცევები ან არასასიამოვნო სუნის ნივთიერების გამოყოფა. მაგალითად, ღამის პეპელა ფრთახალა სფინქსს (Smerinthus ocellatus) უკანა ფრთებზე აქვს ფრინველის თვალის მსგავსი ლურჯი მოხატულობა, რომელიც სიბნელეში ბზინავს. საფრთხისას პეპელა ფრთებს შლის და მბზინავ ორ ლაქას თავდამსხმელი ღამის მტაცებელი ფრინველის თვალად, ხოლო მუცელს ნისკარტად აღიქვამს და ფრთხება.
სამხრეთ ამერიკაში გავრცელებული პეპელა სფინქსის (Leucorampha) მუხლუხს, რომელიც ხის ყლორტებითა და ფოთლებით იკვებება, წინა ნაწილში გააჩნია თვალის მსგავსი მოხატულობა. საფრთხისას მუხლუხი სხეულის წინა ნაწილს მაღლა სწევს, თავს ბერავს, თვალის მსგავსი მოხატულობის ადგილი ებურცება. იგი იერსახით გველს (Bothrops schlagelli) ემსგავსება. შეშინებული თავდამსხმელი სწრაფად შორდება მუხლუხს.
დემონსტრაციული შეფერილობა ყოველთვის შეუღლებულია სპეციფიკურ „მუქარის პოზასთან“. სანიმუშო მაგალითია უდაბნოებში, ნახევრად უდაბნოებსა და სტეპებში ბინადარი ტარანტული. როდესაც ეს ლამაზი, დიდი ზომის ობობა საფრთხეშია, სწრაფად აღიმართება და ორ წყვილ ფეხს მაღლა წევს. კიდურების ზედა ზედაპირის მონოტონური რუხი ფერის ნაცვლად თავდამსხმელს უეცრად თვალში ხვდება ფეხების ქვედა ნაწილის მკვეთრი, კაშკაშა თეთრი და შავი ხავერდოვანზოლიანი მოხატულობა. ხავერდოვან შავ მუცელზე კარგად მოჩანს ნარინჯისფერი ლაქა ამდაგვარ პოზაში ობობა აფრთხობს მტერს.
|
|
12.1.4 მიმიკრია |
▲ზევით დაბრუნება |
მიმიკრია, მფარველობითი შეფერილობის ერთ-ერთი ნაირსახეობაა. მიმიკრია არის ერთი სახეობის თავდაცვის საშუალებებს მოკლებული ცხოველის მიმსგავსება კარგად დაცულ და საჭმელად გამოუსადეგარ არამონათესავე სახეობებთან (მიმეტიზმი) ან გარემოს რომელიმე საგანთან და მცენარეებთან (მიმეზია).
მიმეზიის დროს ცხოველები ემსგავსებიან მცენარის რომელიმე ნაწილს ან გარემოს საჭმელად გამოუსადეგარ საგნებს. გარემოს საგნებთან ცხოველთა მიბაძვის მრავალი მაგალითია ცნობილი. პეპელა მზომელას მატლი საშიშროების შემთხვევაში ხის ტოტზე იჭიმება და როკს ემსგავსება. ჩოქელა სხეულის ფერითა და ფორმით ზოგიერთ ბალახოვან მცენარეთა ფოთოლს ემსგავსება (სურ. 10.6), ჩხირულები ხის ტოტს ემსგავსებიან. ინდონეზიური პეპლის კალიმას ფრთების ქვედა ზედაპირი გამხმარ ფოთოლს ჩამოგავს.
მიმეტიზმი არის მიმიკრიის კერძო შემთხვევა. თავდაცვის საშუალებებს მოკლებული ცხოველები შხამიან ან საჭმელად უვარგის გენეტიკურად არამონათესავე (აპოსემიური ეფერილობის) სახეობებს ემსგავსებიან. მიმიკრიისას ერთი ორგანიზმი იმიტატორი (საჭმელად ვარგისი ფორმა) გარეგნობით ემსგავსება მეორეს - მოდელს (აპოსემიური შეფერილობის ფორმას), რომელსაც შეცდომაში შეჰყავს მესამე - ოპერატორი (პოტენციური მტერი).

სურ. 10.6. მიმეზია მწერებში. A - ჩოქელა ფორმით ემსგავსება მარცვლოვანთა ფოთოლს; B - ფილუმი ემსგავსება ფოთოლს; C - ჩხირულა ემსგავსება ხის ტოტს (მ. ბელიაევი, 1947).
მიბაძვითი მსგავსება დასაცავად მხოლოდ გარკვეულ პირობებში გამოდგება. მოდელს უნდა ჰქონდეს გამაფრთხილებელი აპოსემიური შეფერილობა და რაოდენობრივად უნდა აჭარბებდეს იმიტატორს; როგორც მოდელი ისე იმიტატორი უნდა ბინადრობდნენ ერთსა და იმავე ტერიტორიაზე და უნდა ჰქონდეთ მსგავსი ინსტინქტები. წინააღმდეგ შემთხვევაში მიმიკრია იმიტატორისათვის საზიანოა. მტაცებლები ადვილად ამჩნევენ „სიყალბეს“ და იწყებენ მათ განადგურებას.

სურ. 10.7. მიმიკრია პეპლებში; 1 - საჭმელად უვარგისი პეპელა helikonida; 2 - იმიტატორი პეპელა pierida (ნ. იორდანსკი, 2001).
ცხოველებში გამოყოფენ ორ ძირითად ფორმას: ბეითსისეულ და მიულერისეულ მიმიკრიას. ბეითსის მიმიკრიის დროს (პირველად აღწერა გამოჩენილმა ბუნებისმეტყველმა და მოგზაურმა გ. ბეითსმა). საარსებო გარემოში იმიტატორი (საჭმელად ვარგისი, თავდაცვის საშუალებებს მოკლებული ფორმა) და მოდელი (შხამიანი ან საკვებად გამოუსადეგარი ფორმა) ერთდროულად არიან. მაგალითად, პეპლების ოჯახის ჰეპლიკონიდების წარმომადგენლებს, რომელთაც აპოსემიური შეფერილობა და არასასიამოვნო სუნი აქვთ, ემსგავსებიან ფრინველებისათვის საჭმელად ვარგისი ამაზონის ველის პეპლები თეთრეულასებრთა ოჯახიდან (იხ. სურ. 10.7). კუნძულ შრილანკაზე გავრცელებული ობობა Myrmarachne formosa ბაძავს იმ ჭიანჭველებს, რომლებიც ბუდეს ხის ფოთლებზე იკეთებენ (იხ. სურ. 10.8). ბეითსის მიმიკრია აღწერილია ხერხემლიან ცხოველებშიც. მაგალითად, გუგულის მიმსგავსება მიმინოსთან. უწყინარი ანკარა სიმოფისის (Simophis rhinostoma) იმიტირება მარჯნისფერ ასპიტთან (Micrurus frontalis, აღენიშნება მორიგეობით განლაგებული დიდი ზომის წითელი და პატარა ზომის შავი და ყვითელი ირგვლივი ზოლები).

სურ. 10.8. ობობა (Myrmarachne formosa) იმიტირებს ჭიანჭველას (მ. ბელიაევი, 1947).
მაგალითად, უწყინარი უშხამო აფრიკული კვერცხისმჭამელი გველი (Dasypeltis scaber), როგორც შეფერვით (სხეული მურა-რუხია, თავზე აქვს თეთრი ჯვრისმაგვარი ნახატი, ზურგის გაყოლებით თეთრი მუქარშიიანი ლაქები, გვერდზე თეთრი ზიგზაგისებური ხაზები მუქი არშიით) და ფორმით, ისე მუქარის დემონსტრაციით იმიტირებს მეტად შხამიან გველს ქვიშნარის ეფას (Echis carinata). როდესაც გველი შეშინებულია სხეულის ერთმანეთთან მჭიდროდ მიჯრილ ნახევარრგოლებად კეცავს, გვერდზე განლაგებული ქერცლების ხახუნით გამოსცემს სპეციპიკურ ბგერებს (იხ. სურ. 10.9). პეპელა მინაფრთიანა, არა მარტო გარეგნულად წააგავს კრაზანას, არამედ მუქარის ნიშნად ბზუის კიდეც. ბუზი ჩუხჩუხელა, რომელიც მოხატულობით კრაზანას ასლია, წინა ფეხებს კრაზანას ულვაშების მსგავსად ამოძრავებს და მასავით ბზუის (იხ. სურ. 10.10). მოხატულობით ხმელი ფოთლის მსგავსი მრავალი პეპელა საშიშროებისას მცვივანი ფოთლების მსგავსად წრიულად მოძრაობს და მიწაზე დაყრილ ფოთლებზე ეშვება. მაშასადამე, როდესაც შეფერილობა და ჩამოყალიბებული თავდაცვითი ქცევები (მორფოლოგიური და ფიზიოლოგიური ნიშნები) ურთიერთშეხამებულია იმიტატორი სრულყოფილად არის მიმსგავსებული მოდელთან.
სურ. 10.9. A - აფრიკაში გავრცელებული შხამიანი გველი ქვიშნარის ეფა (Echis carinata) მუქარის დემოსტრაციისას, B - მას იმიტირებს აფრიკული უშხამო გველი (Dasypeltis scaber). (დ. მაკ-ფარლენდი, 1988).

სურ. 10.10. ბეითსის მიმიკრია ბუზებში (ბაძავენ კრაზანას). A - აპოსემიური შეფერილობის კრაზანა ოდინარი (Odynerus parietum); B - ბუზი სტრატიომია (Stratiomia chamaeleon) კრაზანას ბაძავს მუცლის შეფერილობით და ულვაშების ფორმით; C - ბუზი-სილფიდა (Chrysotoxum bicinetum) კრაზანას ბაძავს შეფერილობით და ნაწილობრივ სხეულის ფორმით; D - ბუზი-თავდიდა (Conops flavipes) გარეგნობით წარმოადგენს ზოგიერთი კრაზანას ასლს (ა. იაბლოკოვი, ა. იუსუპოვი, 2004).
ცხოველი, მიმიკრიას თავდასხმისათვის იშვიათად იყენებს. ე.წ. აგრესიული მიმიკრიის სანიმუშო მაგალითია თევზი - დამსუფთავებელი - ტუჩა (Labroides dimidiatus) და მისი იმიტატორი ასპიდონტი. ისინი წყნარი და ინდოეთის ოკეანეების მიმდებარე ნაწილში ცხოვრობენ. ტუჩა თევზს სხეულზე სიგრძივად შავი და თეთრი ზოლები გასდევს. სპეციფიკური შეფერილობითა და ცურვის ხასიათით მას ადვილად ცნობენ მსხვილი თევზები. მათ შორის მტაცებლებიც კი და დასუფთავების შესაძლებლობას აძლევენ. ტუჩა მათ ტანსა და პირის ღრუს პარაზიტებისაგან უსუფთავებს (სიმბიოზი), რომლითაც იგი იკვებება. ტუჩას იმიტირებს ხმალკბილა ზღვის ფინია ასპიდონტი (Aspidontus taeniantus). ასპიდონტი ტუჩას ემსგავსება, როგორც ზომითა და სხეულის შეფერილობით, ისე ცურვის მანერით. მსგავსება იმდენად სრულყოფილია, რომ დიდი თევზები ასპიდონტს ტუჩა დამსუფთავებლად მიიჩნევენ. ისინი ნებას რთავენ მიუახლოვდეს და დაასუფთავოს. ასპიდონტი თავს ესხმის მასპინძელს, ფარფლიდან ნაკუწს გლეჯს და სწრაფად უჩინარდება (აგრესიული მიმიკრია პეკემისეული მიმიკრიის სახელით იწოდება ცნობილი მკვლევარის პეკემის პატივისცემის ნიშნად).
ზოგჯერ იმუნური ფორმები გარეგნობით ერთმანეთს ბაძავენ. ეს მოვლენა პირველად გამოჩენილმა ენტომოლოგმა ფ.მიულერმა აღწერა. მიმიკრიის ამ ფორმას მიულერისეულ მიმიკრიას უწოდებენ. სამხრეთ ამერიკაში გავრცელებული პეპლების სხვადასხვა ოჯახის წარმომადგენლები (Danaidae, Neotropidae, Heliconidae, Acraeidae). ერთმანეთს ემსგავსებიან და ისინი ერთად ბინადრობენ. ამ იმუნური პეპლების ურთიერთმიბაძვით „მიმიკრიის რგოლები“ წარმოიქმნება, რომელიც ხელს უწყობს მასში გაერთიანებული სახეობების გადარჩენას.
მტაცებელს გამოცდილება მსხვერპლის საჭმელად ვარგისიანობაზე პირადი გამოცდილების საფუძველზე თანდათანობით უგროვდება. გაღიზიანებათა გარჩევის საფუძველზე მიმდინარეობს დასწავლა.
ზემოთ მოტანილი მაგალითის შემთხვევაში ფრინველები იმუნურ ფორმებს საკუთარი გამოცდილების საფუძველზე ცნობენ. პეპლების გარკვეული ნაწილი ახალგაზრდა ან გამოუცდელი (დაუსწავლელი) ფრინველების მსხვერპლი ხდება. პეპლების ეს რაოდენობა სხვადასხვა სახეობების პოპულაციათა შორის ნაწილდება, რის გამოც ამა თუ იმ სახეობის ინდივიდთა ელიმინაცია მცირდება.
მცენარეებში მიმიკრია ძირითადად სასარგებლო ცხოველთა მიზიდვისა და მავნე ცხოველთა დაშინებას ემსახურება. მაგალითად, ჭინჭარს (მოდელი) ძლიერ ემსგავსება სრულიად სხვა ოჯახში გაერთიანებული ჭინჭრის დედა (იმიტატორი). ზოგიერთი მცენარე (იმიტატორი) მხოლოდ გარკვეული ორგანოთი ემსგავსება სხვა მცენარეებს (მოდელს) და ამით ატყუებს ცხოველებს. მაგალითად, უნექტარო ყვავილები ემსგავსებიან ნექტრიანებს და მწერებს დასამტვერად იზიდავენ. ჯადვარის ყვავილი ემსგავსება მდედრ ფუტკარსა და კრაზანას და მამრ ინდივიდს იზიდავენ. ზოგიერთი მწერიჭამია მცენარის ფოთოლი ყვავილს ბაძავს შეფერილობით, ფორმით და გამოყოფს სურნელოვან მწებვარე ნივთიერებას (მაგ., დროზერა, ვენერას ბუზიჭერია).
მიმიკრია ყალიბდება ბუნებრივი გადარჩევის ხანგრძლივი მოქმედების შედეგად. იმ ორგანიზმებს, რომელთაც აღმოაჩნდებათ გარეგნულ ნიშნებში ან ქცევებში უმნიშვნელო გადახრა, რაც აძლიერებს მათ მსგავსებას კარგად დაცულ ფორმებთან, მათ გადარჩენისა და მეტი შთამომავლობის დატოვების უნარი გააჩნიათ დანარჩენ ფორმებთან შედარებით, რომლებსაც ამდაგვარი სასარგებლო ცვლილებები არა აქვთ.
|
|
12.1.5 პასიური თავდაცვის სხვა საშუალებები. |
▲ზევით დაბრუნება |
მათ მიეკუთვნება ისეთი სტრუქტურები და თავისებურებები (სამარჯვეები), რაც არსებობისათვის ბრძოლაში სიცოცხლის შენარჩუნებას ემსახურება. ზოგიერთი ცხოველი გარედან დაფარულია მკვრივი, სქელი საფარველით. მაგალითად, კიბოსა და კიბორჩხალას აქვს ჯავშანი, ხოჭოებს ქიტინოვანი ზედა ფრთები და საფარველი. ჯავშნითაა დაფარული ეკალკანიანების სხეული. კუს სხეული შეჯავშნულია ძვლოვანი ფურცლებისაგან შემდგარი ბაკნით. ნიანგის სხეული დაფარულია მსხვილი რქოვანი ფარებით. ძუძუმწოვრების თანამედროვე ფორმებს შორის კარგად განვითარებული ჯავშანი აქვთ ჯავშნისებრსა და პანგოლინებს.
მცენარეთა (მაგ., მარცვლოვანების) უჯრედის გარსის მინერალიზაციის (კაჟოვანათი ან კალციუმის მარილებით) გამო, ზოგიერთი მწერი მას არ ჭამს. ზოგიერთ მცენარეს მოეპოვება კარგად განვითარებული ეკლები (მაგ., კოწახური, კაქტუსი და სხვ.) და ქაცვები (მაგ., ასკილი, მაყვალი და სხვ.), რომლებიც იცავენ მათ ცხოველებისაგან. ზოგიერთი ცხოველი ეკლებით (მაგ., მაჩვზღარბა, ზღარბი და სხვ.) თავს იცავს მტაცებელი ცხოველებისაგან. ამავე ფუნქციას ემსახურება ზოგიერთ მცენარეში ბუსუსები (მაგ., ჭინჭარი, სალბი და სხვა).
|
|
12.1.6 რთული ადაპტაციები |
▲ზევით დაბრუნება |
ევოლუციის სინთეზური თეორია, კარგად ხსნის იმ ურთულესი შეგუების წარმოშობას, რომლებიც მცენარეებსა და ცხოველებს მოეპოვებათ. ვარაუდობენ, რომ რთული ადაპტაციები ყალიბდებოდა დროის დიდ შუალედში. ბუნებრივი გადარჩევის მოქმედების შედეგად უმნიშვნელო სასარგებლო გადახრების თანდათანობითი დაგროვების საფუძველზე. განვიხილოთ ზოგიერთი მათგანი.
|
|
12.1.7 მწერიჭამია მცენარეები. |
▲ზევით დაბრუნება |
მტაცებლობა ავტოტროფ ორგანიზმებში კერძოდ მცენარეებში იშვიათი მოვლენაა. ყვავილოვან მცენარეთა შორის გვხვდება 500-მდე სახეობის მწერიჭამია მცენარე, რომლებიც მეტამორფოზული ფოთლების მეშვეობით იჭერენ მწერებს (იშვიათად სხვა პატარა ცხოველებსაც) და საკვებად იყენებენ. ამ ტიპის მცენარეებიდან საქართველოში გვხვდება დროზერა ანუ ცვრიანა, ალდროვანდა, ბუშტოსანა და ცხიმურა. ისინი მრავალწლოვანი ბალახოვანი მცენარეებია.
სურ. 10.11. დროზერა (Drosera rotundifolia) A - მცენარე; B - ფოთოლი.
ტორფიან ჭაობებში გვხვდება მცირე ზომის ყვავილოვანი მცენარე დროზერა (Drosera rotundifolia). მრგვალი ფორმის ფოთლის ფირფიტის ზედაპირი კაშკაშა მოწითალო ფერის ბუსუსებითაა დაფარული, მათ ფუძესთან განლაგებულია, მბრწყინავი წებოვანი სურნელოვანი ნივთიერების გამომყოფი სეკრეტორული უჯრედები. ფოთლის შეფერილობა და სეკრეტის სუნი იმიტირებს ყვავილს და იზიდავს მწერებს. ფოთოლზე დამჯდარი მწერი ეწებება მას და იწყებს ფართხალს. ამით მწერი აღიზიანებს მგრძნობიარე ბუსუსებს. გაღიზიანება მთელ ფოთოლზე ვრცელდება, საპასუხოდ ბუსუსები იხრებიან მწერისკენ და მჭიდროდ ეხვევიან. ბუსუსის სფერული დაბოლოებიდან გამოიყოფა პროტეოლითური (პეპსინის ტიპის) ფერმენტებისა და ორგანული მჟავების შემცველი სეკრეტი. რამდენიმე დღეს მიმდინარეობს მწერის მონელება და ფირფიტის მიერ მონელებული ნივთიერებების შეწოვა (სურ. 10.11).
ზოგიერთ მწერიჭამია მცენარეს მწერების დასაჭერად „მახე“ (მაგ., ბუშტოსანა, ვენერას ბუზიჭერია) ან „ქილა“ (მაგ., ნეპენთესი) გააჩნია. ჩრდილო ამერიკის ტორფნარში გავრცელებულ ვენერას ბუზიჭერიას (Dionaea muscipula) აქვს ორნაკვთიანი მგრძნობიარე ბუსუსებიანი შეფერილი ფოთლის ფირფიტა. მისი კიდეები იმდაგვარადაა დაკბილული, რომ დაკეცვისას ერთიმეორეში ჯდებიან. ფოთოლზე მწერის მოხვედრისას მგრძნობიარე ბუსუსები ღიზიანდება და ფირფიტა იკეცება. მწერი მახეში ებმება. ფირფიტის უჯრედების მიერ გამოყოფილი სეკრეტით მიმდინარეობს მწერის მონელება.
მწერიჭამია მცენარეები იზრდებიან მტკნარ წყალსაცავებში, დაჭაობებულ და აზატოვანი ნაერთებითა და ნაწილობრივ მინერალური მარილებით ღარიბ ნიადაგზე. ამ მცენარეებს ჰეტეროტროფული კვების გარეშეც შეუძლიათ არსებობა. ცხოველური საკვებით, დამატებითი კვების შემთხვევაში, მათი განვითარება გაცილებით უკეთ მიმდინარეობს. ამდენად სავარაუდოა, რომ ენტომოფაგიისადმი (მწერიჭამიაობა) მიდრეკილება აზოტოვანი ნივთიერებით ღარიბ გარემოში მცხოვრებ მცენარეებს ჩამოუყალიბდათ. თავდაპირველად ენტომოფაგიისადმი მიდრეკილმა ფორმებმა ნებისმიერი ორგანოთი მკვდარი ხრწნადი ნარჩენების შეწოვის უნარი შეიძინეს. მკვდარი ორგანიზმის ნარჩენების დაშლა შესაძლებელია ბაქტერიებით ან მცენარის მიერ გამოყოფილი ჰიდროლიზური ფერმენტებით განხორციელებულიყო. მცენარეებში ამ თვისების თანდათანობით გასრულყოფილებას დაერთო მწერების დაჭერის უნარის გამომუშავება. მცენარე მწერების მისატყუებლად გამოყოფილ არომატულ ნაერთებს ან შეფერილ (მეტამორფიზებულ) ორგანოებს იყენებდა. მომდევნო ეტაპზე ენტომოფაგიის უნარის სრულყოფა მწერების დამჭერი სამარჯვეების ევოლუირების გზით წარიმართა.
ექსპერიმენტულად დადგენილია, რომ მწერიჭამია მცენარეების ფერმენტული სისტემები, რომლითაც მწერებს ინელებენ ტიპურ მუტაციურ პროცესს ექვემდებარებიან. მაშასადამე, მათი სინთეზი და გამოყოფა გენეტიკური აპარატით კონტროლირდება. რითაც მტკიცდება, რომ ეს რთული ადაპტაცია ევოლუციის პროცესში ბუნებრივი გადარჩევის მოქმედებით ჩამოყალიბდა. ვარაუდობენ, რომ საწყის ეტაპზე მწერიჭამია მცენარეებს მოეპოვებოდათ პრიმიტიული მწერსაჭერი სამარჯვეები. შემდგომში მათი გასრულყოფილება და ნაირგვარ ფორმებად დივერგირება მოხდა.
|
|
12.1.8 მხედველობის ორგანოს ევოლუცია. |
▲ზევით დაბრუნება |
ერთ-ერთ ყველაზე რთულ შეგუებას წარმოადგენს მხედველობის ისეთი სრულყოფილი რთული ორგანოს ჩამოყალიბება, როგორიცაა მწერების და განსაკუთრებით ფრიველებისა და ძუძუმწოვრების თვალი. იგი ევოლუციის პროცესში უმდაბლეს ცხოველთა მარტივი მხედველობის ორგანოს თანდათანობითი გართულებით წარმოიქმნა. მხედველობის ორგანოს ევოლუცია კარგად არის შესწავლილი. გამოვლენილია ცვლილებების უწყვეტი რიგი, რომლებიც თანმიმდევრულად და მკაფიოდ გადადიან ერთმანეთში. იგი ადასტურებს, რომ ბუნებრივი გადარჩევის მოქმედებით ჩამოყალიბდა ჯერ სხვადასხვა სირთულის მარტივი და შემდეგ რთული თვალი (იხ. სურ. 10.12).
მხედველობის ორგანოები შედგება ნერვული ნაწილისაგან (შეიცავს ფოტორეცეპტორულ უჯრედებს) და ნაწილისაგან, რომელიც უზრუნველყოფს სინათლის სხივების მოხვედრას რეცეპტორულ ნაწილზე. ისტორიული განვითარების პროცესში თანდათანობით ევოლუირებდა ორივე ნაწილი (ნათაძე, 1962).
პროტისტების ზოგიერთ წარმომადგენელს (გვარი უგლენა, Chlamydomonas...) წინა ბოლოში მოეპოვებათ წითელი სხეულაკი - სტიგმა, რომლითაც აღიქვამენ სინათლით გაღიზიანებას. მისი თანაპოვნიერება ფოტოტაქსისს განაპირობებს.
მრავალუჯრედიან ცხოველებში სინათლის რეცეფცია უმარტივესი სახით ხორციელდება დიფუზურად გაფანტული ფოტორეცეპტორული უჯრედებით. ზოგიერთ ცხოველში იზოლირებული რეცეპტორული უჯრედები ეპიდერმისში (მაგ., ჭიაყელა) ან ნერვულ ქსოვილშია (მაგ., ლანცეტა) დიფუზურად გაფანტული. ისინი მხოლოდ განათების ინტენსიურობას საზღვრავენ, კერძოდ მეტად განათებულ ადგილს ასხვავებენ ნაკლებად განათებულისგან.
მხედველობის ორგანოს განვითარების პირველ ეტაპზე დიფუზურად გაფანტული უჯრედების ჯგუფებად თავმოყრით და გაერთიანებით ყალიბდება მარტივი თვალი. მაგალითად, ზოგიერთ მედუზასა და ტურბელარიაში ფოტორეცეპტორული უჯრედების თავმოყრით უყალიბდებათ თვალის ლაქა (ე.წ. სტიგმა). პლანარიაში (Planaria gonocephala) პიგმენტური უჯრედები წარმოქმნის თასის ფორმის სტრუქტურას. მასში თავმოყრილია რეცეპტორული უჯრედები, რომელიც დაცულია სინათლის გვერდითი სხივების ზემოქმედებისაგან. ამდაგვარი თვალის მფლობელ ინდივიდებს გააჩნიათ ე.წ. ფოტოსკოპური მხედველობა (განარჩევენ სინათლის ინტენსიურობას და დაცემული სხივების მიმართულებას).
მხედველობის ორგანოს განვითარების მომდევნო ეტაპზე ფოტორეცეპტორული უჯრედების მფლობელი ეპიდერმული უბნის თვალის ლაქის ჩაზნექვის შედეგად წარმოიქმნა ფოსო. მის ფსკერზე განთავსებულია ფოტორეცეპტორული უჯრედები (მაგ., ზოგიერთი მედუზა, და მოლუსკი). იგი უკეთაა დაცული სხვადასხვა ფაქტორების ზემოქმედებისაგან. ამდაგვარი თვალით გამოსახულების მიღება შეუძლებელია, ვინაიდან ფოსოს გააჩნია ფართო ხვრელი, რის გამოც საგნის ერთი უბნიდან არეკლილი სხივებით რამდენიმე ფოტორეცეპტორი ღიზიანდება, ხოლო რამდენიმე უბნიდან წამოსული სხივებით - ერთი და იგივე სენსორული უჯრედი.
მხედველობის ორგანოს განვითარების მნიშვნელოვან ეტაპს წარმოადგენს მარტივი კამერიანი თვალის ჩამოყალიბება. ფოსოს ხვრელის კიდეების ერთმანეთთან დაახლოებისა და შეერთებით ჩამოყალიბდა ბუშტუკიანი (მარტივი კამერიანი) თვალი. ბუშტუკის ფსკერზე განფენილია პიგმენტური და სენსორული უჯრედები (წარმოიქმნა რეტინა ანუ ბადურა), ხოლო ღრუ ბლანტი სითხითაა ამოვსებული (მინისებრი სხეული). ამ ტიპის თვალი მოეპოვება ზოგიერთ მრავალჯაგრიან ჭიას, მუცელფეხიან და თავფეხიან მოლუსკს.
ბუშტუკიანი თვალის ინვაგინაციით ჩამოყალიბდა ორშრიანი თასის ფორმის თვალი. წინა ნაწილში ვიწრო ხვრელის წინ ბროლი (ლინზა) განთავსდა. კამერიანი თვალის შემდგომი სრულყოფისა და გართულების შედეგად ჩამოყალიბდა რამდენიმე შრიანი ბადურა, რქოვანა, გუგის შემცველი ფერადი გარსი. ამდაგვარი რთული თვალი მოეპოვებათ თავფეხიან მოლუსკებსა და ხერხემლიანებს. კამერიანი თვალი უზრუნველყოფს საგნის ხატის შეგრძნებას ე.წ. ეიდოსკუპურ მხედველობას. აკომოდაცია (ფოკუსის მანძილის შეცვლა) თევზებსა და თავფეხიან მოლუსკებში ბროლის მოძრაობით ხორციელდება, ხმელეთის ხერხემლიანებში კი ბროლის სიმრუდის რადიუსის ცვლილებით.
თავფეხიან მოლუსკებსა და ხერხემლიანების თვალი ანალოგიური ორგანოებია, აგებულებით მსგავსია, მაგრამ წარმოშობით კი განსხვავდებიან. ამ არამონათესავე ცხოველებს მსგავსი მხედველობის ორგანოები კონვერგენეტული ევოლუციის შედეგად ჩამოუყალიბდათ.
მარტივი თვალის გართულების განსხვავებული გზით არის მიღებული მწერების, კიბოსნაირებისა და ზოგიერთი რგოლოვანი ჭიის რთული ფასეტური თვალი. იგი რთული ორგანიზაციის მარტივი თვალების ე.წ. ომატიდიუმების გაერთიანების შედეგად არის მიღებული. თითოეული ომატიდიუმის ზედაპირი აღჭურვილია ამოზნექილი ექვსწახნაგის - ფასეტის ფორმის უძრავი რქოვანი ლინზით. ომატიდიუმში მოთავსებულია რეტინულა (სენსორულ უჯრედთა ერთობლიობა). იგი არაეფექტური მხედველობის ორგანოა. გარჩევის უნარი გაძლიერებულია მარტივი თვალის რაოდენობის გაზრდით. დროზოფილას მოეპოვება 700, ფუტკარს 6-7 ათასამდე, ნემსიყლაპიას 28 ათასამდე ომატიდიუმი. ფასეტური თვალი დეტალების დაკონკრეტების გარეშე საგნის მოზაიკურ გამოსახულებას იძლევა.
სურ. 10.12. მხედველობის ორგანოს ევოლუცია მოლუსკებში. ევოლუციის პროცესში თანდათანობით კამერიანი თვალის ჩამოყალიბება. 1 - თვალის ლაქა (სტიგმა); 2 და 3 - თასისებრი ფორმის თვალი; 4 და 5 - ბუშტუკიანი (კამერიანი) თვალი ბროლის გარეშე, 6 - ბროლის მქონე კამერიანი თვალი (ი. შმალგაუზენი, 1990).
ამგვარად, შეიძლება დავასკვნათ, რომ ცხოველებში მარტივი თვალის ევოლუცია ორი განსხვავებული გზით წარიმართა, რის შედეგადაც ჩამოყალიბდა, ერთი მხრივ, რთული ფასეტური თვალი, მეორე მხრივ კი, რთული კამერიანი თვალი.
თანამედროვე გენეტიკის მონაცემებით დადასტურებულია, რომ ონტოგენეზში თვალის ჩამოყალიბება გენეტიკური კონტროლით ხორციელდება. დროზოფილაში ინდუცირებულია ნაირგვარი მუტაცია, რომელიც თვალის ფორმის (ომატიდიუმების რაოდენობის) და შეფერილობის ცვლილებას ან უთვალობას იწვევს. ადამიანში ცნობილია მრავალი სპონტანური მუტაცია, რომელიც მხედველობის ორგანოს ნაირგვარ პათოლოგიურ მოქმედების ცვლილებას იწვევს.
ბუნებრივი გადარჩევის სანიმუშო დამამტკიცებელი არგუმენტია თვალის გამარტივება ან რედუქცია მიწაში, მღვიმეებში და ღრმა წყლებში ბინადარ ცხოველებში. ასე მაგალითად, მიწაში მცხოვრებ ჩანთოსან თხუნელას, თვალი საერთოდ არ გააჩნია. ბრუცასა და თხუნელაში რედუცირებული თვალი კანითაა დაფარული და არ ფუნქციონირებს. ამ შემთხვევაში ცხოველებში სწრაფად გროვდება მადეზინტეგრირებელი მუტაციები, რომლებიც ადრე მასტაბილიზებელი გადარჩევით იცხრილებოდა.
|
|
12.1.9 ზოოფილური მცენარეები. |
▲ზევით დაბრუნება |
ჯვარედინმტვერია ყვავილოვან მცენარეებს დამტვერვის ნაირგვარი სამარჯვეები გამოუმუშავდათ. განსაკუთრებით რთული ადაპტაციები მოეპოვებათ ზოოფილურ მცენარეებს. დამტვერავი ცხოველების მიხედვით გამოყოფენ ენტომოფილურ, ორნითოფილურ და ტერიოფილურ მცენარეებს. როგორც ყვავილებს ისე მათ დამტვერავ ცხოველებს განსაცვიფრებელი შეგუებები გააჩნიათ, რომლის ჩამოყალიბებაც მხოლოდ ბუნებრივი გადარჩევის მოქმედებით შეიძლება აიხსნას.
ანემოფილური და ჰიდროფილური მცენარეებისგან განსხვავებით ზოოფილურებში მტვრის მარცვლის გადატანა ერთი ყვავილიდან დანარჩენზე გაცილებით ეფექტურია. ამიტომაც მათში მტვერი მცირე რაოდენობით წარმოიქმნება. ნაირგვარი ადაპტაციები ერთ მთავარ სტრატეგიას ემსახურება: მცენარეებში მცირე რაოდენობით წარმოქმნას მტვერი და მოხდეს მისი ეფექტური გამოყენება. ამდაგვარი სტრატეგიის შედეგად ზოოფილური მცენარეები სხვებთან შედარებით მომჭირნეობით ხარჯავენ ცხოველმყოფელობისთვის მნიშვნელოვან ნივთიერებებს (ცილები, ნახშირწყლები, ნუკლეინის მჟავები და მისთ.). და ზოგავენ მათი სინთეზისათვის საჭირო დიდძალ ენერგიას.
ზოოფილურ მცენარეებსა და მათი დამტვერავი ცხოველების შეუღლებული ევოლუირებით ჩამოყალიბდა ისეთი მაკორდინირებელი ადაპტაციები, რამაც განაპირობა ერთის მხრივ მცენარეთა საიმედო ჯვარედინი დამტვერვა, მეორეს მხრივ ცხოველებში ყუთიანი საკვების მოპოვება. ზოგიერთი ფიტოფაგი ვიწროდაა სპეციალიზებული, მხოლოდ ნექტარითა და მტვერით იკვებება.

სურ. 10.13. სალბის ყვავილი და მისი დამტვერავი მწერი A - ყვავილის ჭრილი: 1 - ბუტკო; 2 - მტვრიანა; B - ყვავილი რომელშიც მწერია ჩამძვრალი; C - მტვრიანები ნორმაში; D - მტვრიანები დაშვებულ მდგომარეობაში (ვ. ლუნკევიჩი, 1974).
ცხოველებში რთული კამერიანი და ფასეტური თვალის წარმოქმნამ, ფერთა გარჩევის უნარმა ზოოფილურ მცენარეებში ნაირგვარი ფორმისა და შეფერილობის ყვავილების ჩამოყალიბება განაპირობა.
ყვავილებისა და მათი დამტვერავი ცხოველების შეუღლებული ევოლუირების შედეგად ჩამოყალიბებული რთული ადაპტაციების საილუსტრაციოდ რამდენიმე მაგალითს განვიხილავთ. ზოგჯერ მწერისა და ყვავილის პროპორციები ურთიერთისადმია მისადაგებული (იხ. §8.8). ასეთი ყვავილი მწერთა ვიწრო ჯგუფით იმტვერება. მაგალითად, სალბს (შალვია) მხოლოდ ფუტკარი, ბაზი ან კრაზანა მტვერავს. ყვავილში გვირგვინის ფურცლების ქვედა ნაწილი მილად არის შეზრდილი, ზედა ნაწილი კი ორტუჩაა. სანექტრეები გვირგვინის ფსკერზე ღრმადაა მოთავსებული. მას ფარავს მტვრიანის ფუძის ფირფიტისებურად გაფართოებული ნაწილი. რკალისებურად მოხრილი მოძრავი გრძელი მტვრიანები ზედა ტუჩში ბოლოვდება. გვირგვინში შეღწევა და ნექტრის მოპოვება მხოლოდ გარკვეული ზომის (უნდა მიესადაგებოდეს გვირგვინის მილის დიამეტრს), მძიმე, ძლიერ მწერს შეუძლია. ბაზი როდესაც ქვედა ტუჩზე ჯდება, სიმძიმის გამო ტუჩთაშორისი ხვრელი ფართოვდება. მწერი ძვრება გვირგვინში. იგი თავით აწვება ფირფიტებს და ცდილობს ხორთუმით ჩაწვდეს სანექტრეებს. ფირფიტებზე ძლიერი დაწოლის გამო მტვრიანები ქვევით ეშვება. იგი მწერს ზურგზე ეცემა და მტვერი ეყრება. როდესაც ბაზი ადრე გაშლილ ყვავილზე გადადის (ხნოვანებასთან ერთად დინგი გრძელდება) ამჟამად მის ზურგს ორად გაყოფილი დინგი ეხება და იმტვერება (იხ. სურ. 10.13).
კვებითი ინსტინქტების გარდა მცენარე ზოგჯერ მისაზიდად მწერის სქესობრივ ინსტინქტებს იყენებს. მაგალითად, ჯადვარის (Ophrys insectifera) უნექტრო ყვავილი ფორმით, შეფერვითა და სუნით მდედრ კრაზანას (Apgogorytes mystaceus) იმიტირებს. მსგავსება იმდენად სრულყოფილია, რომ მამრი კრაზანა ცდილობს ყვავილთან კოპულირებას. რხევისას მწერის სხეულიდან მტვერი ბუტკოს ეყრება, ხოლო იგი სამტვრიდან გადმოყრილი მტვრით იგანგლება. გაწბილებული კრაზანა ტოვებს ყვავილს და სხვა „მდედრისკენ“ - ყვავილისკენ მიეშურება.
ზოგიერთი მწერი ყვავილს (იუკა, ლეღვი და მისთ.) გასამრავლებლად იყენებს. მაგალითად, იუკა და მისი დამტვერავი მწერი პრონუბა (Pronuba insectifera) ერთმანეთის გარეშე ვერ მრავლდებიან. მდედრი მწერი ღამით მიფრინდება იუკას ყვავილებთან. ძვრება შიგ და სახეცვლილი ყბებით მტვერს გუნდად აგროვებს, რომელიც გადააქვს სხვა ყვავილზე. პრონუბა კვერცხსადებით ხვრეტს ნასკვს და ყრის კვერცხებს. სვეტის გავლით აცოცდება დინგზე და მის ღრმულში ტენის მტვრის გუნდას. ამრიგად, მწერში ჩამოყალიბებული რთული ქცევით უზრუნველყოფილია, როგორც შთამომავლობის საიმედო დაცვა და განვითარება, ისე ყვავილის დამტვერვა და განაყოფიერება. კვერცხებიდან გამოსული მატლები საკვებად ნორჩი თესლების გარკვეულ ნაწილს იყენებენ, ნაწილი კი მცენარის გამრავლებას ხმარდება. უნდა აღინიშნოს, რომ იქ სადაც პრობუნა არ გვხვდება იუკა თესლს არ იკეთებს.
სრულიად განსხვავებული ადაპტაციები ჩამოუყალიბდათ ორნითოფილურ მცენარეებს. ფრინველები მხედველობით ორიენტირებენ. ამიტომაც ორნითოფილური მცენარეების ყვავილი უსუნოა, მაგრამ მოეპოვება კაშკაშა წითელი, ლურჯი ან მწვანე (სინათლის სპექტრის ძირითადი ფერები) შეფერილობა. მათ მტვერავენ მცირე ზომის ფრინველები (კოლიბრი, მეთაფლია), ფერის გარდა ფრინველს ყვავილის ფორმაც იზიდავს. ყვავილი მტკიცეა, რომელსაც კარგად განვითარებული მექანიკური ქსოვილი ან ტურგორი იწვევს. იგი უხვად გამოყოფს თხევად ან ლორწოვან ნექტარს. საკვების მოპოვებაში ფრინველებს ეხმარება გრძელი ნისკარტი და ენა (სურ. 10.14).

სურ. 10.14. ორნითოფილური ყვავილს მტვერავს კოლიბრი (ა. პონომარიოვი, 1980).
ტერიოფილური ყვავილები დასამტვერად ძუძუმწოვრებს, მეტწილად ხელთფრთიანებს იზიდავენ. საამისოდ ისინი სპეციფიკურ ადაპტაციებს ფლობენ. ყვავილი განსაკუთრებული სიმტკიცით გამოირჩევა, რომელსაც ღამურები ნექტრის მოპოვებისას ვერ აზიანებენ. ღამურები სარჩოს მოსაპოვებლად ღამით გამოდიან, ამიტომაც გააჩნიათ კარგი ყნოსვა და ცუდი მხედველობა. ტერიოფილური ყვავილები ღამის საათებში გამოყოფენ ნაირგვარ სურნელოვან ნივთიერებებს. ღამურები სუნით პოულობენ მცენარეებს. ისინი იკეთებენ დიდ, ღია ფერის ყვავილებს, რომლებიც ღამით უხვად გამოყოფენ ნექტარს. გარკვეული ადაპტაციები ღამურებსაც გააჩნიათ, კერძოდ მათი დინგი წაგრძელებულია და ენა საკმაოდ გრძელია.
ამრიგად, ზოოფილური ყვავილების შეფერილობა და აგებულება, სპეციფიკური სუნის თანაპოვნიერება, ნექტრის გამოყოფის დრო საოცრად თანხვდება ცხოველის სარჩოს მოპოვების დროს, ასევე შესაბამისობაშია სხეულის აგებულება და ქცევითი რეაქციები. ადაპტაციები განსაკუთრებით მკვეთრად გამოხატული და სპეციფიკურია როდესაც ყვავილის ნექტრით ცხოველთა ვიწრო ჯგუფი იკვებება. იმ შემთხვევაში, თუ ნექტრით ნაირგვარი ცხოველები იკვებებიან, მაშინ ადაპტაციები ზოგადი და უნივერსალური ხასიათისაა.
|
|
12.1.10 ბიოქიმიური და ფიზიოლოგიური ადაპტაციები |
▲ზევით დაბრუნება |
ამ ტიპის ადაპტაციები მრავალგვარია. ადაპტაციათა მთავარი სტრატეგიაა მისი მფლობელი ინდივიდის გადარჩენა და წარმატებული გამრავლება. მნიშვნელოვან ადაპტაციას წარმოადგენს გენეტიკური კოდის ტრიპლეტური ბუნება და გადაგვარებული ხასიათი (ამინომჟავა კოდირდება ერთზე მეტი ტრიპლეტით), რაც მინიმუმამდე ამცირებს ინდუცირებულ სპონტანურ მუტაციების სიხშირეს (იხ. §13). შეიძლება დავასახელოთ დნმ-ის რეპარაციის განსხვავებული მექანიზმების (ფოტორეაქტივაცია, ეკსციზური და პოსტრეპლიკაციური რეპარაცია და მისთ.) არსებობა. ჩამოთვლილი ადაპტაციები უზრუნველყოფენ მემკვიდრული მასალის სტაბილობასა და შთამომავლებში საიმედო გადაცემას.
მაღალი ტემპერატურისა და ჟანგბადის სიჭარბის პირობებში ტროპიკულ მცენარეებს ფოტოსინთეზში ნახშირორჟანგის ფიქსაციის (სიბნელის ფაზა) ახალი C4 - გზა ჩამოუყალიბდათ (პეტჩ-სლეიკის ციკლი). ტროპიკულ მცენარეებს შეუნარჩუნდათ C3 - გზაც (კალვინის ციკლი), მაგრამ იგი არაეფექტურია. ბიოქიმიური ადაპტაციის სანიმუშო მაგალითია ალელოპათიის ფენომენი. მცენარე გამოყოფილი სპეციფიკური ნივთიერებით იზიდავს სასარგებლო კონსორტებს და აფრთხობს მავნეს, რაც უზრუნველყოფს ინდივიდის წარმატებულ კონკურენციას კონკრეტულ გარემოში. მაგალითად, ჩრდილოეთ ამერიკაში გავრცელებული შავი კაკლის (Jugland nigra) ფესვი, ღერო და ფოთოლი გამოყოფს ტოქსინ იუგლონს, რომელიც თრგუნავს ბალახოვანი მცენარეების (პომიდორი, კარტოფილი, იონჯა და მისთ.) განვითარებას.
ზოგიერთი მცენარე გამოიმუშავებს ტოქსინს (ნიკოტინი, პირეტრინი, კოლხიცინი, ატროპინი, რიცინი, მორფინი, ციანოგენური გლიკოზიდები და მისთ.) ან უსიამოვნო გემოს ან სუნის ნივთიერებას, რითაც თავს იცავს ფიტოფაგებისაგან. გამოვლენილია 800-მდე სახეობის ყვავილოვანი მცენარე, მათ შორის სელი, თეთრი სამყურა, ნუში, რომლებიც ციანოგენური გლიკოზიდებს შეიცავენ. ეს ნივთიერება უშუალოდ არატოქსიკურია, მაგრამ ფიტოფაგის ორგანიზმში მოხვედრისას ენზიმური დაშლის შედეგად მიიღება ძლიერი ტოქსიკური ნივთიერება - ციანმჟავა. იგი ცხოველის მოწამვლას იწვევს, ჭარბი რაოდენობით მიღებისას კი სიკვდილს.
ტოქსიკური აგენტისადმი ტოლერანტული ფიტოფაგი (ძირითადად მწერები) ტოქსინს თავდასაცავადაც კი იყენებს. მაგალითად, დანაიდების ოჯახში გაერთიანებული პეპლების მუხლუხები იკვებებიან გულის გლიკოზიდის შემცველი რძიანა მცენარეებით. მუხლუხების ორგანიზმში დიდი რაოდენობით აკუმულირდება ტოქსინი, რომელიც მოდიფიცირების გარეშე გადადის პეპლებში. ისინი აპოსემიური შეფერილობისანი არიან. როდესაც გამოუცდელი ფრინველი (ძირითადად ჩხიკვი, შაშვი) ჭამს პეპელას, გლიკოზიდების მოქმედებით ეწყება ღებინება. ერთი ან ორი სინჯით ფრინველებს გამღიზიანებელზე (ე.ი. პეპლებზე) უარყოფითი კვებითი რეაქცია უყალიბდებათ. ისინი ერიდებიან არა მარტო დანაიდებს, არამედ მოხატულობით მათ მსგავს პეპლებსაც (იხ. მიმიკრია).
მცენარეებს და ცხოველებს ჩამოუყალიბდათ ნაირგვარი ადაპტაცია, რომლითაც წყლის დეფიციტს, ასევე მაღალი ან დაბალი ტემპერატურის უარყოფით ზემოქმედებას ადვილად იტანენ. პოიკილოთერმულ ცხოველებს თერმორეგულაციის ნაირგვარი სამარჯვეები მოეპოვებათ. ისინი სითბურ ენერგიას ძირითადად მზის სხივური ენერგიით იღებენ. ზოგიერთი ცხოველი თერმორეგულაციას სხეულის შეფერილობის ცვლილებით ახორციელებს. მაგალითად უდაბნოს იგუანა (Dipsosaurus dorsalis) ადრე დილით მუქი შეფერილობისაა (გარემოში დაბალია ტემპერატურა. მუქი სხეული მეტ სხივურ ენერგიას შთანთქავს ვიდრე ღია). შუადღისას ღია შეფერილობისაა და გაცილებით ნაკლებ სითბურ ენერგიას იღებს გარემოდან.
ევოლუციის პროცესში ცხოველებს სამი ძირითადი სახის ადაპტური ქცევები ჩამოუყალიბდათ: საკვებმოპოვებითი, თავდაცვითი და გამრავლებითი. თავდაცვითი ქცევების კარგი მაგალითია ზემოთ განხილული დემონსტრაციული და კრიპტული ქცევები. ზოგიერთი საკვებმოპოვებითი ქცევა ხელს უწყობს ახალი საკვები რესურსების ათვისებას და კვებითი ჯაჭვის გაფართოებას. მაგალითად ჯგუფურ ცხოველებს, კერძოდ, ვირთაგვებს მათთვის უცხო საკვების ათვისებისას გარკვეული სტრატეგია ჩამოუყალიბდათ. უცხო საკვებს ჯგუფის მხოლოდ ერთი წევრი ჭამს. დანარჩენები კი აკვირდებიან მის მდგომარეობას. თუ საკვებმა უარყოფითი ფიზიოლოგიური რეაქცია არ გამოიწვია ცხოველში, მხოლოდ მას შემდეგ იწყებენ ჯგუფის სხვა წევრები კვებას. მოყვანილი მაგალითი მეტყველებს ქცევის ორგანიზებულობაზე, რომლის სტრატეგიაა ჯგუფის დაცვა უცხო საკვების მავნე ზემოქმედებისაგან.
ევოლუციური თვალთახედვით მნიშვნელოვანია გამრავლებითი (გამრავლებასთან დაკავშირებული) ქცევების მრავალფეროვნება. პოპულაციები, რომელთაც მრავალფეროვანი გამრავლებითი ქცევები გააჩნიათ „რეპროდუქციულ უპირატესობას“ ფლობენ სხვებთან შედარებით. სანიმუშოდ მოვიყვანთ ერთ მაგალითს. მექსიკური ჩხიკვი, მობუდარი ფრინველია. მუხნარში ან ფიჭვნარში გუნდებად (4-15 ინდივიდი) ბინადრობს და კოლექტიურად იცავენ დაკავებულ ტერიტორიას სხვა გუნდებისაგან. ბუდობს 1-4 წყვილი. გუნდის სხვა წევრები, ძირითადად ახალგაზრდები, მშობლებს ბარტყების გამოკვებაში ეხმარებიან. ამდაგვარი კოოპერაციული ქცევები სასარგებლოა მთლიანად გუნდისათვის. ახალგაზრდა ფრინველები იძენენ ბარტყების აღზრდის ჩვევებს. მშობლის სიკვდილის შემთხვევაში ბარტყები არ იღუპებიან შიმშილით. გუნდი რაციონალურად ითვისებს საკვებ რესურსებს და არეგულირებს დაკავებულ ტერიტორიაზე სიმჭიდროვეს. კოოპერაციული თანაცხოვრებით მიიღწევა გადარჩენისა და შთამომავლობის დატოვების მეტი ალბათობა, მარტოულად მცხოვრებ ფორმებთან შედარებით. ქართველ მკითხველს ამ პრობლემაზე ვრცელი და ამომწურავი ინფორმაციის მიღება შეუძლია ს. ცაგარელის სახელმძღვანელოდან „ბიოსოციოლოგია“ (2004).
|
|
12.2 10.2. ადაპტაციის კლასიფიკაცია ა წარმოქმნის მექანიზმები |
▲ზევით დაბრუნება |
|
|
12.2.1 10.2.1. ადაპტაციის კლასიფიკაცია |
▲ზევით დაბრუნება |
ევოლუციური თვალთახედვით სხვადასხვა ადაპტაციის მრავალფეროვნების ემპირიული აღწერა ნაკლებმნიშვნელოვანია. არსებითია მრავალფეროვანი და მრავალგვარი ადაპტაციების დაჯგუფება (კლასიფიცირება) და მათი წარმოქმნის მექანიზმების დადგენა.
ადაპტაციას აჯგუფებენ წარმოშობის, სხვადასხვა გარემოს ასპექტებისადმი კუთვნილების და მასშტაბის მიხედვით.
წარმოშობის გზების მიხედვით გამოყოფენ პრეადაპტაციურ, კომბინაციურ და პოსტადაპტაციურ ადაპტაციებს. პრეადაპტაციის გზის დროს პოტენციური ადაპტაციური ცვლილების წარმოქმნა ხდება გაცილებით ადრე, მოგვიანებით კი ხორციელდება მისი რეალიზაცია შეცვლილ საარსებო გარემოში. მუტაციური პროცესისა და შეჯვარების (რეკომბინაციის) მეშვეობით სამობილიზაციო რეზერვი იქმნება (იხ. §4.). ახალ საარსებო გარემოში შესაგუებლად ხდება მათი ნაწილის გამოყენება, ახალი შეგუების ჩამოსაყალიბებლად. გენოფონდში ფარულად არსებული ადრე წარმოქმნილი მუტაციის ადაპტაციურ ნიშნად გარდაქმნის კარგი მაგალითია ჩვენ მიერ ზემოთ განხილული ინდუსტრიული მელანიზმის მოვლენა (იხ. §84.) ბაქტერიაში ფაგისადმი რეზისტენტულობის განმსაზღვრელი გენის ინდუქცია ადრე ხდება, ხოლო მისი რეალიზაცია შესაბამის გარემოში (ფაგით დასნებოვნებისას) მიმდინარეობს. იმავე მოვლენას ვხედავთ ბაქტერიაში ანტიბიოტიკებისადმი რეზისტენტული ნიშნის ჩამოყალიბების დროს. ანტიბიოტიკებიან არეში მოხვედრისას გადარჩებიან და შთამომავლობას ტოვებენ მხოლოდ ის ორგანიზმები, რომლებიც რეზისტენტულობის განმსაზღვრელ გენებს შეიცავენ.
პრეადაპტაციის გზის დროს ადრე ჩამოყალიბებული ორგანიზმის თავისებურებები, საჭიროა სრულიად სხვა პირობებისათვის, იგი წარმატებულად გამოიყენება ახალ გარემოში შესაგუებლად. იქმნება შთაბეჭდილება, თითქოს ზოგიერთი რთული შეგუების წარმოქმნა წინ უსწრებდა დროში საადაპტაციო პირობებს. ასე მაგალითად, ძუძუმწოვრებში ქალას ნაკერები აიოლებს მშობიარობას, მაგრამ მისი წარმოქმნა ცოცხალმშობიარობას არ უკავშირდება.
ახალი მუტაციების ერთმანეთთან და გენოტიპთან ურთიერთქმედების გზით ყალიბდება კომბინაციური ადაპტაციები. მუტაციის ეფექტი დამოკიდებულია იმ გენოტიპურ გარემოზე, რომელშიც ახალი მუტანტური ალელები აღმოჩნდებიან. შეჯვარების მეშვეობით მუტანტური ალელების შემცველ გენოტიპთა ნაირგვარი ვარიანტები - კომბინაციები მიიღება. სხვადასხვა გენებთან დამოკიდებულების კვალობაზე მუტანტური ალელების ფენოტიპური გამოვლენის ეფექტი გენოტიპურ ვარიანტებს განსხვავებული ექნებათ. შესაძლებელია, რომ ნიშნის ფენოტიპური გამოვლენა გაძლიერდეს (კომპლემენტაცია) ან დაითრგუნოს (ეპისტაზი), ან მრავალი ადიტიური გენის მოქმედებით გრადუირებული გახდეს (პოლიმერია). ყველა შემთხვევაში ერთი ადაპტაციის მეორეთი სწრაფად შეცვლის რეალური შესაძლებლობა იქმნება. მიაჩნიათ, რომ ადაპტაციის ჩამოყალიბების ეს გზა ყველაზე ფართოდაა გავრცელებული ბუნებაში.
პოსტადაპტაციური გზით ადაპტაციის წარმოქმნის შემთხვევაში ძველი ნიშნის რედუქცია ხდება. ზოგჯერ ადრე ჩამოყალიბებული ნიშან-თვისებები სრულიად სხვა მიზნით გამოიყენება. ამ ნიშნის განმსაზღვრელი გენები რეცესიულ მდგომარეობაში გადადიან. ისინი გენოფონდის სამობილიზაციო რეზერვში ჩაერთვებიან. ხდება გენების შენარჩუნება პოპულაციაში, რომლებიც დრო და დრო ფენოტიპურად ატავისტური ნიშნის სახით ვლინდებიან. თუ გენები ახალ საარსებო გარემოში სასარგებლო აღმოჩნდება, მაშინ ისინი ახალი ადაპტაციური ნიშან-თვისების განვითარებას იწვევენ. ამრიგად, ადრე ადაპტაციური მნიშვნელობის მქონე ნიშანთა ნაწილი, რომელთაც ადაპტაციური მნიშვნელობა დაკარგეს, შესაძლოა ახალ გარემოში კვლავ ადაპტაციური აღმოჩნდეს. მეტწილად არაადაპტაციური ღირებულების მქონე ნიშნები რუდიმენტების სახით შემორჩებიან და ევოლუციის პროცესში იკარგებიან.
პოსტადაპტაციური გზის შემთხვევაში ახალი ადაპტაციები ადრე ჩამოყალიბებული სტრუქტურის გასრულყოფილებისა და ფუნქციის შეცვლის შედეგად წარმოიქმნება. ასე მაგალითად, ხერხემლიანთა წინაპრებს ვისცერალური ჩონჩხი (სპლანქნოკრანიუმი) მრავალი დაუნაწევრებელი სალაყუჩე რკალისაგან ჰქონდათ აგებული და, ძირითადად, სუნთქვით ფუნქციას ემსახურებოდნენ. იგი, ამასთანავე, საჭმლის მომნელებელი მილის წინა ნაწილის დაჭერის ფუნქციასაც ითავსებდა. ევოლუციის პროცესში სასუნთქი მოტორიკის გასრულყოფილებამ, მოითხოვა სალაყუჩე აპარატის მოძრაობა, რასაც თან მოჰყვა სალაყუჩე რკალების რიცხვის შემცირება და ერთიანი რკალების მოძრავ ელემენტებად დაყოფა. იგი წყლის საჭირხნი სისტემის ნაწილი გახდა. შემდგომი ევოლუციისას წინა რკალთაგანმა შეიცვალა ფუნქცია და მიტაცების იარაღად იქცა, მოგვიანებით კი ყბებად ჩამოყალიბდა (ნათაძე, 1962).
ადაპტაციები სხვადასხვა ბიოტურ გარემოში. სხვადასხვა ბიოტური გარემოსადმი კუთვნილების მიხედვით ადაპტაციები განსხვავებულია. ბუნებრივი გადარჩევის ნებისმიერი შედეგი ბიოტურ (ორგანიზაციის დონის) ცვლილებასთან არის დაკავშირებული. ადაპტაცია ამა თუ იმ ბიოტური ცვლილების კვალობაზე ყალიბდება. ორგანიზაციის დონის შესაბამისად შეიძლება დაიყოს გენოტიპურ, უჯრედულ, ორგანიზმულ და ეკოსისტემურ გარემოში მოქმედ ადაპტაციებად.
მოლეკულურ-გენეტიკურ დონეზე ადაპტაციებით ყალიბდება გენოტიპი, როგორც ერთიანი და რთული სისტემა; იგი განსაზღვრავს გენთა დომინირების ხასიათს და კოადაპტაციის (ურთიერთშეგუების) განვითარებას. ამ დონეზე შეინიშნება მოლეკულათა ადაპტური ორგანიზაცია და მათი ურთიერთქმედება. იგი განაპირობებს მოლეკულათა ავტორეპროდუქციასა და ბიოპოლიმერების კონსტრუირებას.
უჯრედულ დონეზე შეინიშნება მრავალი ორგანოიდის რთული სტრუქტურული ორგანიზაცია და მულტიფუნქციონირება, რის შედეგადაც უჯრედი რთულ მორფო-ფიზიოლოგიურ სისტემას იძენს.
ინდივიდის დონეზე ადაპტაციები ონტოგენეზთანაა დაკავშირებული. დროსა და სივრცეში მოწესრიგებულად მიმდინარეობს მემკვიდრული მასალის რეალიზაციის პროცესი ე.ი. ხორციელდება მორფოგენეზი. ორგანიზაციის ამ დონეზე ორგანიზმს კოადაპტაციის ნაირგვარი ფორმები ჩამოუყალიბდა. ადაპტაციები შეიძლება იყოს მორფოლოგიური, ფიზიოლოგიური და ეთოლოგიური (მრავალი მაგალითი ზემოთ უკვე განვიხილეთ).
კონკრეტული ინდივიდი ცოცხლობს და მრავლდება (არსებობს კონკრეტულ დროსა და სივრცეში) პოპულაციის ფარგლებში. პოპულაციურ გარემოში ყალიბდება პოპულაციურ-სახეობრივი ადაპტაციები. მათ მიეკუთვნება პოპულაციის ჰეტეროგენულობა, შიდაგენეტიკური პოლიმორფიზმი, სქესობრივი პროცესი, სიმჭიდროვე და მისთ.
ბიოცენოზში სახეობათა შორის ურთიერთდამოკიდებულება მეტად რთული და მრავალფეროვანია. ადაპტაციის შედეგად ბიოცენოზში ჩამოყალიბდა ორგანიზმთაშორის ურთიერთდამოკიდებულების ისეთი ფორმები როგორიცაა: კონსორციები, სიმბიოზი, კონკურენცია, ალელოპათია და მისთ.
მასშტაბის მიხედვით გამოყოფენ: 1. სპეციალიზებულ ადაპტაციებს, რომელიც სასარგებლოა ლოკალურ საარსებო პირობებში. მაგალითად, ენის აგებულება ჭიანჭველაჭამიებში, გრძელი ენა და მოკლე სატეხისებური ფორმის ნისკარტი კოდალაში, სამ-სამი თითით საკიდურა კიდურები ზარმაცაში, რითაც ხეზე ცხოვრების ნირთან არის შეგუებული; 2. ზოგად ადაპტაციებს, რომელიც ნაირგვარ გარემო პირობებში ფართო სპექტრისთვისაა სასარგებლო და მოეპოვებათ დიდ ტაქსონებს. ასე მაგალითად, ცხოველებში: თევზის სხეულის ფორმა, ფრინველის ფრთა, ხერხემლიანების გულსისხლძარღვთა, სუნთქვისა და ნერვულ სისტემებში მომხდარი მორფო-ფიზიოლოგიური ცვლილებები, ძუძუმწოვრებში ცოცხალმშობიარობა; მცენარეებში: ფოტოსინთეზისა და აერობული სუნთქვის მექანიზმები. უმაღლეს მცენარეებში თესლით გამრავლება და გამეტოფიტის რედუქცია, რამაც გამოიწვია მათი ევოლუციური „გარღვევა“ ახალ ადაპტურ ზონაში.
ზოგადი ადაპტაცია თავდაპირველად კერძო შეგუების სახით ჩნდება და მხოლოდ შემდეგ იძენს უნიკალურ მნიშვნელობას. სპეციალიზებული ადაპტაციის მფლობელი ინდივიდების მცირე ჯგუფი ახალ ადაპტურ ზონაში შეღწევით შეიძლება ფართო ადაპტური რადიაციის ან აროგენეზის გზაზე გავიდნენ. პერსპექტიული ზოგადი ადაპტაცია ჩვეულებრივ ორ ან მეტ ორგანოთა სისტემას მოიცავს. სწრაფად ცვალებადი პირობებისადმი შეგუების შედეგად სწრაფადვე ფორმირებული შესაბამისი მორფო-ფიზიოლოგიური ადაპტაციები ფართო ევოლუციურ მნიშვნელობას ვერ იძენენ.
მოწოდებულია ადაპტაციის სხვა სახის კლასიფიკაციაც; ასე მაგალითად, წარმოქმნილი ადაპტური ცვლილების მიხედვით შეიძლება დაჯგუფდეს შემდეგნაირად: 1. სისტემის აგებულების გამარტივება. 2. სისტემის აგებულების გართულება. 3. აგებულებისა და სირთულის შენარჩუნება. პარაზიტებსა (თასმისებრი ჭიები, ზოგიერთი ნემატოდა, ტრემატოდა და მისთ.) და მჯდომარე ცხოველებს (მარჯნები, კანეკლიანები, გარსიანები და სხვ.) ზოგიერთი ორგანოთა სისტემა გამარტივებული ან რედუცირებული აქვთ. მაგალითად, თასმისებრ ჭიებს არ მოეპოვებათ სუნთქვის, სისხლის მიმოქცევისა და საჭმლის მომნელებელ ორგანოთა სისტემა, გრძნობათა ორგანოები, ხოლო ნერვული სისტემა მეტად გამარტივებულია. მაშინ როდესაც აქტიურ ცხოვრებაზე გადასვლას ხშირად თან სდევს თავდასხმისა და თავდაცვის ორგანოების გასრულყოფილება. სოციალური და საზოგადოებრივი უხერხემლო და ხერხემლიანი ცხოველები გაცილებით რთულ ადაპტაციებს იძენენ ვიდრე დანარჩენი.
ფრიად მრავალფეროვანია შეგუებები ონტოგენეტიკური მასშტაბის (ონტოგენეზში გამოვლენის ხანგძლივობის) მიხედვით. ზოგიერთი ადაპტაცია მეტად ხანმოკლე პერიოდით იჩენს თავს, ზოგიც საკმაოდ ხანგრძლივად ნარჩუნდება. ადაპტაციების გარკვეულ ნაწილს მხოლოდ ემბრიონულ განვითარების პერიოდში ფლობს ინდივიდი, ნაწილს კი პოსტემბრიონული განვითარების პერიოდში. ზოგიერთი ადაპტაცია ციკლურია (მაგ., სეზონური დიმორფიზმი - სეზონების მიხედვით შეფერვის ცვლილება), ზოგიც ონტოგენეზის ყველა ეტაპზე მუდმივად და უცვლელად გააჩნია ორგანიზმს (იხ. ცხრ. 10-1).
ორგანიზმების ყოველ ჯგუფში შესაძლებელია ადაპტაციების ზუსტი შესწავლა და მათი კლასიფიკაცია. ასე მაგალითად ძუძუმწოვრებში ადაპტაციები შეიძლება დაჯგუფდეს შემდეგნაირად: ადგილსამყოფელის ტიპების (ხმელეთის ფორმები ანუ ქთონობიონიტები, წყლის ფორმები, ანუ ჰიდრობიონიტები და ა.შ.); კვების ხერხების (ჰერბივორა, ანუ მცენარის მჭამელები; კარნივორა ანუ მტაცებლები; ომნივორა, ანუ ყველაფრის მჭამელები); მოძრაობის ხერხების (მორბენლები, მცურავები, ხტუნიები, მცოცავები და ა.შ.) მიხედვით და სხვა.
ცხრილი 10-1.
ადაპტაციის კლასიფიკაცია
№ |
ადაპტაციის |
ადაპტაციის |
1. |
წარმოშობის |
პრეადაპტაციური, კომბინაციური, პოსტადაპტაციური |
2. |
სხვადასხვა |
გენოტიპური, |
3. |
ევოლუციური |
ზოგადი, |
4. |
გამოწვეული |
სისტემის |
5. |
ონტოგენეზური |
ხანმოკლე, |
6. |
სრულყოფილების |
ზღვარს მიღწეული, |
ნებისმიერი სახეობა არსებობს დროსა და სივრცეში ე.ი. სახეობა გარკვეულ არეალს და ფილოგენეზურ განვითარებას ფლობს. სახეობას წარმოშობის პროცესში უყალიბდება ადაპტაციების მთელი კომპლექსი. სახეობის ევოლუციის პროცესში ადრე წარმოქმნილი ადაპტაციების გასრულყოფილება მიმდინარეობს. ამასთანავე აბიოტური და ბიოტური საარსებო გარემოს ფლუქტუირების გამო იგი ახალ ადაპტაციებსაც იძენს. ნებისმიერ ადაპტაციას სრულყოფილების გარკვეული ზღვრები მოეპოვება. სრულყოფის დონის მიხედვით ადაპტაციები პირობითად სამ ჯგუფად შეიძლება დაიყოს:
1. სრულყოფის რეალურ ზღვარს მიღწეული ადაპტაციები (დნმ-ის სტრუქტურული ორგანიზაცია, გენეტიკური კოდი, დნმ-ის რეპლიკაცია და რეპარაცია; ფრინველებისა და ძუძუმწოვრების ოთხსაკნიანი გულის აგებულება, ძუძუმწოვრებისა და ადამიანის ყურის აგებულება, რთული კამერიანი თვალი ფრინველებსა და ძუძუმწოვრებში და მისთ.).
2. სრულყოფის ზღვარს მიახლოებული ადაპტაციები (მექანიკური ქსოვილის სიმტკიცე, ენერგეტიკული ცვლის პროცესი და მისთ).
3. ადაპტაციები რომელთაც გასრულყოფილების (სრულქმნის) დიდ პოტენციას ფლობენ (ნეირონის ბოჭკოებში იმპულსის გატარების სიჩქარე, პოპულაციაში ინდივიდთა გადარჩენადობის საშუალო მაჩვენებელი და მისთ.).
|
|
12.2.2 10.2.2. ადაპტაციის წარმოქმნის მექანიზმი |
▲ზევით დაბრუნება |
ორგანიზმის, პოპულაციის, სახეობისა და ეკოსისტემის მიერ ევოლუციის პროცესში შეძენილი ყველა თავისებურება ამა თუ იმ სახის ადაპტაციას წარმოადგენს. ფართო მნიშვნელობით ადაპტაცია გულისხმობს საარსებო გარემოსადმი ორგანიზმების (მათ შორის პოპულაციების, სახეობების) სრულ ჰარმონიას; ვიწრო მნიშვნელობით კი სპეციალურ თვისებებს, რითაც კონკრეტულ გარემოში მიიღწევა ორგანიზმის სიცოცხლის შენარჩუნება და წარმატებული გამრავლება. მაშასადამე, ადაპტაცია ორგანიზმების (ინდივიდის, პოპულაციის, სახეობის) და მათი ორგანოების აგებულებისა და ფუნქციის გარემო პირობებისადმი შეგუების პროცესს (ადაპტაციოგენეზი) გამოხატავს, იმავდროულად შედეგსაც ე.ი. კონკრეტულ ნიშან-თვისებათა თავისებურებებს, რომელიც ორგანიზმების მაღალ სიცოცხლისუნარიანობას, ფერტილობასა და კონკურენტუნარიანობას უზრუნველყოფს.
ევოლუციური თვალთახედვით ცნება „ადაპტაცია“ ვრცელდება როგორც ინდივიდებზე, ისე პოპულაციებსა და სახეობებზე. სხვადასხვა საარსებო გარემოში ორგანიზმის ნიშან-თვისების გამოვლენის ხარისხი გარკვეულ (რეაქციის ნორმის) ფარგლებში იცვლება. ასე მაგალითად, ფოთლის ფირფიტის ფორმის ცვლილება ამფიბიოტურ მცენარეებში (ისარა, წყლის ბაია და მისთ.); სეზონური დიმორფიზმი ცხოველებში და სხვ. ეს უნარი ორგანიზმებმა ისტორიული განვითარების პროცესში შეიძინეს და მათ მემკვიდრული აპარატი (გენოტიპი) აკონტროლებს. შეცვლილ საარსებო პირობებს ორგანიზმები აკლიმაციით (მაგ., ტემპერატურის ხანგრძლივი თანდათანობითი ცვლილების დროს ძუძუმწოვრებში ბეწვის, ფრინველებში ბუმბულის შეცვლა...) და მოდიფიკაციით (ერთნაირი გენოტიპის ინდივიდებზე გარემოს მოქმედებით გამოწვეული ნაირგვარობა; იხ. §5) რეაგირებს.
ადაპტაციის წარმოშობის აუცილებელი წინა პირობაა ელემენტარული ევოლუციური მასალის - მემკვიდრული ცვალებადობის არსებობა. იგი უნდა დაექვემდებაროს ევოლუციის ელემენტარული ფაქტორების, უპირველეს ყოვლისა, ბუნებრივი გადარჩევის მოქმედებას. პოპულაციაში სასარგებლო გენების შემცველი გენოტიპების ფორმირება ადაპტაციას ჯერ კიდევ არ წარმოადგენს. სელექციური ღირებულების მქონე გენოტიპების ჩამოყალიბება არის ელემენტარული ადაპტაციური მოვლენა. ელემენტარული ადაპტაციური მოვლენა არ ნიშნავს ადაპტაციის ჩამოყალიბებას. ადაპტაციის წარმოშობა ხდება მას შემდეგ, როდესაც პოპულაციაში გარემოს ელემენტების მიმართ სპეციალიზებული ნიშანი წარმოიქმნება. თუ ჩამოყალიბებული ნიშნით პოპულაციაში ინდივიდთა ელიმინაცია ქვეითდება, ასეთ ნიშანს სელექციური ღირებულება ენიჭება. როდესაც ნიშანი მამოძრავებელი გადარჩევის მოქმედებით მიესადაგება საარსებო გარემოს ელემენტებს, მასზე მოქმედებას იწყებს მასტაბილიზებელი გადარჩევა და პოპულაციაში მისი შენარჩუნება ხდება. როდესაც ნიშანი სელექციურ მნიშვნელობას კარგავს, ე.ი. არ ექვემდებარება მამოძრავებელი, მასტაბილიზებელი ან დიზურპტული გადარჩევის მოქმედებას, იგი რედუცირდება და ქრება. ამრიგად, ადაპტაციის ჩამოყალიბება და მისი სრულყოფა ბუნებრივი გადარჩევის მოქმედების შედეგად ხორციელდება. ადაპტაცია გარკვეულად განიცდის ევოლუციურ ცვლილებებს შესაბამისი ორგანიზმების გარემოსთან მიმართების შეცვლის გამო ახალი ადაპტაციების გაჩენასთან დაკავშირებით.
|
|
12.3 10.3. ადაპტაციის ფარდობითი ხასიათი |
▲ზევით დაბრუნება |
ნებისმიერი ადაპტაცია კონკრეტული საარსებო პირობებისათვის ყალიბდება. იგი უზრუნველყოფს ინდივიდის გადარჩენას და შთამომავლობის დატოვებას მხოლოდ იმ კონკრეტულ პირობებში, რომელშიაც ევოლუციის ელემენტარული ფაქტორების ზემოქმედებით ყალიბდება. ამდენად ნებისმიერი ადაპტაცია ფარდობით ხასიათს იძენს. გარკვეული კონკრეტული პირობებისადმი ჩამოყალიბებული ადაპტაცია, სხვა პირობებში არა მარტო უსარგებლოა, არამედ საზიანოც შეიძლება აღმოჩნდეს. ასე მაგალითად, ნექტონურ სწრაფად მცურავი თევზების თითისტარისებრი ფორმა და ლაყუჩებით სუნთქვა სრულყოფილად აგუებს ცხოველს წყლიან გარემოს, იგივე სამარჯვეები სრულიად გამოუსადეგარია ხმელეთზე არსებობისათვის. ნამგალასა და მერცხალს გააჩნიათ გრძელი ფრთები და კარგად არიან ადაპტირებული ჰაერში ფრენასა და სარჩოს მოპოვებასთან. მაგრამ მიწაზე ისინი სავსებით უსუსურნი არიან. სეზონური დიმორფიზმის მფლობელი ცხოველი (თეთრი კურდღელი, თეთრი გნოლი, ყარსაღი და სხვ.) თოვლზე შეუმჩნეველია, თუ თოვლი ადრე არ მოვიდა, მაშინ თეთრი შეფერილობა მათ ააშკარავებს.
ადაპტაცია იმ კონკრეტული საარსებო პირობებისთვისაც კი, რომელშიაც ის არის ჩამოყალიბებული შეფარდებითი ხასიათისაა. ასე მაგალითად, კუს ჯავშანი ეფექტურად იცავს ზოგიერთი თავდამსხმელისაგან, მაგრამ არაეფექტურია მტაცებელი ფრინველების წინააღმდეგ. მტაცებელი ფრინველი ბრჭყალებით იტაცებს ცხოველს, ააქვს ჰაერში და ჩამოაგდებს. ბაკანი სკდება, სხეული შიშვლდება და ფრინველისთვის საჭმელად ხელმისაწვდომი ხდება. მრავალი ფრინველი მფარველობითი შეფერილობით საიმედოდ ეხმარება ფონს, მაგრამ სუნი ააშკარავებს და მტაცებლის მსხვერპლი ხდება. ჭინჭარს მსუსხავი ბუსუსები საიმედოდ იცავს ზოგიერთი ცხოველისაგან (ლოკოკინა, ლოქორა და მისთ), მაგრამ საკვებად იყენებს პეპელა მეჭინჭრიას მუხლუხი. მრავალი ცხოველი არ ეტანება ეკლიან მცენარეს, მაგრამ ზოგიერთი ცხოველი მათ საკვებად იყენებს (მაგალითად, აქლემი იკვებება ცერცვეკალათი). ფარდობითია თავდაცვითი შეფერილობა, მიმიკრიის მოვლენა. მრავალი ფრინველი არ ეტანება მოხატულობით ფუტკრისა და კრაზანას მსგავს მწერებს, მაგრამ ზოგი ფრინველი (მაგ., კვირიონი) კენკავს ფუტკარსა და კრაზანას, ასევე მის მსგავს მწერებსაც.
ადაპტაციოგენეზში ზოგჯერ ორგანიზმებს რეციპროკული ადაპტაციები უყალიბდებათ. ზოგიერთ ფორმაში თავდაცვითი თავისებურებების გამომუშავება, პოტენციურ მომხმარებლებში საპირისპირო სამარჯვეების ჩამოყალიბებას იწვევს, რითაც თავდაცვითი შეგუების ეფექტი ნეიტრალდება ან უკიდურესად ქვეითდება. ზოგიერთი რძიანა მცენარე შეიცავს ტოქსინს (გულის გლუკოზიდი, სხვა ალკალიდი), რის გამოც მრავალი ცხოველი არ ეტანება, მაგრამ ზოგიერთი პეპლის მუხლუხის (დანაიდა, სფინქსი...) ძირითადი საკვებია. შხამიანი გველით იკვებება ზოგიერთი ცხოველი (მანგუსტი, გველიჭამია არწივი, ფრინველმდივანი და მისთ.). უნდა აღინიშნოს, რომ ზოგიერთი გველისმჭამელი ცხოველი შხამისადმი იმუნურია.
შეგუების ფარდობითობა ვლინდება არა მარტო სივრცეში, არამედ დროშიც. ამ უკანასკნელის დასტურია დედამიწაზე ბიოსფეროს განვითარების პერიოდში მრავალი მცენარისა და ცხოველის ამოწყდომა. დარვინამდელ მეცნიერებს მიაჩნდათ, რომ შეგუება აბსოლიტურ ხასიათს ფლობდა. ჩ.დარვინმა დაასაბუთა შეგუების ფარდობითობა. შეგუების ფარდობითი ხასიათის გამო მუდამ არსებობს მიკროევოლუციის მამოძრავებელი ფაქტორებისათვის მოქმედების განუსაზღვრელი ასპარეზი, იმ შემთხვევაშიც კი როდესაც საარსებო პირობები უცვლელი რჩება. ბუნებრივი გადარჩევის მოქმედებით ხორციელდება ადაპტაციის სრულქმნა.
სახეობის არსებობა შეზღუდულია სივრცესა და დროში. ადაპტაცია ფარდობითია, რადგან შეზღუდულია შესაბამისი სახეობის არსებობის დროთი. სახეობის ჩამოყალიბება იმავე დროს მათი დამახასიათებელი ადაპტაციების მთელი კომპლექსის ჩამოყალიბების პროცესია. ადაპტაცია ჩნდება სახეობის არსებობისგანსაზღვრულ საფეხურზე და შემდგომ ევოლუირებს და მისი გასრულყოფილება ხდება. ადაპტაციას გარკვეული საზღვრები გააჩნია. ორგანულ სამყაროში ვერცერთი ადაპტაცია აბსოლუტურ სრულყოფილებას ვერ აღწევს. ამრიგად იგი ყოველთვის ფარდობით ხასიათს ატარებს.
|
|
13 § 11. სახეობა-ევოლუციის პროცესის ძირითადი ეტაპი |
▲ზევით დაბრუნება |
|
|
13.1 11.1. სახეობის კონცეფციები |
▲ზევით დაბრუნება |
სახეობის პრობლემა - ბიოლოგიის ერთ-ერთ ცენტრალურ და ურთულეს საკითხს წარმოადგენს. ცოცხალი მატერია მაღალორგანიზებულობით გამოირჩევა. მისი ორგანიზაციის ზოგიერთი ერთეული - უჯრედი, ინდივიდი, პოპულაცია გადარჩევის შესაძლო დამოუკიდებელი ერთეულია, თითოეულს საამისო თვისება გააჩნია, მაგრამ ცოცხალი სისტემის უმნიშვნელოვანეს იერარქიულ დონესა და ტაქსონომიურ ერთეულს სახეობა წარმოადგენს. უკვე კარგა ხანია, რაც გამოყოფილია ორგანიზმთა ჯგუფები, რომელთაც სახეობას უწოდებენ, მაგრამ თუ რა არის სახეობა ბოლომდე არ არის გარკვეული.
სადღეისოდ არსებობს სახეობის სამი მთავარი კონცეფცია. პირველი, ფილოსოფიურ და ლოგიკურ კატეგორიას განეკუთვნება. იგი არისტოტელედან მომდინარეობს. სახეობა არის აზროვნებითი კატეგორია, უფრო ზუსტად გარკვეული კლასი (ლოგიკური აზრით), რომელთაც ნებისმიერი სულიერი და უსულო საგანი განეკუთვნება. მას უნდა ჰქონდეს გარკვეული ზოგადი თვისება.
მეორე არის ტიპოლოგიური (ე.წ. მორფოლოგიური) კონცეფცია, იგი ლოგიკურ კონცეფციას ეფუძნება. ე.ი. ლოგიკური განმარტების დამატებაა, რომელიც მხოლოდ ცოცხალ სისტემაზე ვრცელდება. ტიპოლოგიური განმარტებით სახეობა არის „სახეობრივი ნიშნის მიხედვით ერთნაირ ინდივიდთა ერთობლიობა“. სახეობას გააჩნია ინვარიანტული (მონოტიპური) ნიშნები, რომლებიც სადიაგნოსტიკო მნიშვნელობას იძენენ სახეობის იდენტიფიცირებისას. ინვარიანტული მორფოლოგიური, ფიზიოლოგიური, ეთოლოგიური და ბიოქიმიური ნიშნებით ადვილად შეიძლება კონკრეტული სახეობის გამიჯვნა დანარჩენებისაგან. ახალი სახეობის წარმოქმნა სალტაციების (მყისიერად გენომის მსხვილი რეორგანიზაცია) მეშვეობით მიმდინარეობს. ტიპოლოგეიური კონცეფციის მომხრენი უარყოფენ ბუნებრივი გადარჩევისა და სხვა მიკროევულუციის მამოძრავებელი ძალების როლს სახეობათა წარმოქმნის პროცესში. ამგვარი შეხედულებები ძირითადად ფეხმოკიდებული იყო XVIII-XIXსს. მკვლევრებში. ამ იდეას ზოგიერთი მეცნიერი ამჟამადაც იზიარებს.
მესამე კონცეფცია ე.წ. პოპულაციონისტური ანუ ბიოლოგიური, სათავეს ჩ. დარვინის მოძღვრებიდან იღებს, ამ შეხედულების თანახმად, სახეობა შედგება პოპულაციებისაგან, რომელთა წევრებს თავისუფალი შეჯვარების (პანმიქსის) უნარი აქვთ. სახეობა განისაზღვრება როგორც „რეალურად ან პოტენციურად შეჯვარებადი ჯგუფები, რომლებიც რეპროდუქციულად იზოლირებული არიან ასეთივე ჯგუფებისაგან“. სახეობისათვის დამახასიათებელი ნიშნები პოლიტიპურია, ამდენად სისტემატიკისათვის არ არის ხელსაყრელი. სახეობათა წარმოქმნის პროცესი გრადუალურ (თანდათანობით) ხასიათს ატარებს. არ გამორიცხავენ მის მყისიერად (კვანტურ) წარმოქმნასაც. სახეობათა წარმოქმნას ევოლუციის ელემენტარული ფაქტორების მოქმედება აპირობებს, მათგან მთავარი და წარმმართველი არის ბუნებრივი გადარჩევა.
ტიპოლოგიური კონცეფციის მიხედვით მთავარი და არსებითია მსგავსება და ერთგვაროვნება, პოპულაციონისტურით - შეჯვარების უნარი. ამ ორი კონცეფციის მიხედვით იდენტიფიცირებული სახეობა ერთიმეორეს მეტ-ნაკლებად ემთხვევა. კერძოდ, ერთმანეთის მსგავსი ინდივიდები ხშირად ეჯვარებიან ერთიმეორეს, ხოლო ინდივიდები რომლებიც ერთიმეორეს ეჯვარება ხშირად მორფოლოგიურად მსგავსია. მორფოლოგიურად მსგავს ინდივიდებს მრავალი ერთნაირი გენი მოეპოვებათ, რაც მათ ერთგვაროვნებას და შეჯვარების უნარს განაპირობებს. ორივე კონცეფცია, მხოლოდ ერთ პუნქტში თანხვდება ერთმანეთს, კერძოდ, აღიარებენ რეპროდუქციულ იზოლაციის წამყვან როლს სახეობათა ფორმირების პროცესში. ამ კონცეფციათა მსგავსება მოჩვენებითი და ზედაპირულია.
ზოგჯერ ერთი და იმავე სახეობის ინდივიდები ერთიმეორისგან მორფოლოგიურად განსხვავდებიან. სანიმუშო მაგალითია სქესობრივი და სეზონური დიმორფიზმი. კ. ლინემ მორფოლოგიური კრიტერიუმიდან გამომდინარე დედალი და მამალი გარეული იხვი განსხვავებულ სახეობებად აღწერა. ნიადაგის მჟავიანობის მიხედვით ჰორტენზიას (Hydrangia hortensia) ყვავილი ლურჯი ან მოწითალოა (იხ. ფენოტიპური ცვალებადობა §5.1), ლოკოკინას (Cepaea nemoralis) ნიჟარები შეფერილობითა და ზოლიანობით განსხვავდებიან (იხ. § 8.8 და 9.1).
სახეობა წარმოადგენს ყველაზე დიდ პოპულაციურ ერთეულს. ერთი და იმავე პოპულაციის წევრები თავისუფლად ეჯვარებიან ერთმანეთს. ზოგჯერ ცხოველი შორ მანძილზე მიგრირებს ან მცენარის თესლი ქარს, ცხოველს ან წყალს შორს გადააქვს. ყველა ამ შემთხვევაში ხდება ერთი პოპულაციის წევრების მეორეში მიღწევა და სხვადასხვა პოპულაციათა შორის გენთა მიმოცვლა. გენთა ნაკადის მეშვეობით გენოფონდი ერთიანი ხდება. მისი მეშვეობით მიიღწევა სახეობის, როგორც ევოლუციის ეტაპის მდგრადობა (იხ. §7.3).
სახეობის კონცეფციების განვითარების ისტორია. სახეობის სპეციფიკისა და კრიტერიუმების შესახებ შეხედულებები ბიოლოგიის სხვადასხვა დარგში მოპოვებული ახალი ფაქტების დაგროვების კვალობაზე ვითარდებოდა. სახეობის კონცეფციის განსაზღვრა დამოკიდებულია მკვლევრის მეთოდოლოგიურ მიდგომაზე, რომლის მომარჯვებითაც მოპოვებული ფაქტობრივი მასალის ინტერპრეტირებას ახდენს.
ტერმინი „სახეობა“ (Species) პირველად თავის შრომებში არისტოტელემ გამოიყენა. იგი ამ ტერმინით მსგავს ცხოველებს აღნიშნავდა. სახეობის ცნებას არისტოტელესთან მხოლოდ ფილოსოფიური და ლოგიკური დატვირთვა აქვს. ამასთანავე, ის სახეობას სიცოცხლის ორგანიზაციის სპეციფიკურ დონედ კი არ განიხილავს, არამედ როგორც ზოგადი კატეგორიის (გვარის) შემადგენელ კერძო კატეგორიას. ორივე მათგანი, გაცილებით დიდ, იერარქიულად ზემდგომ ერთობლიობაშია გაერთიანებული, როგორც მთელის შემადგენელი კერძო ნაწილი.
XVIს. ბოლომდე ტერმინი „სახეობა“ ბიოლოგიაში არისტოტელესეული განსაზღვრებით იხმარებოდა. პირველად სახეობის ცნებას ბიოლოგიური შინაარსი ინგლისელმა ბოტანიკოსმა ჯ. რეიმ (1627-1705) მისცა. იგი სახეობას ტიპოლოგიური პოზიციიდან განსაზღვრავდა. მანვე განამტკიცა ბიოლოგიაში სახეობის აღნიშვნის ბინარული პრინციპი, რომელიც პირველად შემოიღო შვედმა ბოტანიკოსმა კასპარ ბაუგინმა (1623). სახეობები რეიმ გააერთიანა უფრო დიდ ტაქსონებში. რეის შეხედულებით, სახეობა ელემენტარული (დაუყოფადი) ერთეულია, რომელიც მორფოლოგიურად ერთგვაროვანი ინდივიდებისგან შედგება. სახეობის შემადგენელი ინდივიდები ურთიერთშეჯვარებით მრავლდებიან და ნიშან-თვისებებს შთამომავლობას გადასცემენ.
თანამედროვე მეცნიერული სისტემატიკის ფუძემდებელმა, გამოჩენილმა ნატურალისტმა და ექიმმა კ. ლინემ (1707-1778) ნაშრომში „ბუნების სისტემა“ (1735), პირველმა ჩამოაყალიბა შეხედულება სახეობის, როგორც უნივერსალურ, დისკრეტულ და ბუნებაში რეალურად არსებულ კატეგორიაზე. მისი შეხედულებით, სახეობა მდგრადი და უცვლელია, რომელიც მკვეთრადაა გამიჯნული დანარჩენი სახეობებისგან. სახეობის აღნიშვნის აუცილებელ პირობად დაამკვიდრა საერთაშორისო (ლათინური) ბინარული ნომენკლატურა. ლინეს თეორიული შეხედულებები საფუძვლად დაედო სახეობის ტიპოლოგიურ კონცეფციას. ამ მიდგომას პრაქტიკულ სისტემატიკაში ამჟამადაც იყენებენ.
ბუნებაში სახეობის მუდმივობისა და მდგრადობის იდეის აღიარებამ ხელი შეუწყო ბიოლოგიაში ტიპოლოგიური კონცეფციის გაბატონებას. ამ იდეას დაუპირისპირდა ტრანსფორმიზმი - მოძღვრება სახეობათა ცვლილებების შესახებ (ჟ. ბიუფონი, ჟ. სენტ-ილერი და სხვ.) ჩამოყალიბდა სახეობის ცნობილი კრიტერიუმი: ინდივიდები რომლებიც ერთმანეთს ეჯვარებიან და ნაყოფიერ შთამომავლობას წარმოქმნიან ერთსა და იმავე სახეობას მიეკუთვნებიან (ბიუფონი).
პირველი ერთიანი ევოლუციური თეორიის ავტორი ჟან ბატისტ ლამარკი ნაშრომში „ზოოლოგიის ფილოსოფია“ (1809), რომელშიც მისი ევოლუციური იდეებია ჩამოყალიბებული, უარყოფდა სახეობის დისკრეტულობას. ბუნებრივია, აქედან გამომდინარე იგი უარყოფდა სახეობის რეალურად არსებობას ბუნებაში. მას მიაჩნდა, რომ სახეობა და სხვა სისტემატიკური ერთეულები ხელოვნურად გამოგონილი კატეგორიებია, რათა ადვილად მოახდინონ ბუნებაში არსებული ფორმების დაჯგუფება. რადგანაც ლამარკი სახეობას განიხილავდა როგორც ნომინალურ კატეგორიას (ე.ი. ბუნებაში რეალურად არ არსებობს). მოგვიანებით მეცნიერებმა ამ პრინციპს ნომინალისტური უწოდეს. ლამარკის შეხედულებებმა სახეობის შესახებ თანამედროვეთა შორის აღიარება ვერ პოვა.
სახეობის პოპულაციონისტურ ანუ ბიოლოგიურ კონცეფციას საფუძველი ჩ. დარვინის ფუნდამენტურმა ნაშრომმა „სახეობათა წარმოშობამ“ (1859) ჩაუყარა. დარვინის შეხედულებით, სახეობა არის ბუნებაში რეალურად არსებული კატეგორია, რომელიც არამდგრადი და დინამიურია. დროში (ხანგრძლივი პერიოდის მანძილზე) დივერგირების შედეგად მისგან ახალი სახეობები წარმოიქმნება. სახეობა ფლობს რთულ სტრუქტურას და სახესხვაობისგან (ქვესახეობები) შედგება. დარვინის შეხედულებით, სახესხვაობა წარმოადგენს ფორმირებად სახეობას.
XX საუკუნის დასაწყისში სახეობის ტიპოლოგიურ კონცეფციას საფუძველი საკმაოდ შეერყა. მცენარეებსა და ცხოველებში კლასიფიკაციის ძირითად ერთეულად გეოგრაფიული რასა წამოაყენეს. იგი მიაჩნდათ დისკრეტულ (დაუყოფად) სტრუქტურად. სახეობა განიხილებოდა როგორც გეოგრაფიულ რასათა ან ქვესახეობათა ერთობლიობა. მოგვიანებით გამოირკვა, რომ ქვესახეობას რთული სტრუქტურა გააჩნია. მკვლევარებმა მასში გამოავლინეს სეზონური, ეკოლოგიური და ფიზიოლოგიური რასები. სახეობის შესახებ შეხედულებაზე არსებითი გარდატეხა მოხდა მას შემდეგ, რაც გენეტიკისა და ევოლუციური თეორიის სინთეზი განხორციელდა. პოპულაციებში ჩატარებული გენეტიკური ანალიზით დაადგინეს, რომ სახეობას რთული გენეტიკური სტრუქტურა მოეპოვება. ზოგიერთი მკვლევარი ბუნებაში სახეობის რეალურად არსებობის უარყოფამდეც კი მივიდა. მათ ტრადიციული „ლინესეული“ სახეობა ე.წ. ლინეონი (ლინეს გვარის მიხედვით) დაჰყვეს მრავალ მცირე, მაგრამ მდგრად სახეობად ე.წ. ჟორდანონებად (ფრანგი ბოტანიკოსის ჟორდანის მიხედვით). რადგანაც სახეობათა იდენტიფიცირების ეს პრინციპი ბუნებაში არსებულ რეალურ სურათს არ ასახავდა, ამდაგვარი ნოვაცია სისტემატიკაში ვერ დაინერგა. მას მხოლოდ ისტორიული მნიშვნელობა შემორჩა.
სახეობის თანამედროვე პოპულაციონისტური ანუ ბიოლოგიური კონცეფციის თანახმად, იგი არის რეალურად არსებული რთული გენეტიკური სისტემა. ერთ სახეობაში გაერთიანებული ინდივიდების გენთა ერთობლიობა ქმნის გენოფონდს. სახეობა სხვა სახეობის გენების შეღწევისაგან საიზოლაციო მექანიზმებით არის დაცული. სახეობა ზოგიერთ შემთხვევაში შეიცავს შიგასახეობრივ ფორმებს (პოპულაციათა ჯგუფი, ქვესახეობა და მისთ.). ამ ფორმების წარმომადგენლები შეიძლება დრო და დრო ერთიმეორეს შეეჯვარონ და ფერტილური შთამომავლობა წარმოქმნან. სახეობის თანამედროვე კონცეფციის ჩამოყალიბებაზე გადამწყვეტი როლი გენეტიკის უახლესმა მიღწევებმა მოახდინა. სახეობის თეორიის დამუშავებაში დიდი როლი შეასრულა ჯ. ჰაქსლის, ე. მაირის, ჯ. სიმპსონის, ფ. დობჟანსკის, ვ. გრანტის და სხვათა გამოკვლევებმა.
|
|
13.2 11.2. სახეობის ცნების ფორმულირება |
▲ზევით დაბრუნება |
ხშირად ორგანული ბუნების ჩვეულებრივი და ფართოდ გავრცელებული მოვლენების ფორმულირება რთულია. ეს სიტუაცია მაშინ ხდება, თუ მოვლენას წაშლილი ან ლაბილური საზღვრები აქვს, აგრეთვე, როდესაც მოვლენა რთული შინაგანად წინააღმდეგობრივი კომპონენტებისგან შედგება. ასეთ კატეგორიათა რიცხვს მიეკუთვნება სახეობის ცნება. ორგანულ ბუნებაში სახეობები მეტად მრავალფეროვანია, ჩვენი ცოდნა კი - მწირი. თანამედროვე ცხოველთა სამყაროში 2 მლნ. სახეობაა აღწერილი, ხოლო მცენარეთა სამეფოში 0,5 მლნ. სახეობა. ყოველწლიურად ათეულობით ახალი სახეობის გამოვლენა ხდება. მიაჩნიათ, რომ ამჟამად დედამიწაზე 5 მილიონზე მეტი სახეობა სახლობს (გრანტი, 1991). სისტემატიკურ ტაქსონებში (გვარი, ოჯახი, რიგი, კლასი და მისთ.). სახეობები არათანაბრადაა განაწილებული. მაგალითად, მწერების კლასი 750 ათასზე მეტ სახეობას შეიცავს, ხოლო რეპტილიების ნისკარტთავიანისებრთა რიგში ამჟამად მცხოვრები ერთადერთი სახეობა ტუატარა (hateria, Sphenodon punctatus) არის გაერთიანებული; ასევე შიშველთესლოვანების გინკოალების რიგში ამჟამად ერთი ოჯახია თავის ერთი სახეობით - გინკგოთი (Ginkgo biloba) წარმოდგენილი.
სადღეისოდ სახეობის ცნების საყოველთაოდ აღიარებული განმარტება ჯერ კიდევ არაა შემუშავებული. გვაქვს მხოლოდ სახეობის ფორმულირება სახეობა წარმოადგენს ევოლუციის პროცესის თვისებრივ ეტაპს.
სახეობა არის ინდივიდთა ერთობლიობა, რომელთაც აქვთ საერთო მორფოფიზიოლოგიური ნიშნები და ერთმანეთთან შეჯვარების უნარი, ქმნიან პოპულაციების სისტემას, რომლებიც საერთო (მთლიან ან წყვეტილ) არეალს იკავებენ; სახეობები ბუნებაში გამიჯნულია ერთმანეთისგან და მდგრად გენეტიკურ სისტემას ქმნიან.
ნებისმიერი განმარტება არასრულია და არ ამოწურავს ცნებას. ბუნებრივია წარმოდგენილი სახეობის განმარტებაც ნაკლულია. ამდენად საჭიროდ მიგვაჩნია ზოგიერთი მნიშვნელოვანი დამატების შეტანა. ნებისმიერი სახეობა წარმოადგენს პოპულაციათა რთულ სისტემას, რომელთა ფორმირება კონკრეტული ეკოსისტემის ეკოლოგიურ ნიშებში მიმდინარეობს. პოპულაციათა ამ სისტემას საერთო ევოლუციური ბედი გააჩნია. ჩვეულებრივ, თითქმის ყველა სახეობა პოლიტიპურია ე.ი. შეიცავს მორფოლოგიურად განსხვავებულ (ქვესახეობა, ნახევარსახეობა და მისთ.) ჯგუფებს.
სადღეისოდ სახეობათა უმნიშვნელო ნაწილია სრულყოფილად შესწავლილი. მათი გარკვევისას შესაძლებელია ზემოთ მოყვანილი ფორმულირების ყველა პუნქტს გაეცეს პასუხი. ნეონტოლოგიური (ამჟამად მცხოვრები) სახეობების შესწავლით გაირკვა, რომ ბუნებაში ისინი განსხვავდებიან მორფოლოგიურად ან ფიზიოლოგიურად; ეკოლოგიურად ან ეთოლოგიურად, ასევე გეოგრაფიული გავრცელებით. ერთი და იმავე სახეობის პოპულაციის წევრები თავისუფლად ეჯვარებიან ერთიმეორეს (პანმიქსია), განსხვავებული სახეობები კი ერთმანეთისგან რეპროდუქციულადაა იზოლირებული. მაშასადამე სახეობის იდენტიფიცირებისას არსებითი მნიშვნელობა მისი შემადგენელი ინდივიდების ერთმანეთთან შეჯვარებისა და ნაყოფიერი შთამომავლობის წარმოქმნის უნარს ენიჭება.
პრინციპული ხასიათის სიძნელე იქმნება აგამურ (უსქესო), ობლიგატურ პართენოგენეზურ და მკაცრად თვითგამანაყოფიერებელ ფორმებში. მაგრამ სახეობები ამდაგვარ ფორმებსაც მოეპოვებათ. ამჟამად სახეობას განმარტავენ, არა როგორც დახშულ გენეტიკურ სისტემას, არამედ მდგრად გენეტიკურ სისტემას. ამდაგვარი ფორმირებისას წარმოდგენილი სახეობის კონცეფცია ვრცელდება ნებისმიერ სახეობაზე; მათ შორის აგამურ, ობლიგატური პართენოგენეზულ და თვითგამანაყოფიერებელ სისტემებზეც. ასეთ ფორმებში სახეობად მიჩნეულია ახლო მდგომ ბიოტიპთა სისტემები - ფენოტიპურად ახლომდგომი ინდივიდები, რომელთაც გააჩნიათ ახლონათესაური გენოტიპები, საერთო არეალს იკავებენ და საერთო ევოლუციური ბედი აქვთ. მათი ერთიანობა გენოტიპთა ერთგვაროვნობითაა განპირობებული. ხატოვნად თუ ვიტყვით, აგამური ფორმები მრავალი მცირე ნაწილებისგან შედგენილ ერთ დიდ ორგანიზმად შეიძლება წარმოვიდგინოთ. ამასთანავე აგამურობა, პართენოგენეზულობა და თვითგამანაყოფიერებლობა მეორეული მოვლენაა, რომელიც ევოლუციის პროცესში ამ მიმართულებით სპეციალიზებულ ჯგუფებს ჩამოუყალიბდათ. მოგვიანებით მათ შეიძინეს რეკომბინაციის სხვადასხვა მექანიზმები. ასე მაგალითად, მიკროორგანიზმებს საამისოდ გააჩნიათ ტრანსფორმაცია, ტრანსდუქცია და კონუგაცია. ტრანსდუქციით ვირუსების მეშვეობით ხორციელდება გენეტიკური მასალის ლატერალური ანუ ჰორიზონტული გადატანა ერთი ბაქტერიული უჯრედიდან მეორეში. ბოლო პერიოდში ანალოგიური მოქმედების მექანიზმები გამოვლენილია ეუკარიოტულ ორგანიზმებშიც (იხ. §5.3. და 12.10).
პრინციპული სიძნელე იქმნება „სახეობის“ ცნების პალეონტოლოგიაში გამოყენების შემთხვევაშიც. პალეონტოლოგი იკვლევს სხვადასხვა განამარხებულ ფორმას, განფენილს, როგორც სივრცეში (მსგავსად ნეონტოლოგიისა) ისე დროში. შეუძლებელია თეზის - „ურთიერთ შეჯვარების უნარის“ გამოყენება, რითაც ახლონათესაურ ურთიერთდამოკიდებულებას ადგენენ. პალეონტოლოგიაში პრაქტიკულად გამოიყენება მორფოლოგიური და რეკონსტრუქციულ-ბიოლოგიური (ცხოველმყოფელობის პროდუქტები, ნაფეხურები, კაპროლიტები და მისთ.) კრიტერიუმი. სახეობრივი რანგით ნამარხი ფორმების გამიჯვნა იშვიათ შემთხვევაში შეიძლება. კერძოდ, როდესაც ერთსა და იმავე ადგილას განსხვავებულ ჰორიზონტულ სიბრტყეში მრავალი მორფოლოგიურად განსხვავებული ფორმაა განამარხებული. ამდენად სახეობის ზემოთ მოყვანილი განსაზღვრების გამოყენება პალეონტოლოგიაში შეუძლებელია. პალეონტოლოგიაში ფილოგენეზური ხის მონაკვეთს, რომელიც პალეონტოლოგიური სახეობის ექვივალენტურია აღნიშნავენ ტერმინით ფრატრია (ფილუმი).
სახეობის ზოგადი ნიშნები: დისკრეტულობა. სახეობა სიცოცხლის ერთ-ერთი მნიშვნელოვანი დონეა. იგი სიცოცხლის კომპლექსური და რთული საფეხურია. ცალკეული სახეობა სივრცეში განცალკევებულია (დისკრეტულია) სხვა ანალოგიური ფორმირებისგან. იგი ფლობს სპეციფიკურ ორგანიზაციას და თაობათა უწყვეტ რიგში ამ სპეციფიკურობის შენარჩუნების უნარს. რიცხოვნობა. სახეობა შედგება მრავალი ინდივიდისაგან. გააჩნია რიცხოვნობის საშუალო დონე, რომელიც მნიშვნელოვან მახასიათებელს წარმოადგენს. ერთიანობა. სახეობა რთულად ორგანიზებული სისტემაა. მოეპოვება რთული შინაგანი სტრუქტურა, რომელიც გენთა ნაკადით ერთიმეორესთანაა დაკავშირებული და იქმნება უნიკალური ერთიანი გენეტიკური სტრუქტურა - გენოფონდი. მდგრადობა. სახეობის შემადგენელი ინდივიდების არსებობა დროში განსაზღვრულია. სახეობაში მიმდინარეობს თაობათა მონაცვლეობის პროცესი, რის გამოც იგი ხანგრძლივი დროის განმავლობაში არსებობს (ე.ი. დროსა და სივრცეში). სახეობის არსებობა დროში არ არის დაპროგრამებული. ისტორიულობა. მიუხედავად იმისა, რომ სახეობის არსებობის ვადა დროში არ არის განსაზღვრული მისი მდგრადობა და ერთიანობა ფარდობითია. თაობათა უწყვეტ რიგში (ე.ი. დროში) მიმდინარეობს თვისებრივი ცვლილებები ე.ი. ევოლუირებს. ევოლუციის ფაქტორების პირუკუ მოქმედება გარკვეულ გარემო პირობებში მის ამოწყვეტას იწვევს. როგორც ახალ სახეობათა წარმოქმნა, ისე ამოწყდომაც წარმოადგენს ბუნებრივ პროცესს. ყოველ სახეობას მისთვის დამახასიათებელი ფილოგენია აქვს.
|
|
13.3 11.3. სახეობის კრიტერიუმები |
▲ზევით დაბრუნება |
სახეობები ერთმანეთისაგან მრავალი ნიშან-თვისებით განსხვავდებიან. ერთ გვარში გაერთიანებული მონათესავე სახეობები საერთო მსგავსების მიუხედავად განსხვავდებიან მორფოლოგიურად, გეოგრაფიული გავრცელებით, ასევე ქცევებით, ფიზიოლოგიური ნიშნებით და სხვ. სახეობისათვის დამახასიათებელ ნიშან-თვისებებს კრიტერიუმებს უწოდებენ. კრიტერიუმებს ბუნებიდან სახეობის გამოსაყოფად იყენებენ. განასხვავებენ სახეობის რამდენიმე კრიტერიუმს.
მორფოლოგიური კრიტერიუმი. მეცნიერებაში გამოყენებულ კრიტერიუმთა შორის ერთ-ერთი უძველესია. იგი ეფუძნება ერთ სახეობაში გაერთიანებული ინდივიდების მორფოლოგიურ მსგავსებას. იგი ერთადერთი კრიტერიუმია, რომელიც პალეონტოლოგიაში გამოიყენება. ამ მეთოდით მსჯელობენ გარდაქმნის დონეზე. ერთიმეორეს ადარებენ თანამედროვე ფორმებს, ასევე თანამედროვე და წინაპარ ფორმებს. ერთ გვარში გაერთიანებული (მონათესავე) სახეობის ერთი ნაწილი მორფოლოგიურად მკვეთრად განსხვავდება, მეორე ნაწილი - უმნიშვნელოდ. უკანასკნელ შემთხვევაში ახლომონათესავე სახეობები მორფოლოგიური ნიშნებით ერთიმეორეს ფარავენ, რის გამოც გაძნელებულია ბუნებიდან სახეობის გამოყოფა-იდენტიფიკაცია.
ჩრდილო ამერიკაში გავრცელებულ დროზოფილაში (D. pseudoobscura) უწინ აერთიანებდნენ ორ ქვესახეობას. გენეტიკური გამოკვლევებით დაადგინეს, რომ ისინი ორი რეპროდუქციულად იზოლირებული სახეობაა. ეს ორი სახეობა მორფოლოგიური ნიშნების მიხედვით (სანერწყვე ჯირკვლების ფორმა, ფრთების ზომა და სხვ.) უმნიშვნელოდ განსხვავდება. ასევე ეკოლოგიური თავისებურებებით განსხვავება უმნიშვნელოა. ერთი მათგანი სიცივის ამტანია, გავრცელებულია უფრო ჩრდილოეთით და მთიან რაიონებში. განსხვავდებიან სქესობრივი მომწიფების ვადებით: ერთი მწიფდება გამოჩეკიდან 32-36 სთ, მეორე კი 44-48 სთ-ის შემდეგ.
მორფოლოგიური კრიტერიუმით შეუძლებელია სახეობა-ორეულების გამოყოფა. სახეობა-ორეულები გარეგანი ნიშნებით თითქმის არ განსხვავდებიან, რადგან ევოლუცია სადიაგნოსტო ნიშნების დივერგენციის გზით არ მიმდინარეობდა. ასეთი ფორმები ხშირად გვხვდება უხერხემლო ცხოველებში, ხერხემლიანებში კი იშვიათია. იგი გამოვლენილია ამფიბიებში, ქვეწარმავლებში, ფრინველებში, ძუძუმწოვრებშიც (მცირე ზომის მღრღნელებში). ჩვეულებრივ მემინდვრიაში გვხვდება ორი სახეობა-ორეული. ერთს 46, მეორეს კი 54 ქრომოსომა აღენიშნება. ასევე შავ ვირთაგვაში ერთ სახეობას 38, ხოლო მეორეს 42 ქრომოსომა აქვს. ვინაიდან ზოგჯერ სახეობები მორფოლოგიურად მკაფიოდ არ არიან გამიჯნული, ხოლო ზოგიერთი მათგანი (სახეობა-ორეულები) პრაქტიკულად მორფოლოგიური ნიშნებით არ განსხვავდებიან, ამდენად, მხოლოდ ეს კრიტერიუმი არ არის საკმარისი იმის განსასაზღვრავად, თუ რომელ სახეობას ეკუთვნის ინდივიდი.
ფიზიოლოგიურ-ბიოქიმიური კრიტერიუმი. ამ კრიტერიუმის მიხედვით ერთსა და იმავე სახეობაში გაერთიანებულ ინდივიდებში მსგავსი ცხოველქმედების პროცესები უნდა მიმდინარეობდეს. გამოირკვა, რომ მონათესავე სახეობები ფიზიოლოგიური და ბიოქიმიური ნიშნებით ნაკლებ განსხვავდებიან. გარკვეული სპეციფიკური სახის მაღალმოლეკულური ორგანული ნივთიერებების სინთეზი დამახასიათებელია ცალკეულ სახეობათა ჯგუფებისათვის. მაგ., ალკალოიდების სინთეზით და დაგროვების უნარით ერთიმეორისაგან განსხვავდებიან ერთ ოჯახში (ძაღლყურძენასებრთა, შროშანისებრთა, ორქიდეისებრთა და სხვ.) გაერთიანებული სახეობები.
მოლეკულური ბიოლოგიის უახლესი მეთოდების გამოყენებით შესწავლილია ჩ ციტოქრომისა და სისხლის პიგმენტების სტრუქტურა ცხოველებში. გამოირკვა, რომ ჰემოგლობინის სტრუქტურა სისტემატიკურად ახლო მდგომ სახეობებში მსგავსია. იგივე კანონზომიერება ვლინდება მრავალი სპეციფიკური ცილის მიმართაც. ინსულინი მხოლოდ ქორდიანებშია აღმოჩენილი. ამ ჰორმონის სტრუქტურის შესწავლამ ცხადყო, რომ ამინომჟავური შემადგენლობა ყველა მათგანში თითქმის მსგავსია. მსგავსი შედეგებია მიღებული გენოსისტემატიკის მეთოდების გამოყენებითაც. მონათესავე სახეობათა დნმ ნუკლეოტიდების თანმიმდევრობით უმნიშვნელოდ განსხვავდებიან. ზოგიერთი ამინომჟავის (მაგ. ლიზინი) მეტაბოლიზმის გზა მონათესავე ცხოველებში განსხვავებულია, მაშინ როდესაც სისტემატიკურად დაცილებულ სახეობებში (E.coli, N.crassa) ზოგიერთი ამინომჟავას (მაგ., არგინინი, ჰისტიდინი) მეტაბოლიზმი ერთნაირია. ამრიგად, მხოლოდ ეს კრიტერიუმი არ არის საკმარისი სახეობის იდენტიფიცირებისათვის.
გეოგრაფიული კრიტერიუმი. სახეობას განსაზღვრული არეალი უჭირავს, ე.ი. მისთვის ხელსაყრელ პირობებში გვხვდება. სახეობის არეალის განსაზღვრულობა ეკოლოგიური პირობების დისკრეტულობაზე მიუთითებს. ამიტომაც მას ეკოლოგიურ კრიტერიუმთან აერთიანებენ და ეკოლოგიურ-გეოგრაფიულ კრიტერიუმს უწოდებენ. არეალის ფართობი, ფორმა და განლაგება ბიოსფეროში მნიშვნელოვანი სახეობრივი ნიშანია. ყოველ სახეობას თავისთვის გამოსადეგი ტერიტორია უჭირავს, აქვს წარმოშობის კერა. იგი სპეციფიკურ დამოკიდებულებას ამყარებს არეალში გავრცელებულ სხვა სახეობებთან. სახეობის დახასიათება არაა სრულყოფილი, თუ არ მიენიშნა არეალის სპეციფიკური თავისებურებები.
მაგრამ ამ კრიტერიუმსაც არაერთი ნაკლოვანება აღენიშნება, რის გამოც იგი სახეობის დასახასიათებლად უნივერსალური არ არის. არსებობენ ისეთი სახეობები, რომელთაც ერთი და იგივე არეალი გააჩნიათ. მაგალითად, ქონდარა ფიჭვს (Prunus pumila), მიდენდორფის არყს (Betula middendorfii ), დაურიის ლარიქსს (Larix daurica) და სურნელოვან ვერხვს (Populus suaveolens). ისინი მჭიდროდ არიან ერთმანეთთან დაკავშირებულნი და ერთსა და იმავე თანასაზოგადოებაში შედიან. ბუნებაში გვხვდება კოსმოპოლიტი სახეობები, რომლებიც გავრცელებულია მთელი დედამიწის ზედაპირზე ან მის უმეტეს ნაწილში. ზოგ მათგანს ესა თუ ის კონტინენტი ან აკვატორია უკავია. ამ შემთხვევაში გეოგრაფიული კრიტერიუმი თავის მნიშვნელობას კარგავს. კოსმოპოლიტი სახეობებია მაგ., დელფინი ორცინუსი (Orcinus orca), ჭაობის ბუ (Asio flammeus), ყორანი (Corvus corax), ლელი (Phragmites communis), წყლის მრავალძარღვა (Alisma plantagო), ჭინჭარი (Urtika dioica) და სხვ. სწრაფგანსახლებად სახეობებს შემოსაზღვრული არეალი არ გააჩნიათ. არეალი არ გააჩნია ადამიანის მიერ უნებურად გავრცელებულ სახეობებს (სახლის თაგვი, ვირთაგვა, ტარაკანა, ოთახის ბუზი და სხვ.) და რუდერულ მცენარეებს. ამრიგად, მხოლოდ გეოგრაფიული კრიტერიუმი არაა საკმარისი სახეობის განსასაზღვრავად.
გენეტიკური კრიტერიუმი სახეობის რანგის განსაზღვრისათვის მთავარი და ყველაზე სარწმუნო კრიტერიუმია. ამ კრიტერიუმის საფუძველია სახეობის გენეტიკური ერთიანობა. სახეობა არის დახშული რეპროდუქციულად იზოლირებული სისტემა. იგი შედგება ღია ქვესისტემებისაგან - პოპულაციებისაგან. რეპროდუქციული იზოლაცია ძირითადი დებულებაა სახეობის ბიოლოგიურ კონცეფციაში. სახეობაში გაერთიანებულ პოპულაციებს შორის მიმდინარეობს გენთა ნაკადი. იგი ცალკეული პოპულაციის იზოლირებული გენოფონდების ინტეგრაციას უზრუნველყოფს, რაც სახეობის ერთიანობას განაპირობებს (იხ. 7.3). არსებობს წყვეტილი არეალის მქონე სახეობები (მაგ., ცისფერი კაჭკაჭი, ხლაკუნა, ისლი და სხვ.), ასევე აგამური ფორმები, ობლიგატური პართენოგენეზური სახეობები. ფორმები, რომლებიც მხოლოდ თვითგანაყოფიერებით მრავლდებიან. ზემოთ დასახელებულ ფორმებს შორის გენეტიკური ინფორმაციის გაცვლა არ მიმდინარეობს და ინდივიდთა გენეტიკური ერთიანობა არ წარმოიქმნება.
მეორე მხრივ, ზოგჯერ ახლო მდგომ სახეობებში გაერთიანებული ინდივიდები ერთმანეთს ეჯვარებიან და ფერტილურ შთამომავლობას წარმოქმნიან. ასეთ სახეობებში დამოუკიდებელი გენეტიკური სისტემის რღვევა არ ხდება. ამდენად სახეობა წარმოადგენს გენეტიკურად მდგრად სისტემას.
თეორიულად შემუშავებული კრიტერიუმების პრინციპების პრაქტიკული გამოყენება გაძნელებულია. სახეობის დასადგენად ყველა კრიტერიუმის გამოყენება შეუძლებელია. კონკრეტულ შემთხვევაში ინდივიდების დაჯგუფების რანგის განსაზღვრა ხელმისაწვდომი კრიტერიუმებით ხორციელდება. მართალია, სახეობა დისკრეტული სისტემაა, მაგრამ თუ რომელ სახეობას ეკუთვნის ესა თუ ის ინდივიდი, მისი განსაზღვრა ყოველთვის არ არის ადვილი.
|
|
13.4 11.4. სახეობის სტრუქტურა არეალი |
▲ზევით დაბრუნება |
სახეობა არის სიცოცხლის ორგანიზაციის ერთ-ერთი იერარქიული დონე და ამასთანავე ძირითადი ტაქსონომიური ერთეული. სახეობის ელემენტარულ სტრუქტურულ ერთეულს პოპულაცია წარმოადგენს. სახეობის შემადგენელ სტრუქტურებს, რომლის ერთიანობა გენთა ნაკადით მიიღწევა, გენოფონდი განსაზღვრავს. სახეობისათვის ნიშანდობლივია სივრცობრივი სტრუქტურა ე.ი. მას გარკვეული არეალი უკავია. არეალი ეწოდება დედამიწის ზედაპირის (ხმელეთი ან აკვატორია) იმ ნაწილს, რომელიც ამა თუ იმ სახეობას უკავია. არეალში სახეობა არათანაბრადაა გავრცელებული. სახეობას მხოლოდ მისთვის დამახასიათებელი ადგილსამყოფელი უკავია. შემდგომში შესაძლოა სახეობა განსახლდეს და არეალი განივრცოს, ან პირუკუ ამოწყდომის გამო მნიშვნელოვნად შემცირდეს. არეალი სახეობის ერთ-ერთი მნიშვნელოვანი მახასიათებელია, მიუხედავად იმისა, რომ იგი მუდმივი და უცვლელი არ არის.
არეალი შეიძლება იყოს მთლიანი (უწყვეტი) ან წყვეტილი (დიზუნქციური). არეალი მთლიანია, როდესაც მთელ მის სივრცეზე სახეობას თავისი სასიცოცხლო მოთხოვნილების შესაბამისი ყველა ადგილსამყოფელი უკავია. აქვე უნდა აღინიშნოს, რომ ბუნებაში აბსოლუტურად უწყვეტი არეალი არ არსებობს; კერძოდ მჭიდროდ დასახლებული ტერიტორიები შედარებით მეჩხრად დასახლებულ ან დაუსახლებელ ტერიტორიებს ცვლიან. მაშასადამე სახეობა არეალის ფარგალში მისთვის შესაფერის ეკოლოგიურ პირობებში გვხვდება.
მცენარეებიდან მთლიანი არეალის ნათელ სურათს იძლევა ციმბირის ტაიგის შემქმნელი სახეობები: ციმბირული ფიჭვი (Pinus sibirica), ციმბირული ნაძვი (Picea obovata) და ციმბირული სოჭი (Abies sibirica); ევროპის ტერიტორიაზე გაბატონებული ევროპული ნაძვი (Picae excelsa); საქართველოს დასავლეთ ნაწილში მასიურად გავრცელებული აღმოსავლური ნაძვი (P. orientalis). ცხოველებიდან - კურდღელი (Lerus europaeus), ციყვი (Sciurus vulgaris), მელა (Vulpes vulpes), მგელი (Canis lupus) და სხვ.
წყვეტილი არეალის შემთხვევაში სახეობის გავრცელების არე იმდენადაა დაშორებული მეორისგან, რომ მათ შორის ყოველგვარი კონტაქტი (გენთა ნაკადი) გამორიცხულია. დიზუნქციურ არეალს ფლობს პრომეთესეული მემინდვრია (Prometheomys schaposchnikovi) რომელიც ოთხი იზოლირებული პოპულაციითაა წარმოდგენილი, მათგან ორი დიდ კავკასიონზე ბინადრობს, ორიც კი - მცირე კავკასიონზე. იგივე ითქმის არჩვის (Rupicarpa rupicarpa) შესახებ. იგი გავრცელებულია კავკასიონზე, პირინეებზე, კარპატებზე და ტავრიის მთებზე (სურ. 11.1). მათი არეალი მრავალი ათეული კილომეტრით არის დაშორებული. მცენარეებიდან შეიძლება დავასახელოთ კავკასიისა და ევროპის გარკვეულ ტერიტორიაზე გავრცელებული ბუერა (Petasites albus).
სურ. 11.1. ალპური დიზუნქციური არეალის ტიპური ნიმუშია არჩვის (Rupicarpa rupicarpa rupicarpa) გავრცელება. იგი გვხვდება ცენტრალური და სამხრეთ ევროპის მთიან რაიონებში, კავკასიასა და მცირე აზიაში. მეოთხეულ პერიოდში გამყინვარებამ არეალის დანაწევრება გამოიწვია (ა. იაბლოკოვი, 1987).
წყვეტილი არეალი რიგ შემთხვევაში სახეობის ისტორიას ასახავს. წარსულში ვრცელი არეალი იზოლირებულ ნაწილებად ან უკანდახევის გამო სადღეისოდ ორ ან მეტ დიზუნქციურ არეალად არის ჩამოყალიბებული. ლეისტოცენში კლიმატის მკვეთრმა ცვლილებამ (გამყინვარება) ცისფერი კაჭკაჭის (Cyanopica ciana) და თევზ ხლაკუნას (Misgurnus fossilis) არეალის დანაწევრება გამოიწვია. ეს სითბოს მოყვარული ცხოველები ევრაზიაში ფართოდ იყვნენ გავრცელებული. გამყინვარების შედეგად საკმაოდ ვრცელ ტერიტორიაზე გადაშენდნენ. ამჟამად ისინი დასავლეთ ევროპისა და შორეულ აღმოსავლეთშია შემორჩენილი (იხ. სურ. 7.4). კონტინენტთაშორისი დიზუნქციის სანიმუშოდ შეიძლება დავასახელოთ ქვედა დევონში ფართოდ გავრცელებული ორგვარადმსუნთქავი თევზები, რომლებიც უმნიშვნელო ფორმების სახით სადღეისოდ შემორჩენილია ავსტრალიის, აფრიკისა და სამხრეთ ამერიკის ზოგიერთ წყალსატევში. მცენარეებიდან მსგავს არეალს ფლობს ისლი (Sarex hostiana), რომელიც ევროპის გარკვეულ ტერიტორიაზე (მათ შორის კავკასიაში) გვხვდება, ასევე ჩრდილოეთ ამერიკის აღმოსავლეთ სანაპირო უკავია (იხ. სურ. 11.2).
სურ. 11.2. ისლის შარეხ ჰოსტიანა წყვეტილი არეალი. არეალის ერთი ნაწილი ევროპის კონტინენტზეა. მეორე კი ჩრდილოეთ ამერიკის აღმოსავლეთ სანაპიროზე. არეალის გაყოფა ამერიკის და ევრაზიის კონტინენტების დათიშვამ გამოიწვია (ნ. ტიმოფეევ-რესოვსკი და თანაავტ., 1977).
წყვეტილი არეალის ჩამომყალიბებლად რიგ შემთხვევაში ანთროპული ფაქტორი გვევლინება. ტიპურ მაგალითს სიასამურის (Martes zibelina) არეალი წარმოადგენს. ძვირფასი ბეწვის გამო ცხოველის უსისტემო რეწვამ არეალის ფრაგმენტაცია გამოიწვია (იხ. სურ. 7.5).

სურ. 11.3. რელიქტიური ფორმები: A - hateria (Sphenodon punctatus); B - latimeria (Latimeria chalumnae); C - oposumi (Didelphis marsupialis); D - ginkgo (Ginkgo biloba) (ა. იაბლოკოვი, ა. იუსუპოვი, 2004).
განასხვავებენ ვრცელ (ფართო) და ვიწრო არეალს. ფართო არეალს ხშირად უკავია მთელი კონტინენტი ან რამდენიმე კონტინენტი, მთელი ზღვა ან მთელი ოკეანე. ვრცელ არეალს ფლობს კოსმოპოლიტი სახეობები. ასე მაგალითად, ჭაობის ბუ, ყორანი, ჩვეულებრივი დელფინი, დელფინი ორცინუსი, მელა, მგელი, ჭინჭარი, წყლის მრავალძარღვა, წყლის ვაზი, ლელი და სხვ.
ვიწრო არეალი ენდემურ სახეობებს უკავიათ. ენდემები ეწოდება იმ სახეობებს, რომლებიც გავრცელებულია მხოლოდ განსაზღვრულ რეგიონში (რაიონი, ოლქი, ქვეყანა და მისთ.) და სხვაგან არსად არ გვხვდება. ენდემების არეალი შეიძლება განისაზღვროს მხოლოდ რამდენიმე მწვერვალით ან მღვიმით, კონტინენტიდან დიდად დაშორებული კუნძულით, რომელიმე ტბით და სხვ. ასე მაგალითად, კუნძულ მადაგასკარზე (გამოყოფენ ზოოგეოგრაფიულ ქვეოლქად) გავრცელებულ ძუძუმწოვართა 36 გვარიდან 32 გვარი ენდემურია, ხოლო ფრინველების 127 გვარიდან თითქმის ნახევარი. წმინდა ელენეს კუნძულზე (ატლანტის ოკეანე) მოსახლე უმაღლესი მცენარეების 85%25 ენდემური ფორმებითაა წარმოდგენილი. ბაიკალის ტბაში (აზია), ფაუნის სახეობათა რაოდენობის 75%25 ენდემურია. კავკასიაში, მათ შორის საქართველოში, მრავალი ენდემური სახეობაა გავრცელებული. მაგალითად, კავკასიური სალამანდრა, კავკასიური გველგესლა, კავკასიური შურთხი, კავკასიური როჭო, აღმოსავლეთ კავკასიური და დასავლეთკავკასიური ჯიხვი, მეგრული არყი, იმერეთის მუხა, ქართული ზამბახი, არმაზის მაჩიტა და სხვ.
უკიდურესი ვიწრო არეალი ნიშანდობლივია რელიქტური (ე.წ. პალეოენდემები) სახეობებისათვის. რელიქტებს უწოდებენ წარსულ გეოლოგიურ ეპოქებში ფართოდ გავრცელებულ ფლორისა და ფაუნის შემორჩენილ სახეობებს. ისინი შემონახულია მათი ფართოდ გავრცელების ეპოქისდროინდელ პირობების მსგავს გარემოში. რელიქტების სანიმუშო მაგალითია მცენარეებიდან გინკგო, ველვიჩია, არაუკარია, მაგნოლია; ცხოველებიდან - ლატიმერია, ჰატერია, ოპოსუმი, იხვნისკარტა, ექიდნა და სხვა (სურ. 11.3). საქართველოში გავრცელებული რელიქტებიდან აღსანიშნავია: ძელქვა, წყავი, პონტოური შქერი, წაბლი, მრგვალი წამალი, ბიჭვინთის ფიჭვი, ელდარის ფიჭვი, კავკასიური გველგესლა და სხვ. (დამატებით იხ. §3 და 4).
ტერიტორიული ურთიერთდამოკიდებულების მიხედვით სახეობები შეიძლება დაჯგუფდეს შემდეგნაირად: ალოპატრიული და სიმპატრიული. ალოპატრიულ სახეობებს უჭირავთ სხვადასხვა გამოცალკევებელი, გადაუფარავი არეალები, ხოლო სიმპატრიულ სახეობებს - გადამფარავი ან ერთი საერთო არეალი. ტერიტორიულ ურთიერთდამოკიდებულებას იზოლაციის ფორმები და სახეობათა წარმოშობა განსაზღვრავს (იხ. § 7.5 და 12).
|
|
13.4.1 სახეობის პოლიტიპურობა |
▲ზევით დაბრუნება |
ჯერ კიდევ დარვინი, სახეობის აღმავლობას ტერიტორიის განვრცობასა და ჯგუფებად დაყოფას უკავშირებდა. იგი აღნიშნავდა: „მეტად აყვავებული სახეობები ანუ მათ გაბატონებულიც შეიძლება ეწოდოს, - ისეთები, რომლებიც ფართოდაა გავრცელებული, მეტად დაქსაქსულია და ყველაზე მდიდარია ინდივიდებით, - ხშირად დასაბამს აძლევენ კარგად გამოხატულ სახესხვაობებს, რომლებიც ჩემი ღრმა რწმენით ჩანასახოვანი სახეობებია“.
სახეობაში დიფერენცირებული ჯგუფების არსებობის იდეამ შემდგომი განვითარება ევოლუციის სინთეზურ თეორიაში პოვა, სახეობის პოლიტიპურობის კონცეფციის სახით. ე. მაირის მიხედვით, სახეობა პოლიტიპურია თუ მასში ორი ან მეტი ქვესახეობა, რასა ან პოპულაციაა გაერთიანებული. მონოტიპური სახეობები ქვესახეობებს არ შეიცავენ. ცოცხალ ბუნებაში პოლიტიპური სახეობები ფართოდაა გავრცელებული, კერძოდ ფარულთესლოვანებში, ძუძუმწოვრებსა და ფრინველებში სადღეისოდ აღწერილი სახეობებიდან 75%25 პოლიტიპური აღმოჩნდა.
ქვესახეობა არის ლოკალურ პოპულაციათა ერთობლიობა, რომელსაც სახეობის არეალის გარკვეული ტერიტორია უჭირავს და ამავე სახეობის სხვა პოპულაციების ინდივიდებისგან მდგრადი მორფოლოგიური ნიშნებით განსხვავდება. ამდაგვარი ტაქსონომიური განსხვავება მხოლოდ იმ შემთხვევაშია სარწმუნო, როდესაც პოპულაციის ან პოპულაციათა ჯგუფის ინდივიდთა არანაკლებ 75%25 ფლობს სხვა პოპულაციებისგან განმასხვავებელ ნიშნებს. როდესაც ამ პირობას აკმაყოფილებს ქვესახეობა, მაშინ მას გამოყოფენ დამოუკიდებელ ტაქსონად. ქვესახეობას ტრინალური ნომენკლატურით აღნიშნავენ. მაგ., შუა რუსეთის მელა Vulpes vulpes vulpes, სტეპის მელა V.V. stepensis, ამიერკავკასიის მთის მელა V.V. alticola. ზოოლოგიურ სისტემატიკაში ქვესახეობა ყველაზე დაბალი ტაქსონომიური ერთეულია.
პოლიტიპური სახეობა არის კავკასიის ენდემი მთის შროშანი (Lilium monadelphum). მისი არეალი ჩრდილოეთ კავკასიას მოიცავს. იგი ორი ქვესახეობითაა წარმოდგენილი პირიქითა ხევსურეთში, თუშეთში, ხევსა და დაღესტნის სარტყელში გვხვდება ქვესახეობა საქართველოს მთის შროშანი (L.m. georgicum), იგი მეორე ფორმისგან განსხვავდება მორფოლოგიური ნიშნებით. კერძოდ, მას გააჩნია მუქი ყვითელი უკანგადაგრეხილი ყვავილსაფარი, მურა ფერის სამტვრეები და იისფერი დინგი (გაგნიძე, 2005).
ევრაზიაში გავრცელებული ციყვი (შციურუს ვილგარის) მრავალ ქვესახეობას ფლობს, რომელთა არეალი ერთიმეორეს არ ფარავს (იხ. სურ. 11.4) კარელიასა და კოლის ნახევარკუნძულზე გავრცელებულ ქვესახეობაში ხშირად გვხვდება წითელკუდა და წითელყურა ფორმები. უკრაინაში გავრცელებულ ფორმებს ზამთარში ბეწვი ყავისფრად აქვთ შეფერილი. საქართველოში მოსახლე ინდივიდების ზურგის მხარე მურა მორუხო ჩალისფერია, მოშავო მურა წინწკლებით, კუდი მუქი წაბლისფერ-ჟანგისფერია. ციყვში გაერთიანებული ფორმები ალოპატრიული ქვესახეობებია.
სურ. 11.4. ევრაზიაში ციყვის (Sciurus vulgaris) ქვესახეობის არეალი (ლ. კაიდანოვი, 1996).
სიმპატრიული ქვესახეობების სანიმუშო მაგალითია ერთწლოვანი ბალახოვანი მცენარე გილია (Gilia achilleifolia). მასში ორი ქვესახეობაა გაერთიანებული რომელიც კალიფორნიის ვირჯინულ მუხნარშია გავრცელებული. ერთი მათგანი ჯვარედინ მტვერიაა (მტვერავენ ფუტკრები). G.a. achilleifola სავანის ტიპის ტყის შემოფარგლულ მზით განათებულ ველებზე იზრდება. გააჩნია მოლურჯო-იისფერი თავაკი. მეორე ქვესახეობა კი (G.a. multicaulis) მეჩხერტყეშია გავრცელებული. ის იკეთებს ღია შეფერილობის თავაკებს და არის თვითმტვერია. ვრცელ ტერიტორიაზე ტყიანი და ღია ადგილები მონაცვლეობს, რაც შესაბამისად გილიას ორივე ქვესახეობის გავრცელების ხასიათს განსაზღვრავს.
ბოტანიკურ ტაქსონომიაში ლოკალური ეკოლოგიური რასის სახით გამოყოფენ ეკოტიპს. ტერმინი შემოიტანა 1922 წ. შვედმა ბოტანიკოსმა გ. ტურესონმა. ეკოტიპი არის სახეობაში გაერთიანებულ მცენარეთა ჯგუფი, რომელთაც გააჩნიათ მემკვიდრულად განსაზღვრულ ნიშანთა კომპლექსი და შეგუებულია განსაზღვრულ საარსებო გარემოსთან - ბიოტოპთან. მაგალითად თურქესტანის ღვიაში აღწერილია მაღალმთის დაბალტანიანი სახესხვაობა (Varietas) - Juniperus turcestanica var. fruticosa.. ეკოტიპი შეიძლება იყოს კლიმატური - (წარმოქმნილია კლიმატური ფაქტორების მოქმედებით), ედაფური (შეგუებულია სპეციფიკურ ნიადაგურ პირობებთან), ცენოტური (განსაზღვრულია მცენარეული თანასაზოგადოებით).
ქართულ მუხაში (Quercus iberica), რომლის არეალი მოიცავს კავკასიას, ლაზისტანსა და ჩრდილოეთ ირანს. საქართველოში ტიპური ფორმის გარდა გვხვდება სამი სახესხვაობა. თრიალეთზე მშრალ კლიმატურ პირობებში ჩამოყალიბდა გრძელყუნწიანი მუხა (Quercus iberica var. longipedunculata) და კრებადნაყოფებიანი მუხა (Q.i. var. sorocarpa), ხოლო იმერეთისა და აფხაზეთის კირქვიანებზე კოზლოვსკისეული მუხა (Q.i. var. kozlowskyi) (გაგნიძე, 2000).
კლიმატური ეკოტიპის სანიმუშო მაგალითია Atriplex litorale ფორმები. შვეციის ქარისაგან დაცულ სამხრეთ სანაპიროზე გავრცელებულ მცენარეებს გააჩნიათ მაღალი სწორმდგომი ღერო. დასავლეთის სანაპიროებზე, სადაც ქარი გამუდმებით ქრის, დაბალტანიანი გვერდზე განტოტვილი მცენარეებია გავრცელებული. სამხრეთ რაიონებში გავრცელებული ფორმებისგან განსხვავებით შვეციის ჩრდილოეთში სადაც მკაცრი კლიმატური პირობებია მცენარეებს გააჩნიათ მოკლე სავეგეტაციო პერიოდი და ადრე ყვავილობენ.
„კვებითი“ რასები გამოვლენილია ფიტოფაგ მწერებში. ისინი კონკრეტულ პატრონ მცენარეზე სახლობენ. ვიწროდ შეგუებული ფიტოფაგი მწერი ზოგჯერ იცვლის პატრონ მცენარესა და იფართოებს კვებით „რაციონს“. ვაშლის ნაყოფჭამიაში, რომელიც შემთხვევით შეტანილ იქნა ჩრდილოეთ ამერიკაში, რამდენიმე წლის შემდეგ ჩამოყალიბდა ახალი რასა, რომელიც კაკლისა და მსხლის ნაყოფს აზიანებს. მსგავსი მაგალითების დასახელება მრავლადაა შესაძლებელი.
მტკნარი წყლის თევზებში (ორაგულისებრნი, ზუთხისებრნი, კობრისებრნი...) გამოვლენილია სეზონური ე.წ. „საგაზაფხულო“ და „საშემოდგომო“ რასები. ისინი განსხვავდებიან ტოფობის პერიოდით. ორხიდისა (მაკედონია) და სევანის (სომხეთი) ტბაში გავრცელებულ კალმახში გამოყოფენ ეკოლოგიურ იზოლატებს, რომლებიც განსხვავდებიან ტოფობის პერიოდით, საქვირითე ადგილით და სიღრმით.
სპეციფიურ ბიოტოპს (ეკოლოგიურად ერთგვაროვანი ტერიტორიის მონაკვეთი, რომელიც წარმოადგენს ამა თუ იმ ინდივიდთა დაჯგუფების ადგილს) ირჩევენ მეგაზაფხულე ყარანის, ვერცხლისფერი თოლიას, ჩხართვის რასები. ზაზუნას (Peromyscus polinotus) ორი განსხვავებული: ღია და მუქი შეფერილობის ფორმა გავრცელებულია შესაბამისად ღია ქვიშნარ და მუქ სუბსტრატზე.
საჭიროა აღინიშნოს, რომ შიგასახეობრივი დიფერენცირება თვით ფართოდ გავრცელებული სახეობებისათვისაც კი არ წარმოადგენს აუცილებელ პირობას. ამდაგვარი სიტუაცია ფრინველთა იმ დისპერსიული სახეობისთვისაა ნიშანდობლივი, რომელთა იუვენილური ფორმები გამოჩეკვის ადგილიდან შორ მანძილზე მიგრირებენ. გარეული იხვი (Anas platyrhynchus) ევრაზიისა და ჩრდილოეთ ამერიკის ვრცელ ტერიტორიაზე ბინადრობს. ეს მიმომფრენი ფრინველი ქვესახეობად არ არის დიფერენცირებული. მხოლოდ გრენლანდიის ჩრდილო-დასავლეთ ნაწილში სახლობს მობუდარი ფორმა. სხვა ფორმებისგან განსხვავებით მათ კარგად განვითარებული თვალ-ცხვირის ჯირკვალი გააჩნიათ, ვინაიდან რეგულარულად ზღვის მლაშე წყალს იყენებენ სასმელად.
ბეღურასნაირების ზოგიერთი სახეობა, მათ შორის დიდი წივწივა, მეგაზაფხულე ყარანი და თეთრწარბა დისპერსიულ ტიპს მიეკუთვნება. მათთვის ნიშანდობლივია მაღალი ეკოლოგიური პლასტიურობა, კერძოდ იკვებებიან ნაირგვარი საკვებით, გააჩნიათ გაწელილი გამრავლების პერიოდი და სხვადასხვა ადგილებში ბუდობენ.
|
|
13.4.2 სახეობის პოპულაციური სტრუქტურა |
▲ზევით დაბრუნება |
ადგილობრივ პოპულაციათა ერთობლიობით იქმნება ეკოტიპი, ეკოლოგიური რასა და ქვესახეობა. მათი რაოდენობა სახეობის განსახლების ფართობზე და შიგასახეობრივ დიფერენცირებაზეა დამოკიდებული. სახეობის პოპულაციური სტრუქტურის შესწავლის საწყის ეტაპზე საჭიროა დადგინდეს პოპულაციათა შორისი საზღვარი, ხოლო შემდგომ მათ შორის არსებული განსხვავება (იხ. §3 და 4). პოპულაციები ჩვეულებრივ სხვადასხვა მოცულობის უწყვეტ რიგებს ქმნიან. მომდევნო ეტაპზე არკვევენ პოპულაციათა თანმიმდევრულ რიგებში ცვალებადობის ხარისხს. განსაკუთრებული ყურადღება ეთმობა სახეობის ერთიან არეალში განმხოლოებული პოპულაციების - გეოგრაფიული იზოლატების (თუ ამდაგვარი მოეპოვებათ) გამოვლენას. ასევე მნიშვნელოვანია გამიჯნულ პოპულაციათა ან ქვესახეობათა საზღვარზე მკვეთრად გაზრდილი ცვალებადობის ადგილის - ჰიბრიდული ზონის გამოვლენა. „კუნძულებრივად“ დასახლებულ (ფართო მნიშვნელობით) ან ძნელად გადასალახავი ბუნებრივი ზღუდეებით გამიჯნულ პოპულაციებს შორის საზღვარი მკაფიოდ არის გამოხატული. საზღვრის განსაზღვრისას გასათვალისწინებელია ცხოველთა გადაადგილებისა და გამრავლების თავისებურებები, მცენარეებში სპორებისა და თესლების გავრცელების შესაძლებლობები. არსებითია რეპროდუქტული არეალი, რომლითაც პოპულაციები მეტნაკლებად გამიჯნული უნდა იყვნენ ე.ი. გენთა ნაკადი მინიმალური სიხშირით უნდა მიმდინარეობდეს.
ფართო არიალის მფლობელ სახეობებში პოპულაციათა შორის მკვეთრი საზღვრის გავლება გაცილებით რთულია. გარდამავალი ფორმების არსებობა არ იძლევა საზღვრის გავლების საშუალებას. ხშირად ამა თუ იმ გეოგრაფიული (ჰორიზონტალური ან ვერტიკალური) მიმართულებით ურთიერთთანამიმდევრულად განლაგებულ პოპულაციათა რიგში შეინიშნება ნიშან-თვისებათა თანდათანობითი ცვლილება. კიდურა პოპულაციების წევრები მკვეთრად განსხვავდებიან ერთმანეთისგან. მოსაზღვრე პოპულაციათა უწყვეტ რიგში რომელიმე ნიშნის (გრადიენტი) თანდათანობით ცვლილებას კლინური ცვალებადობა ეწოდება.
კლინები (ნიშანთა გრადიენტები) აღენიშნება თითქმის ყველა კონტინენტურ სახეობას. მათი წარმოქმნა გამოწვეულია კლიმატური და სხვა გარემო ფაქტორების სივრცობრივი ცვლილებებით, ასევე მიგრაციული პროცესის თანამდევი გენთა ნაკადით. რაც უფრო უახლოვდება პოპულაცია სახეობისათვის ნიშანდობლივ საზღვარს, მით მკვეთრად ვლინდება გარემოს არახელსაყრელი პირობების ზემოქმედება. დგება ისეთი ზღვარი, რომლის იქითაც შეუძლებელია სახეობამ განავრცოს საზღვარი. ამდენად სახეობის არეალის საზღვართან მეტწილად ყალიბდება გეოგრაფიული იზოლატები. ჩვეულებრივ საზღვრისპირა პოპულაციები არის მცირე ზომის, ნაკლებმდგრადი და შემადგენლობითაც ღარიბი.
გეოგრაფიული იზოლატი წარმოადგენს პოპულაციას ან პოპულაციათა ჯგუფს, რომელიც სივრცობრივად განცალკევებულია და ბუნებრივი ზღუდეები აბრკოლებენ გენთა მიმოცვლას ამავე სახეობის სხვა პოპულაციასთან. მუტაციური პროცესი მოქმედებს პოპულაციის დინამიკაზე და ხელს უწყობს მიკროევოლუციური პროცესის მიმდინარეობას. ამ სახით იზოლატები ევოლუციის ერთგვარ „მზვერავადაც“ გვევლინებიან, რამდენათაც ისინი არაორდინალურ, ხშირად ექსტრემალურ პირობებში იმყოფებიან. გეოგრაფიული იზოლატების სანიმუშო მაგალითს ოკეანური კუნძულები წარმოადგენს. ხშირად მათი გავრცელება ცალკეული კუნძულით შემოიფარგლება. სოლომონის კუნძულზე (წყნარი ოკეანე) გავრცელებული ფრინველების 52%25 იზოლატების სახითაა წარმოდგენილი. ხვლიკი (Lacerta muratis) პირინეის ნახევარკუნძულის ვრცელ ტერიტორიაზე მხოლოდ სამი რასითაა წარმოდგენილი, ხოლო ბალიარისა და პიტიუზის კუნძულზე (ხმელთაშუა ზღვა) იმავე სახეობის 50 ერთმანეთისგან კარგად გამიჯნული რასით სახლობს. მსგავსი მაგალითების ჩამონათვალი მნიშვნელოვნად შეიძლება განივრცოს.
ამრიგად, სახეობა რთულ კომპლექსურ სისტემას წარმოადგენს. მასში რიგითობის მიხედვით გაერთიანებულია ურთიერთმოქმედი კომპონენტები (ნაწილები). სახეობის მნიშვნელოვანი ერთეული არის პოპულაცია. პოპულაციის შიგნით შეიძლება ჩვეულებრივ გამოიყოს გენეტიკური ნათესაობით ურთიერთდაკავშირებული დროებითი დაჯგუფებები. ასეთ ერთობლიობას ცხოველებში დემებს, მცენარეებში ბიოტიპებს უწოდებენ.
სურ. 11.5. სახეობის - მრავალგანზომილებიანი იერარქიული სისტემის აგებულების სქემა (ა. ბარნოვი, 1986).
თუ ზეპოპულაციურ რიგებს განვიხილავთ მაშინ ცხოველებში რასებსა და ქვესახეობებს, ხოლო მცენარეებში ეკოტიპებსა და ქვესახეობებს გამოვავლენთ (იხ. სურ. 11.5). სისტემატიკასა და ევოლუციურ ბიოლოგიაში შიგასახეობრივი იერარქიული რანგების ერთიანი (ყველა სამეფოსათვის საერთო) ნომენკლატურა სადღეისოდ არ არის შემუშავებული. მაგრამ სახეობის პრობლემებზე მომუშავე ყველა ბიოლოგი ერთნაირი თვალსაზრისისაა; რომ სახეობა არის დროსა და სივრცეში ინტეგრირებული მრავალი დონის შემცველი ბიოქოროლოგიური ჯგუფების რთული იერარქიული სისტემა, რომლის ერთიანობა გენთა ნაკადით მიიღწევა.
|
|
13.5 11.5. სახეობა ევოლუციის ეტაპია |
▲ზევით დაბრუნება |
ნებისმიერი სახეობა წარმოადგენს ორგანიზაციის სხვადასხვა დონეზე მყოფი იერარქიული რანგებისაგან შემდგარ რთულ სისტემას, ხოლო რანგებს შორის ინტეგრაცია გენეტიკური ინფორმაციის გაცვლის მეშვეობით ხორციელდება. მიუხედავად იმისა, რომ სახეობაში თავს იჩენს პოლიმორფიზმი და ჰეტეროგენულობა, სხვადასხვა იერარქიული დაჯგუფების არსებობისას, იგი ევოლუციური პროცესის უმცირესი დისკრეტული გენეტიკურად მდგრადი სისტემაა. სახეობის ბიოლოგიური არსი იმაში მდგომარეობს, რომ მას აქვს ნაირგვარი საიზოლაციო მექანიზმებით სხვა სახეობების გენთა ნაკადისაგან დაცული გენოფონდი. ერთი მხრივ სახეობა, როგორც ევოლუციის შედეგი, ერთიანი და მთლიანი სტაბილური სისტემაა* მეორე მხრივ იგი, როგორც ევოლუციური პროცესის საწყისი ეტაპი, დინამიურია, მკაფიო საზღვრები არ გააჩნია და ლაბილურია.
ნებისმიერი სახეობა შედგება დისკრეტული, დამოუკიდებელი საკუთარი ევოლუციური ბედის მქონე ერთეულებისაგან - პოპულაციებისაგან. პოპულაციათა შორის კავშირი გენთა ნაკადით მყარდება, რის შედეგადაც ერთიანი რთული სისტემა წარმოიქმნება. გენთა ნაკადი ხელს უშლის ცალკეულ პოპულაციასა და მის ჯგუფებში მნიშვნელოვანი ცვლილებების დაგროვებას. მისი მეშვეობით სახეობა რთული ინტეგრირებული სისტემის სახით არსებობს.
როდესაც პოპულაციათა შორის გენთა ნაკადი წყდება და მისი აღდგენა ხანგრძლივი დროის განმავლობაში არ ხდება, მაშინ ორი განსხვავებული პოპულაციური ღია სისტემიდან ორი დამოუკიდებელი განსხვავებული სისტემა წარმოიქმნება, ანუ ადგილი აქვს ორი განსხვავებული სახეობის ჩამოყალიბებას. ახალი სახეობის წარმოშობით მთავრდება ცოცხალ ბუნებაში გრანდიოზული და მუდმივ მიმდინარე ევოლუციური პროცესის ერთ-ერთი უმნიშვნელოვანესი ეტაპი და იწყება მეორე ეტაპი. ამდენად ნებისმიერი სახეობა იმავდროულად არის ევოლუციური პროცესის თვისებრივი ეტაპი.
|
|
14 § 12. სახეობათა წარმოქმნა - მიკროევოლუციის შედეგი |
▲ზევით დაბრუნება |
საწყისი სახეობიდან ერთი ან რამდენიმე ახალი სახეობის ჩამოყალიბების პროცესს სახეობათა წარმოქმნა ეწოდება. სახეობათა წარმოქმნა არის გენეტიკურად ღია სისტემის (პოპულაცია) დახშულ სისტემად გარდაქმნის პროცესი. სახეობათა წარმოქმნა მიმდინარეობს სახეობის შიგნით მუდმივად მოქმედი მიკროევოლუციის პროცესის შედეგად. სახეობათა წარმოშობაზე დაკვირვება ბუნებაში ძალზე ძნელია, რადგან მისი ხანგრძლივობა საკმაოდ დიდია. მის მექანიზმზე მხოლოდ თეორიული მსჯელობა შეიძლება. სახეობათა წარმოქმნის ძირითადი და ერთადერთი მიმართული ფაქტორი არის მიმმართველი ბუნებრივი გადარჩევა. სახეობათა წარმოქმნა მიკროევოლუციის საბოლოო შედეგია. იგი სახეობის შიგა დონეზე პოპულაციების დიფერენცირებით იწყება, რომელიც დივერგენციის შედეგად ახალ სახეობად გარდაიქმნება. სახეობის არეალში ელემენტარული ევოლუციური ფაქტორების მოქმედების შედეგად სხვადასხვა პოპულაციებში მიმდინარეობს ელემენტარული ევოლუციური მოვლენა-პოპულაციის გენეტიკური სტრუქტურის (გენოფონდის) მდგრადი, შეუქცევადი მიმართული ცვლილება. ევოლუციის ელემენტარული ფაქტორების ზემოქმედებით ასეთი ცვლილებები შეიძლება აკუმილირდეს პოპულაციებში, რის შედეგადაც სხვაობა ღრმავდება, ამას კი ქვესახეობის წარმოქმნა მოჰყვება. იგი ახალი სახეობების წარმოქმნის წინა ეტაპს წარმოადგენს. იზოლაციის ხანგრძლივი მოქმედების გამოგენთა ნაკადი წყდება, რის გამოც ასეთი პოპულაციები ან ქვესახეობები ახალ სახეობად გარდაიქმნებიან.
|
|
14.1 12.1. სახეობათა წარმოქმნის კონცეფციები |
▲ზევით დაბრუნება |
თანამედროვე ეტაპზე საყოველთაოდ არის აღიარებული ევოლუციის სინთეზური თეორია. თავდაპირველად განვიხილოთ ამ თეორიის ძირითადი კონცეპტუალური დებულებები. ევოლუციის სინთეზური თეორიის მიხედვით სახეობა წარმოადგენს ბიოლოგიის ფუნდამენტურ კატეგორიას, სახეობათა წარმოქმნის პროცესი კი - ევოლუციის საკვანძო ეტაპს.
სახეობა - დახშული უნიკალური გენეტიკური სისტემაა, რომელიც სხვადასხვა საიზოლაციო მექანიზმებით დაცულია სხვა გენოფონდების გენთა ნაკადისაგან. პოლიტიპური სახეობა შეიცავს ქვესახეობებს, რასებს და პოპულაციებს, რომლებსაც აქვთ დამოუკიდებლად არსებობის უნარი. ამასთან ერთად ნებისმიერი სახეობა ისტორიულად ჩამოყალიბებული მთლიანი სისტემაა. პოპულაცია არის ევოლუციის ელემენტარული ერთეული, რომელსაც კონკრეტულ ეკოსისტემაში აქვს ბუნებრივ-ისტორიული ბედი, ამასთანავე კვებით ჯაჭვებსა და ენერგიის ნაკადში მისთვის ნიშანდობლივი ადგილი უკავია. პოპულაცია წარმოადგენს ღია გენეტიკურ სისტემას, რომელიც გენთა ნაკადით ანალოგიურ სისტემებთან არის დაკავშირებული. მიმდინარეობს გენეტიკური ინფორმაციის ურთიერთცვლა. მაშასადამე, გენთა ნაკადი არსებით როლს ასრულებს, სახეობის როგორც ევოლუციის ეტაპის, მდგრადობის შენარჩუნებაში. გენთა ნაკადის შეწყვეტა ახალ სახეობათა ფორმირების ერთ-ერთ საწყის წინაპირობად არის მიჩნეული.
კონკრეტულ სახეობაში გაერთიანებული ინდივიდის გენოტიპი ევოლუციის პროცესში ჩამოყალიბებული გენთა ბალანსირებული სისტემაა. ნებისმიერი სახის მუტაცია ამ სისტემის დესტაბილიზაციას ახდენს. მუტაციის დამაზიანებელი ეფექტი ალელთშორისი (სრული და არასრული დომინირება, ზედომინანტობა) და გენთაშორისი (ეპისტაზი, სუპრესია, პოლიმერია) ურთიერთქმედებით მცირდება. ბუნებრივი გადარჩევის მოქმედებისათვის ყველაზე ხელსაყრელ მასალად მიჩნეულია პირობით საზიანო მცირე მუტაციები. ახალ საარსებო გარემოში ალელთა და გენთაშორისი ურთიერთქმედებით მუტანტური გენის კორექცია ხორციელდება. მისი საზიანო ეფექტი შეიძლება შესუსტდეს ან სასარგებლო გახდეს. არსებითი მნიშვნელობა მუტირებულ გენზე გენოტიპური გარემოს ზემოქმედებას ენიჭება.
მუტაციური პროცესისა და რეკომბინოგენური მოქმედების შედეგად შიგასახეობრივი დიდი „სამობილიზაციო რეზერვი“ წარმოიქმნება. პანმიქსია, აგრეთვე სპეციფიკური გენეტიკური მექანიზმები (მაგალითად, ზედომინანტობა), პოპულაციაში გენეტიკური მასალის მდგრად წონასწორობას უზრუნველყოფენ.
ბუნებრივი გადარჩევის მოქმედებით მიიღწევა პოპულაციის გენეტიკური სტრუქტურის დიფერენცირება. ფლუქტუირებადი საარსებო გარემოსადმი (სეზონური ციკლური ცვლილებები, განათება, ტემპერატურა, ქარი და მისთ.) განსხვავებული ადაპტური ღირებულების გენოტიპური კლასები ყალიბდება. პოპულაციის გენეტიკური ჰეტეროგენურობითა და პოლიმორფულობით მისი გარდაქმნის წინაპირობა იქმნება.
ხშირად პოპულაციის საზღვრები ბიოქოროლოგიური ერთეულის - ეკოსისტემის საზღვრებს ემთხვევა. სივრცობრივი იზოლაციის განმტკიცება პოპულაციების განკერძოებულად დიფერენცირებას განაპირობებს. სახეობის პოპულაციური სტრუქტურა მის ბიოლოგიურ პოტენციას და აღმავლობითი დონის უნარს გამოხატავს. ამასთანავე, იგი ფლობს შიგასახეობრივ რანგებად, შემდგომ კი ახალ სახეობად დივერგირების წანამძღვრებს.
ევოლუციის სინთეზური თეორიის მიხედვით სახეობის წარმოშობის კლასიკური სქემა ამგვარია: ცალკეული პოპულაციის ან პოპულაციათა ჯგუფის სივრცობრივად გამიჯვნა; მიმართული გადარჩევით გენოფონდის ვექტორიზებული და შეუქცევადი ცვლილება; ქვესახეობის (გეოგრაფიული რასის), შემდგომ სახეობის დიფერენცირება და თანმხლები პროცესის - რეპროდუქციული იზოლაციის ფორმირება; მთლიანად ეს პროცესი თანდათანობით - გრადუალურად, ხანგრძლივად მიმდინარეობს (იხ. სურ. 12.1) დიფერენცირების პროცესი ნებისმიერ ეტაპზე შეიძლება შეწყდეს. ამიტომაც პოპულაციათა მეტად მცირე ჯგუფი ყალიბდება ქვესახეობად, მხოლოდ ერთეულები
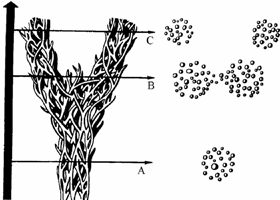
სურ. 12.1. სახეობათა წარმოშობის სქემა. ცალკეული ტოტი შეესაბამება პოპულაციას. A - საწყისი სახეობის დონე. B - სახეობების გამიჯვნის მომენტი. C - ორი ახალი სახეობა (ფ. დობჟანსკი, 1957).
იძენენ სახეობრივ სტატუსს. სახეობათა წარმოქმნა განაპირობებს ბიომრავალფეროვნებას. ევოლუციის სინთეზური თეორიის თვალთახედვით სახეობა ევოლუციის ქვაკუთხედია. ევოლუციის სინთეზური თეორია ჩამოყალიბდა თანდათანობით (იხ. §1). ბოლო ათწლეულის მანძილზე კი მნიშვნელოვნად შეიცვალა და დაიხვეწა. განვითარების საწყის ეტაპზე პოპულაციონისტები მხოლოდ სახეობათა წარმოქმნის ალოპატრიულ მოდელს აღიარებენ. სიმპატრიული მოდელის წინააღმდეგ მოსაზრებები მთელი სისრულით წარმოდგენილია გამოჩენილი ევოლუციონისტის ე. მაირის ნაშრომში „ზოოლოგიური სახეობა და ევოლუცია“ (1968). მისი შეხედულებით სიმპატრიული კონცეფცია საჭირო აღარ არის, რადგან მასში იგნორირებულია სახეობათა წარმოქმნა პოპულაციურ დონეზე. სახეობათა წარმოქმნის ძირითადი ეტაპი - პოპულაციის გენეტიკური სტრუქტურის გარდაქმნა - მხოლოდ მაშინ მიმდინარეობს, თუ იგი დროებით უცხო გენთა ნაკადიდან არის დაცული. მას ადვილად ახორციელებს გარეგანი ბარიერი - სივრცობრივი იზოლაცია.
ე. მაირის შეხედულება მრავალმა ევოლუციონისტმა გაიზიარა. ზოგიერთმა კი არა. დისკუსია განსაკუთრებით გამწვავდა XXს. 70-იან წლებში, როდესაც ამერიკელმა პალეობიოლოგებმა ნ. ელდრიჯმა და ს. გულდმა სახეობათა წარმოქმნის „წყვეტილი წონასწორობის“ ანუ პუნქტირალიზმის (განახლებული სალტაციონიზმი) მოდელი წარმოადგინეს. იგი ემყარება ტიპოლოგიურ კონცეფციას. პუნქტურალიზმის კონცეფციის მიხედვით სახეობას განვითარების პროცესში აღენიშნება ხანგრძლივი მოსვენების პერიოდი - სტაზისი. ეს პერიოდი დაახლოებით 1015-1017 თაობას მოიცავს. იგი იცვლება ნახტომისებური (სალტაციური) ცვლილების პერიოდით, რომელიც საშუალოდ 103-104 თაობას შეადგენს. ამ პერიოდში ხდება ახალ სახეობათა ჩამოყალიბება. ახალი სახეობრივი ფორმები სათავეს რამდენიმე ინდივიდიდან - მაკრომუტანტიდან იღებს (იხ. §14).
გრადუალიზმი თუ სალტაციონიზმი, ამ დილემის გარშემო კამათს საკმაოდ დიდი ხნის ისტორია აქვს. იგი დარვინის დროიდან იღებს სათავეს. ს. გულდის შეფასებით დარვინი გრადუალური გზით სახეობის ჩამოყალიბებას ისეთსავე პირველხარისხოვან მნიშვნელობას ანიჭებდა, როგორსაც ბუნებრივი გადარჩევის თეორიას.
ჰ. დე ფრიზის მუტაციური თეორია ეწინააღმდეგებოდა დარვინის იდეას (იხ. § 1). მოგვიანებით პოპულაციურმა გენეტიკამ, რომელსაც ეყრდნობა ევოლუციის სინთეზური თეორია, ერთგვარი მოდიფიცირებით გაიზიარა მუტაციონიზმის იდეა. კერძოდ, ბუნებრივი გადარჩევისთვის მასალას ე.წ. „მცირე“ მუტაციები წარმოადგენს. ევოლუციის სინთეზური თეორიის მიხედვით მუტაციური პროცესი ელემენტარული მასალის მიმწოდებელი ფაქტორია, ხოლო ინდუცირებული მუტაციები - ელემენტარული მასალა (იხ. § 5.2 და 7.1).
XXს. 40-იან წლებში გამოჩენილი მეცნიერი რ. გოლდშმიტი, გენეტიკოსებს შორის პირველი, დაბეჯითებით მიუთითებდა მაკრო - ანუ სისტემური მუტაციების დიდ როლზე ევოლუციაში. ეს მოსაზრება თითქმის არავინ გაიზიარა. მაგრამ ბოლო წლებში ამ იდეამ აქტუალობა შეიძინა.
სალტაციური გზით სახეობათა წარმოქმნა შეიძლება განისაზღვროს შემდეგნაირად: საწყისი სახეობიდან რეპროდუქციულად იზოლირებული ინდივიდის ან ინდივიდთა მონათესავე ჯგუფის ჩამოყალიბება, რომელსაც გააჩნია პოტენცია დამოუკიდებელ სახეობად ჩამოყალიბებისა. ახალი ტაქსონების სალტაციის შედეგად წარმოქმნას ხშირად მაკროგენეზს უწოდებენ. სალტაციონიზმის ანუ მაკროგენეზის კონცეფცია უპირისპირდება ევოლუციის სინთეზურ თეორიას. უპირველეს ყოვლისა ეს ეხება თეზას: „პოპულაცია, როგორც ევოლუციის ელემენტარული ერთეული“. ე.მაირის შეხედულებით მაკროგენეზი გულუბრყვილო კონცეფციაა, ვინაიდან თვალნათლივ ვერ ასაბუთებს (შეუძლებელიცაა!) ევოლუციაში ისეთი მნიშვნელოვანი სალტაციის არსებობას, რომელიც ერთი მხრივ, რეპროდუქციულ იზოლაციას, მეორე მხრივ, ეკოლოგიურ შეთავსებადობას განაპირობებს.
ამ კონცეფციათა შორის პრინციპული სხვაობა არის აგრეთვე სახეობათა წარმოქმნის პროცესის ინტერპრეტაციაში. ევოლუციის სინთეზური თეორიის თანახმად სახეობათა ჩამოყალიბება პოპულაციათა დონეზე მიმდინარეობს. მაკროგენეზის მიხედვით, ახალ სახეობათა წარმოშობა ხორციელდება სტაბილიზაციის ფაზის გავლის შემდეგ ერთ ან რამდენიმე ინდივიდში მომხდარი დიდი გენეტიკური რეორგანიზაციის შედეგად.
ცნობილი ევოლუციონისტი ნ.ვორონცოვი (2004) მიიჩნევს, რომ გრადუალიზმსა და სალტაციონიზმს შორის დაპირისპირება დასრულდება მრავალ პრობლემაზე ერთიანი შეხედულების შემუშავებით. დისკუსიებმა თვალნათლივ გვიჩვენა, რომ ბუნებაში სახეობათა წარმოქმნის ორივე ფორმა (გრადუალიზმი და სალტაციონიზმი) მოქმედებს ე.ი. თანაარსებობს სახეობათა მყისიერად და თანდათანობითი ჩამოყალიბების მექანიზმები.
|
|
14.2 12.2. სახეობათა წარმოქმნა, როგორც ბიომრავალფეროვნების სათავე |
▲ზევით დაბრუნება |
ბიომრავალფეროვნება ორგანული ბუნების ერთ-ერთი განსაკუთრებული თვისებაა. ამ მრავალფეროვნების საოცარი თავისებურება სისტემათა დისკრეტულად არსებობაში მდგომარეობს. ორგანული ბუნება ნაირგვარი და მრავალი დისკრეტული ერთეულებისგან - სახეობებისგან შედგება. ვიდრე სახეობაში გაერთიანებულ განსხვავებულ პოპულაციებს შორის მიმდინარეობს გენეტიკური ინფორმაციის მიმოცვლა (გენთა ნაკადი, იმიგრაცია - ემიგრაცია), მანამდე იგი ერთიანი რთულად ინტეგრირებული სისტემაა. როდესაც პოპულაცია ან პოპულაციათა ჯგუფი საიზოლაციო მექანიზმებით ხანგრძლივად რჩება გამიჯნული დანარჩენი პოპულაციებისაგან, მაშინ მათ შორის გენეტიკური ინფორმაციის მიმოცვლა წყდება. გენოფონდის ხანგრძლივი და ვექტორიზებული ცვლილებით შესაძლებელია მოხდეს სახეობათა დიფერენცირება, რასაც, ბუნებრივია, თან სდევს რეპროდუქციული იზოლაციის ჩამოყალიბება. გამიჯნული ფორმების განმეორებითი შეხვედრისას საიზოლაციო მექანიზმების მოშლის გამო ისინი ერთმანეთს აღარ ეჯვარებიან, რადგან გენეტიკურ დამოუკიდებლობას ფლობენ.
სახეობის შემადგენელ ნაწილთა შორის წარმოქმნილი რეპროდუქციული იზოლაციით ერთი საწყისი სახეობის ორად გაყოფა არის სახეობათა წარმოშობის პროცესი. სქემატურად ეს პროცესი მოცემულია 12.1 სურათზე. ამრიგად სახეობათა წარმოქმნა არის საწყისი სახეობის დროსა და სივრცეში ორ ან მეტ სახეობად ჩამოყალიბება. სხვაგვარად თუ ვიტყვით, სახეობათა წარმოშობა არის გენეტიკურად ღია სისტემის (ამდაგვარია სახეობის შემადგენელი პოპულაციები და სხვა ზეპოპულაციური რანგები) დახშულ სისტემად გარდაქმნის პროცესი. სახეობათა ჩამოყალიბება სახეობის შიგნით მიმდინარე მიკროევოლუციის გზით ხორციელდება.
მუტაციური პროცესისა და კომბინაციური ცვალებადობის საფუძველზე პანმიქსიით სახეობაში შენარჩუნებულია ჰეტეროზიგოტულობა და ჰეტეროგენულობის მაღალი დონე. ცვალებადობის ამ „სამობილიზაციო რეზერვით“ იქმნება მრავალფეროვანი (გენეტიკური კომბინაციები) მასალა ბუნებრივი გადარჩევისათვის. ამასთანავე, კონკრეტული სახეობის შიგნით პანმიქსიით ხორციელდება განსხვავებათა ნიველირება, რაც აბრკოლებს შედარებით დიდი დაჯგუფებების (პოპულაციები და მათი ჯგუფები) ევოლუციურ დიფერენცირებას. სახეობის გენეტიკური სტრუქტურა (გენოფონდი) საიზოლაციო მექანიზმებით დაცულია უცხო (სხვა სახეობის) გენთა ნაკადებისაგან. სახეობის შიგნით მიმდინარე გენთა ნაკადის მეშვეობით კი გენოფონდი ერთიანი ხდება. ეს პროცესები არსებით როლს ასრულებენ სახეობის მდგრადობის შენარჩუნებაში. ამრიგად, ნებისმიერი სახეობა მისთვის ნიშანდობლივ უნიკალობასა და ადაპტურ სპეციფიკურობას ინარჩუნებს. ზემოთ აღნიშნულის შედეგად, ჩვენს პლანეტაზე არა მარტო შენარჩუნებულია, არამედ თანდათანობით იზრდება სახეობათა მრავალფეროვნება.
|
|
14.3 12.3. სახეობათა წარმოქმნის პროცესი და გზები |
▲ზევით დაბრუნება |
ევოლუციური ბიოლოგიის ერთ-ერთ ცენტრალურ პრობლემას სახეობა და სახეობათა წარმოქმნის პროცესი წარმოადგენს. ბიოლოგიის ვერცერთმა დარგმა ვერ შეძლო ამ პრობლემის გადაჭრა. ჩ.დარვინი თავის ძირითად ნაშრომში „სახეობათა წარმოშობა...“ (1859) სახეობათა წარმოქმნასა და ევოლუციას აიგივებდა. დარვინის ძირითადი მიზანი იყო ევოლუციის პროცესის ჩვენება, რასაც ჯ. რომანესმა (1897) უწოდა „სახეობათა გარდაქმნა დროში“, ხოლო ჯ. სიმპსონმა (1944) „ფილეტური ევოლუცია“. ე. მაირის (1968) აზრით, ჭეშმარიტ სახეობათა წარმოქმნის დროს სახეობათა რაოდენობა იზრდება, რასაც რომანესმა უწოდა, „სახეობათა გამრავლება სივრცეში“. ამ დროს საწყისი ერთგვაროვანი სახეობიდან რამდენიმე „შვილეული“ სახეობა ყალიბდება. ბუნებაში ეს პროცესი სახეობათა წარმოქმნის გზით მიმდინარეობს. ამ პრობლემის გადაჭრაში დიდი წვლილი მიუძღვის ჰარვარდის (აშშ) უნივერსიტეტის პროფესორს ე.მაირს, მაგრამ არაერთი საკითხი კვლავ გადაუწყვეტელია. ამ პრობლემისადმი დიდი ინტერესი იმით აიხსნება, რომ სახეობათა წარმოქმნის პროცესი ევოლუციის ერთ-ერთი უმნიშვნელოვანესი ეტაპია.
სახეობათა წარმოქმნის გზები. ბოლო ათწლეულებში ბიოლოგიურ ევოლუციაში მიღწეული წარმატებები სახეობათა წარმოქმნის ძირითადი გზების კლასიფიცირების საშუალებას იძლევა. ჰექსილის (1942) მოსაზრებით, ცოცხალ ბუნებაში სახეობათა წარმოქმნა სამი გზით (გეოგრაფიული, ეკოლოგიური და გენეტიკური) ხორციელდება. ამ კლასიფიკაციის კრიტერიუმი კარგად ვერაა შერჩეული, რადგან სხვადასხვა კატეგორიები ერთიმეორეს ფარავენ. ასე მაგალითად, გეოგრაფიულ სახეობათა წარმოქმნის ნებისმიერი ფორმა იმავდროულად ეკოლოგიურსა და გენეტიკურსაც მოიცავს, ადრე ე. მაირმა (1942) ადრე სახეობათა წარმოქმნის ოთხი გზა გამოყო, ხოლო ს. რაიტმა (1949) ათი ურთიერთგამომრიცხავი შესაძლო გზა აღწერა: კლასიფიკაციას ორი კრიტერიუმი დაედო საფუძვლად, კერძოდ: 1. ახლად წარმოქმნილ სახეობას რამდენმა საწყისმა სახეობამ დაუდო სათავე. 2. საწყისი პოპულაციის ზომა რომლიდანაც ჩამოყალიბდა ახალი სახეობა.
ცხრილი 11.1.
სახეობათა წარმოქმნის კლასიფიკაცია
|
სახეობათ წარმოქმნის |
I. |
სახეობათა გარდაქმნა
1. აუტოგენური ტრანსფორმაცია
2. ალოგენური ტრანსფორმაცია |
II. |
სახეობათა შემცირება |
III. |
სახეობათა რიცხვის გაზრდა
1. სახეობების მყისიერი
ა. ქრომოსომული გარდაქმნები
2. თანდათანობითი (გრადუალური)
ა. სიმპატრიული |
ე. მაირმა წარმოდგენილი სქემების ურთიერთშეჯერებით გამოყო სახეობათა წარმოქმნის 12 შესაძლო გზა. კლასიფიკაცია სამ ძირითად კრიტერიუმს ეფუძნება: 1. ახალი რეპროდუქციული იზოლაციის ჩამოყალიბების უნარი; 2. სახეობათა წარმოქმნის მექანიზმის (გრადუალური ან მყისეული) ფორმა; 3. საწყისი პოპულაციის გეოგრაფიულ იზოლაციასთან (მოქმედებდა თუ არა) დამოკიდებულება. სახეობის პრობლემის აღიარებული მკვლევრის ე.მაირისა და სხვა თანამედროვე ავტორთა მონაცემების გათვალისწინებით 12.1 ცხრილში მოტანილია სახეობათა წარმოქმნის შედარებით გამარტივებული კომბინირებული კლასიფიკაცია.
სახეობათა წარმოშობას განიხილავენ ტერიტორიულ და ფილოგენეტიკურ ასპექტში. შეიძლება ახალი სახეობა ფორმირდეს საწყისი სახეობის არეალის პერიფერიაზე პოპულაციის ან პოპულაციათა მცირე ჯგუფისაგან. იმის გამო, რომ სახეობათა წარმოქმნა სხვადასხვა ტერიტორიაზე მიმდინარეობს, ამ გზას გეოგრაფიული ანუ ალოპატრიული (ბერძნ. ალლოს - სხვა+პატრის სამშობლო) უწოდეს. როდესაც ახალი სახეობა ყალიბდება საწყისი სახეობის არეალში, ასეთ სახეობათა წარმოქმნის გზას სიმპატრიული (SYN-ერთად+პატრის სამშობლო) ეწოდება.
ახალი სახეობა შეიძლება ჩამოყალიბდეს არა მარტო სივრცეში, არამედ დროშიც. როდესაც ახალი სახეობის ფორმირება ხანგრძლივი დროის განმავლობაში თანდათანობით დივერგირების გარეშე მიმდინარეობს, ამ გზას ფილეტური ეწოდება. ახალი სახეობა შეიძლება ჩამოყალიბდეს ხანგრძლივი დროის განმავლობაში საწყისი სახეობის დივერგირებით და ამით რამდენიმე ახალ სახეობას ეძლევა დასაბამი. ამ გზას დივერგენტული ეწოდება ზოგჯერ ახალი სახეობა წარმოიქმნა ორი ადრე ჩამოყალიბებული სახეობის ჰიბრიდიზაციით. სახეობათა ჩამოყალიბების ასეთ გზას ჰიბრიდოგენული ეწოდება.
ფილეტურ და ჰიბრიდოგენულ სახეობათა წარმოქმნისას სახეობათა რაოდენობა არ მატულობს და საიზოლაციო მექანიზმები არ ევოლუირებს. ამასთანავე, საწყისი სახეობის თვითმყოფადობა იკარგება. ალოპატრიულ, კვანტურ და სიმპატრიულ სახეობათა წარმოშობისას საწყისი (დედისეული) სახეობიდან ერთი ან რამდენიმე სახეობას ეძლევა დასაბამი. ამასთანავე საწყისი (ძველი) სახეობა, წესისამებრ, თვითმყოფობას ინარჩუნებს. ამიტომაც ამ ტიპის სახეობათა წარმოქმნას სახეობის რაოდენობის მატებას ანუ ჭეშმარიტ სახეობათა წარმოქმნას უწოდებენ.
სახეობათა წარმოქმნის პროცესი. სახეობები არის ერთიმეორისგან რეპროდუქციულად იზოლირებული ჯგუფები. თუ გვინდა დავადგინოთ, როგორ მიმდინარეობს სახეობათა წარმოქმნის პროცესი, უპირველეს ყოვლისა უნდა შევისწავლოთ, როგორ ყალიბდება რეპროდუქციული იზოლაცია. თავდაპირველად რეპროდუქციული იზოლაცია წარმოიქმნება როგორც გენეტიკური დივერგენციის გვერდითი შედეგი. მისი ჩამოყალიბება, სრულქმნა და დახვეწა ბუნებრივი გადარჩევის გზით ხორციელდება. გამოვლენილი და შესწავლილია სახეობათა წარმოქმნის მრავალი გზა, რომელიც სხვადასხვა საშუალებებით ხორციელდება; მიუხედავად ამისა, მათში შეიძლება გამოიყოს ორი ძირითადი სტადია (იხ. სურ. 12.2).
I სტადია. სახეობათა წარმოქმნის საწყის ეტაპზე კონკრეტულ სახეობის ორ პოპულაციას შორის გენთა ნაკადი წყდება. ე.ი. გენეტიკური ინფორმაციის მიმოცვლა აღარ მიმდინარეობს. იმის გამო, რომ პოპულაციების საარსებო გარემო რამდენადმე განსხვავებულია, თითოეული მათგანი კონკრეტულ გარემოსთან, ან განსხვავებულ ცხოვრების ნირთან გენეტიკურად სპეციფიკურად დიფერენცირდება. გენეტიკური დიფერენცირების პროცესში გარკვეულ როლს ასრულებს გენთა დრეიფი. პოპულაციათა შორის გენთა ნაკადის შეწყვეტა ახალ სახეობათა ფორმირების ერთ-ერთი მნიშვნელოვანი წინაპირობაა, ვინაიდან წინააღმდეგ შემთხვევაში მათ ერთიანი გენოფონდი ექნებათ, რის გამოც მათი გენეტიკურად დიფერენცირება შეუძლებელია. პოპულაციათა შორის გენეტიკური განსხვავების აკუმილაციასთან ერთად, დიფერენცირებასა და ამოქმედებას იწყებს რეპროდუქციული საიზოლაციო მექანიზმები. თუ პოპულაციის გენოფონდი არ არის კოადაპტირებული ჰიბრიდულ ინდივიდებში ცხოველმყოფელობა და ნაყოფიერება შესაბამისად დაქვეითებულია.
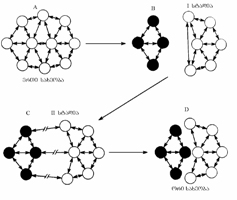
სურ. 12.2. სახეობათა წარმოქმნის ზოგადი მოდელი. A. სახეობის შემადგენელი პოპულაციები მოცემულია რგოლებით ისრით - გენთა ნაკადი. B. პოპულაციები ორ ჯგუფად იზოლირდება და დამოუკიდებლად დიფერენცირდება. ჩ. რეპროდუქციული საიზოლაციო მექანიზმების ჩამოყალიბების გამო, გენთა ნაკადი შემცირებულია, რაც გაწყვეტილი ისრით არის გამოსახული. ისინი სახეობათა წარმოშობის მეორე სტადიაზე იმყოფებიან. D. ორივე ჯგუფი რეპროდუქციულად იზოლირებულია და გენთა ნაკადი სრულადაა შეწყვეტილი. სახეობათა წარმოშობა დასრულებულია (ფ. აიალა, 1984).
ამრიგად, სახეობათა წარმოქმნის პირველ სტადიას მოეპოვება შემდეგი ნიშნები: 1. რეპროდუქციული იზოლაცია თავდაპირველად პოსტზიგოტური საიზოლაციო მექანიზმებით წარმოიქმნება; 2. ჩამოყალიბებული საიზოლაციო მექანიზმი გენეტიკური დიფერენცირების გვერდითი (დამატებითი) რეზულტატია. ამ ეტაპზე რეპროდუქციული იზოლაციის ჩამოყალიბებაში ბუნებრივი გადარჩევა უშუალოდ არ მონაწილეობს.
გენეტიკური დიფერენცირება და მასთან თანხვედრილი პოსტზიგოტური საიზოლაციო მექანიზმები თანდათანობით ყალიბდება. თუ ორ კონკრეტულ პოპულაციას პოსტზიგოტური იზოლაცია მიჯნავს, მაშინ ისინი სახეობათა წარმოქმნის პირველ სტადიაზე იმყოფებიან.
II სტადია. ამ სტადიაზე მთავრდება რეპროდუქციული იზოლაციის ჩამოყალიბება. დავუშვათ, რომ მოიშალა საიზოლაციო მექანიზმები, რომლებიც I სტადიაზე პოპულაციებს შორის გენთა ნაკადს აბრკოლებდა. მაგალითად, ეს შეიძლება მოხდეს, როდესაც ტერიტორიულად დაცილებულ ორივე პოპულაციაში იზრდება რიცხოვნობა და ინდივიდთა ერთი ნაწილი ახალ ტერიტორიას იკავებს. თუ ორივე პოპულაციამ ერთი და იგივე ტერიტორია დაიკავა, მაშინ შეინიშნება ორი შედეგი: 1. ყალიბდება ერთიანი გენოფონდი; თუ ჰიბრიდებში ცხოველქმედება უმნიშვნელოდ არის დაქვეითებული, მაშინ არ ბრკოლდება პოპულაციათა აღრევა (პანმიქსია); 2. წარმოიქმნება ორი დამოუკიდებელი სახეობა; რადგან ბუნებრივი გადარჩევის მოქმედებით საიზოლაციო მექანიზმების განმტკიცება და სრულქმნა ხორციელდება.
სახეობათა წარმოქმნის პირველი სტადია შექცევადია: თუ გენეტიკური დიფერენცირების პროცესი ღრმად და შორს არ არის წასული, მაშინ ორ პოპულაციას შეუძლია შერწყმა და ერთიანი გენოფონდის ჩამოყალიბება. მათ შორის არსებული განსხვავება ნიველირდება. იმ შემთხვევაში თუ პოპულაციათა შორის ჰიბრიდებში ცხოველმყოფელობა და ნაყოფიერება მნიშვნელოვნადაა დაქვეითებული, მაშინ ბუნებრივი გადარჩევის მოქმედება ხელს უწყობს შიგაპოპულაციურ ჰიბრიდიზაციას (ბუნებრივია მნიშვნელოვნად ბრკოლდება პოპულაციათაშორისი შეჯვარებები). სამაგალითოდ განვიხილოთ გამარტივებული მოდელი. დავუშვათ, რომელიმე ლოკუსი ორი A1 და A2 ალელითაა წარმოდგენილი. A1 ალელი ხელს უწყობს შიგაპოპულაციურ ჰიბრიდიზაციას, A2 - პოპულაციათაშორის ჰიბრიდიზაციას. A1 ალელი მაღალი სიხშირით ექმნებათ შიგაპოპულაციის ჰიბრიდიზაციით მიღებულ შთამომავლებს, ე.ი. მაღალი ცხოველმყოფელობისა და ნაყოფიერების ინდივიდებს; ხოლო A2 ალელი გაცილებით მაღალი სიხშირით პოპულაციათაშორისი შეჯვარებით მიღებულ ჰიბრიდებს მოეპოვებათ. ვინაიდან ამ უკანასკნელში ადაპტირების უნარი დაქვეითებულია, მათი ხვედრითი წილი მომდევნო თაობებში თანდათანობით შემცირდება. ბუნებრივი გადარჩევა ხელს უწყობს A1 ალელის წილის გაზრდას (ვინაიდან გაცილებით კონკურენტუნარიანია და აპირობებს შიგაპოპულაციურ შეჯვარებას) და ახდენს A2 ალელის მქონე ჰიბრიდთა ელიმინაციას. ყოველივე ეს მიგვანიშნებს, რომ ბუნებრივი გადარჩევა მოქმედებს პრეზიგოტური რეპროდუქციული საიზოლაციო მექანიზმების ჩამოყალიბების სასარგებლოდ და თრგუნავს პოპულაციათაშორისი ჰიბრიდული ზიგოტების წარმოქმნას.
სახეობათა წარმოქმნის მეორე სტადია ხასიათდება შემდეგი ნიშნებით: 1. რეპროდუქციული იზოლაცია ვითარდება ძირითადად პრეზიგოტური საიზოლაციო მექანიზმების სახით; 2. პრეზიგოტური საიზოლაციო მექანიზმების ჩამოყალიბება ბუნებრივი გადარჩევის მოქმედებით ყალიბდება. ეს ორი ნიშანი ამ სტადიას პრინციპულად ასხვავებს პირველი სტადიისაგან. უნდა აღინიშნოს, რომ სახეობათა წარმოქმნა ზოგჯერ შეიძლება მეორე სტადიის გარეშეც წარიმართოს. პოპულაციათაშორის გენთა ნაკადის შეწყვეტისას, შესაძლებელია სრული იზოლაცია ჩამოყალიბდეს (თუ გენეტიკური დიფერენცირება ხანგრძლივი დროის განმავლობაში მიმდინარეობს). ასე მაგალითად, იზოლირებულ ოკეანურ კუნძულებზე შემთხვევით მოხვედრილი ინდივიდები ქმნიან პოპულაციას, რომლის გენეტიკური დიფერენცირება დიდ პერიოდს მოიცავს. მათ უყალიბდებათ საიზოლაციო მექანიზმები და დამოუკიდებელი ენდემური სახეობის სტატუსს იძენენ. იმის გამო, რომ ბუნებრივი გადარჩევა უშუალოდ მონაწილეობს რეპროდუქციული იზოლაციის მექანიზმების ჩამოყალიბებაში მეორე სტადია მნიშვნელოვნად აჩქარებს სახეობათა ჩამოყალიბების პროცესს.
|
|
14.4 12.4. სახეობათა ალოპატრიული წარმოქმნა |
▲ზევით დაბრუნება |
ჭეშმარიტი სახეობათა წარმოქმნის პროცესის ასახსნელად შემოთავაზებული ჰიპოთეზებიდან ყველაზე ფართო აღიარება მოიპოვა ალოპატრიულმა (გეოგრაფიულმა) თეორიამ, რომელიც ფორმულირებულია გამოჩენილი გერმანელი ევოლუციონისტის მორიც ვაგნერის (1868) მიერ.
სახეობათა ალოპატრიული წარმოქმნა ეწოდება სივრცობრივად იზოლირებულ პოპულაციათა დიფერენცირებას, რაც ისე შორს მიდის, რომ შემთხვევითი კონტაქტის დროს თავისუფალი შეჯვარება (პანმიქსია) შეუძლებელია. იგი არის სახეობათა წარმოქმნის ყველაზე ბუნებრივი და გავრცელებული გზა. ალოპატრიულ სახეობათა წარმოქმნა შეიძლება ოთხ ეტაპად დაიყოს: 1. იზოლაცია - საწყისი გენოფონდის დაყოფა ორ ან ორზე მეტ იზოლირებულ ჯგუფად. 2. დიფერენცირება - ორი ან მეტი გენოფონდის დამოუკიდებლად ევოლუირება; 3. მეორადი შერწყმა. 4. ახალ გენოფონდებს შორის კონკურენცია.
იზოლაცია. ფიზიკური გარემოს (ან ანთროპული ფაქტორის) ზემოქმედებით პოპულაცია ორ ან ორზე მეტ იზოლირებულ ჯგუფად იყოფა. იგი იწვევს პოპულაციის მეტ-ნაკლებად გამიჯნული ნაწილების სრულ ან ნაწილობრივ იზოლაციას. ბუნებრივია, მათ შორის გენთა ნაკადი სუსტდება ან სრულიად წყდება. ვინაიდან იზოლირებულ ჯგუფებს განსხვავებულ საარსებო პირობებში უხდებათ არსებობა და ადაპტირება, ისინი განსხვავებულ გენეტიკურ სტრუქტურას იძენენ. ე.ი. მათი დიფერენცირება სხვადასხვა მიმართულებით მიმდინარეობს. დიფერენცირება ეწოდება პროცესს, რომლის მეშვეობით ორი ან ორზე მეტი ერთიმეორისგან იზოლირებული პოპულაცია განსხვავებული მიმართულებით ევოლუირებს. საწყის ეტაპზე მადიფერენცირებელი ფაქტორებია: მუტაციური პროცესი, რეკომბინაცია და ბუნებრივი გადარჩევა. დიფერენცირების პროცესში ბუნებრივი გადარჩევის გარდა მნიშვნელოვან როლს ასრულებს ევოლუციის სხვა ელემენტარული ფაქტორებიც (განსაკუთრებით გენთა დრეიფი, ინბრიდინგი).
არეალის შერწყმის გამომწვევი ფაქტორები მსგავსია იმ ფაქტორებისა, რამაც საწყისი ერთიანი პოპულაციის დანაწევრება გამოიწვია. მიმდინარეობს I ეტაპის პირუკუ პროცესი - საიზოლაციო მექანიზმების მოშლა. ძირითადად ეს გარემოს ცვლილებაა, რომელიც სახეობის არეალის გაფართოებას იწვევს. ზოგჯერ ეს გამოწვეულია ისეთი მიგრაციით, როდესაც ორი განსხვავებული პოპულაცია ერთსა და იმავე ახალ ტერიტორიას ითვისებს. არეალის ურთიერთგადაფარვის გამო, განსხვავებული პოპულაციის წევრები იმდენად უახლოვდებიან ერთმანეთს, რომ შესაძლებელია ჰიბრიდიზაცია. მყარი სარეპროდუქციო იზოლაციის ფორმირების შემთხვევაში განსხვავებული პოპულაციის წევრები არ ეჯვარებიან ერთიმეორეს. შეჯვარების შემთხვევაში მიღებული ზიგოტები სიცოცხლის უუნაროა და მათგან შთამომავლობა არ მიიღება. ამ შემთხვევაში სახეობათა წარმოქმნის პროცესი დასრულებულია.
მაგრამ, თუ ახლად ფორმირებული განსხვავებული პოპულაციის ინდივიდები მსგავს ეკოლოგიურ მოთხოვნას ფლობენ და ერთიმეორეს ეჯვარებიან, მაშინ ისინი ურთიერთქმედებენ ერთიმეორეზე. ამ შემთხვევაში საბოლოო შედეგი ჩამონათვალიდან ერთ-ერთით მთავრდება:
1. ერთი სახეობა აძევებს დაკავებული ტერიტორიიდან მეორეს და სახეობის არეალი გადაუფარავი ხდება.
2. ბუნებრივი გადარჩევის მოქმედებით ორი ახლად ჩამოყალიბებული სახეობა იყოფს ტერიტორიას, მათ შორის ესა თუ ის დაბრკოლება წარმოიქმნება. სახეობათაშორისი ჰიბრიდიზაცია წყდება ან დაბალი სიხშირით მიმდინარეობს.
3. სახეობათშორისი ჰიბრიდიზაციით ფერტილური შთამომავლობა მიიღება (იხ. სახეობათა ჰიბრიდოგენური წარმოქმნა).
ორი იზოლირებული მონათესავე (საწყისი სახეობიდან ახლადფორმირებული) სახეობის განმეორებითი შეხვედრისას, ისინი კონკურენციის სხვადასხვა ფორმით ურთიერთქმედებენ (ეს საკითხი ვრცლადაა განხილული სახეობათაშორის ბრძოლაში (იხ. §8.3) და მასზე აღარ შევჩერდებით).
სურ. 12.3. სახეობათა წარმოშობა ავსტრალიურ თუთიყუშში (გვარი Pachycephala) სახეობების ამჟამინდელი გავრცელება მოცემულია მე-6 რუკაზე (ე. მაირი, 1968).
ალოპატრიული სახეობის ჩამოყალიბების ერთ-ერთი ხერხი დაკავშირებულია არეალის იზოლირებულ ნაწილებად დაყოფასთან. საწყისი, ფართო გავრცელების სახეობის ფრაგმენტაციის შედეგად შეიძლება ორი ან მეტი ახალი სახეობა ჩამოყალიბდეს. ამ გზით წარმოიშვა შროშანას გვარში (Convallaria) გაერთიანებული სახეობები. საწყისი სახეობა გავრცელებული იყო ევრაზიის ფოთლოვანი ტყეების დიდ არეალზე. მეოთხეულ პერიოდში გამყინვარების შედეგად არეალი ნაწილებად დაიყო. მცენარე შემორჩა ამიერკავკასიის, სამხრეთ ევროპისა და შორეული აღმოსავლეთის ტყიან ზოლში. ცალკეულ ტერიტორიაზე გადარჩენილ ფორმათაგან სამი დამოკიდებული სახეობა ჩამოყალიბდა, ევროპაში C. majalis, ამიერკავკასიაში C. transcavcasica, შორეულ აღმოსავლეთში კი C. keiskii.
გეოგრაფიული იზოლატი ალოპატრიულ სახეობათა წარმოქმნის კერაა. ჩვეულებრივ ასეთი ტერიტორია ერთ სახეობას უკავია. იშვიათად გვხვდება ორი მონათესავე სახეობა, რაც არეალის იზოლირებულ ნაწილებად დაყოფასა და შემდგომ სახეობის განმეორებით განსახლებითაა გამოწვეული. სანიმუშოდ მოვიყვანთ განმეორებითი განსახლების კლასიკურ მაგალითს. ამჟამად ავსტრალიაში გავრცელებულია თუთიყუშის ორი ახლომონათესავე სახეობა (Pachycephala rufogularis და P. inornata). პალეოგეოგრაფიული და პალეოკლიმატოლოგიის მონაცემების გათვალისწინებით შეძლეს სახეობათა ჩამოყალიბების ისტორიის დადგენა და, ერთი შეხედვით, მეტად უცნაურ სიტუაციაში გარკვევა. კერძოდ, ერთი სახეობის მიერ დაკავებულ ტერიტორიაზე მეორე მონათესავე სახეობა საკმაოდ ვიწრო არეალზეა განსახლებული. (სურ. 12.3 რუკა 6). თავდაპირველად საწყის სახეობას ფართო არეალი ეკავა (სურ. 12.3-1). მშრალი პერიოდის დადგომამ და მცენარეული საფარის (შერეული წვიმის ტყე). გარკვეულ ტერიტორიაზე შეცვლამ საწყისი სახეობის აღმოსავლეთისა და დასავლეთის ნაწილები ერთიმეორეს მშრალი ზონის ფართო კორიდორით გამიჯნა. მეტ-ნაკლებად განსხვავებულ საარსებო პირობებში მიკროევოლუციური პროცესების შედეგად თითოეული მათგანი დამოუკიდებელ სახეობად დიფერენცირდა (სურ. 12.3-3,4). მომდევნო პერიოდში კლიმატი კვლავ შეიცვალა და ტერიტორიაზე ტენის მოყვარული მცენარეები დასახლდა. დასავლეთში ჩამოყალიბებულმა თუთიყუშის სახეობამ შეძლო არეალის განვრცობა და აღმოსავლეთით საკმაოდ დიდი ტერიტორიის ათვისება (სურ. 12.3-5). განმეორებითი განსახლების შედეგად ორი მონათესავე თუთიყუშის სახეობა კვლავ შეხვდა ერთიმეორეს (მეორადი შერწყმა). ორივე სახეობა გენეტიკურ დამოუკიდებლობას ფლობს. მათ აქვთ დიფერენცირებული გენოფონდი. რადგანაც თუთიყუშის ორივე სახეობაში მდგრადი რეპროდუქციული იზოლაციაა ჩამოყალიბებული, ისინი ერთიმეორეს აღარ ეჯვარებიან. კლიმატის განმეორებით შეცვლამ და შესაბამისად მცენარეულ საფარში მომხდარმა ცვლილებებმა დასავლეთის სახეობა ორ ნაწილად გაყო. აღმოსვლეთ ავსტრალიაში გავრცელებულ თუთიყუშში ამდაგვარი პერეტრუბაციები არ მომხდარა, მას სტაბილურად საწყისი არეალი უკავია.
სურ. 12.4. დიდი თოლიების ქვესახეობის უწყვეტი რგოლები; ვერცხლისფერი თოლია (Larus argentatus) და დიდთოლია (L. fuscus). გამოყოფილია ცალკეული ქვესახეობის არეალი. ჩრდილო-დასავლეთ ევროპაში ჯაჭვის კიდურა რგოლები (ორმაგი შტრიხი) ერთმანეთს ხვდება. ბუნებაში ისინი არ ეჯვარებიან ისრებით ნაჩვენებია განსახლების ისტორიული გზა (ე. მაირი, 1968).
სახეობის ჩამოყალიბების მეორე გზა დაკავშირებულია საწყისი სახეობის არეალის გაფართოებასთან. არეალის გაფართოების დროს სახეობის პოპულაციები ახალ საარსებო გარემოში აღმოჩნდებიან. შემდგომმა მიკროევოლუციურმა პროცესმა შეიძლება ახალი სახეობის წარმოქმნა გამოიწვიოს. ბალტიისა და ჩრდილოეთის ზღვების სანაპიროზე გავრცელებულია თოლიას ორი სახეობა: ვერცხლისფერი თოლია (Larus argentatus) და დიდთოლია (L. fuscus) (სურ. 12.4). ეს ორი სახეობა ერთიმეორეს უკავშირდება ქვესახეობით წარმოქმნილი უწყვეტი რგოლებით. ისინი გვხვდება ერთი მხრივ, ჩრდილო ევრაზიაში, მეორე მხრივ - გრენლანდიასა და ჩრდილოეთ ამერიკაში. ზემოთ დასახელებული სახეობების წარმოშობის ისტორია შემდეგია: ამ თოლიების წინაპარი რამდენიმე ათასი წლის წინათ ამჟამინდელ ბერინგის სრუტესთან ბინადრობდა. იგი განსახლდა აღმოსავლეთით და დასავლეთით. ჩამოყალიბდა ქვესახეობისაგან შემდგარი ორი უწყვეტი ჯაჭვი. ყველა მეზობელ ქვესახეობაში გაერთიანებული ინდივიდები ერთიმეორეს ეჯვარებიან და ფერტილურ შთამომავლობას იძლევიან. ბალტიისა და ჩრდილოეთის ზღვის რაიონში აღმოსავლეთისა და დასავლეთის ჯაჭვის კიდურა ქვესახეობის რგოლები ერთიმეორეს შეხვდნენ. მიკროევოლუციის პროცესში ცალკეულ ქვესახეობებში ბიოლოგიურ განსხვავებათა დაგროვება საკმარისი აღმოჩნდა ორი დამოუკიდებელი სახეობის ჩამოსაყალიბებლად. თუ ორი სახეობის დამაკავშირებელი რომელიმე რგოლი გაწყდა, მაშინ ორი, სრულიად დამოუკიდებელი, სახეობა ჩამოყალიბდება.
სახეობათა ალოპატრიულ წარმოქმნას სივრცობრივი იზოლაციის ესა თუ ის ფორმა უძევს საფუძვლად. იგი შედარებით ნელა, ასეულ ათას თაობათა პოპულაციებში მიმდინარეობს. დროის ასეთ დიდ შუალედში იზოლირებულ პოპულაციათა შორის ისეთი ბიოლოგიური ცვლილებები ყალიბდება, რომელიც მათ რეპროდუქციულ იზოლაციას იწვევს. სახეობათა ალოპატრიული წარმოქმნა ყოველთვის სახეობრივი არეალის ფორმირებასთან არის დაკავშირებული.
|
|
14.5 12.5. სახეობათა კვანტური წარმოქმნა |
▲ზევით დაბრუნება |
სახეობათა კვანტური წარმოქმნა არის საწყისი სახეობის იზოლირებული „კუნძულებრივი“ (ფართო მნიშვნელობით) ან პერიფერიული პოპულაციიდან ახალი სახეობრივი ფორმების ჩამოყალიბება. სახეობათა წარმოშობის ეს ხერხი არ ეწინააღმდეგება ევოლუციის სინთეზური თეორიის ძირითად დებულებებს. ვინაიდან იგულისხმება პოპულაციის დონეზე სივრცობრივი იზოლაციის მონაწილეობით სახეობის ფორმირება. სახეობათა წარმოშობის კლასიკური შემთხვევისაგან (პოპულაცია Ⴎ პოპულაციათა ჯგუფი Ⴎ გეოგრაფიული რასა Ⴎ ალოპატრიული ქვესახეობა) განსხვავებით ახალი სახეობრივი ფორმა შუალედური რგოლების გარეშე ყალიბდება. მაშასადამე, ეს გზა სახეობათა ფორმირების შემოკლებული ხერხია. ტერმინი შემოთავაზებულია ვ. გრანტის მიერ, კვანტურ სახეობათა წარმოქმნის იდეა კი ჯ. სიმპსონის და ე. მაირის კონცეფციებს ეფუძნება (მაირი იყენებს ტერმინს „გენეტიკური რევოლუცია“).
ჯ. სიმპსონის შეხედულებით, კვანტური რევოლუციის დროს, პოპულაცია ერთი ადაპტური პიკიდან მეორეზე მყისიერად გადადის. ბუნებრივია, მასში ადრე ჩამოყალიბებული ადაპტური წონასწორობა ირღვევა, რის გამოც პოპულაციათა ელიმინაციის ალბათობა საკმაოდ მაღალია. ამდაგვარ პოპულაციაში ღრმა ადაპტური ცვლილებები მიმდინარეობს. გენოფონდში აკუმულირებულია მრავალი მცირე მუტაცია, რომელთაგანაც ნაწილი საზიანოა.
ფორმირებადი პოპულაცია, ფლუქტუირებად ახალ საარსებო გარემოსადმი ან გენოფონდში არაადაპტური მუტაციის ფიქსაციის გამო, არამდგრადია, ამიტომაც კვანტური ევოლუციის გზით მრავალი პოპულაციიდან სახეობად მხოლოდ რამდენიმე თუ ყალიბდება. კვანტური ევოლუციით სახეობა წარმატებულად მხოლოდ იმ შემთხვევაში ფორმირდება, თუ პოპულაცია ახალ საარსებო საშუალებისადმი ადაპტურ ცვლილებებს ფლობს.
წესისამებრ კვანტურ სახეობათა წარმოქმნა იწყება ფართო არეალის, მაღალი პოლიმორფული სტრუქტურის პოპულაციებში. მაირის შეხედულებით, კვანტურ სახეობათწარმოშობაში არსებით როლს ფართო არეალის პერიფერიულად (უკიდურესი საზღვრისპირა უბნები) განთავსებული იზოლატები ასრულებენ (იხ. § 11.4). ამ სახის იზოლატებს, ჩვეულებრივ, საფუძველს უყრის საწყისი პოპულაციიდან მიგრირებულ ინდივიდთა ძალზე მცირე ჯგუფი (დამფუძნებლის პრინციპი). დამფუძნებელი ინდივიდებით ყალიბდება პირველადი პოპულაცია. იმის გამო, რომ მცირერიცხოვანი ჯგუფი (ზოგჯერ ინდივიდი) შემთხვევითი ამორჩევით ხვდება ახალ საარსებო გარემოში, მათგან ფორმირებული კოლონია გენეტიკური სტრუქტურით (ალელთა სიხშირით) ატიპურია და საკმაოდ განსხვავდება საწყისი პოპულაციისაგან. პოპულაციის საწყის ეტაპზე კოლონიის წევრები ურთიერთშეჯვარებით მრავლდებიან (ასორტული შეჯვარება). მათ აღენიშნებათ იძულებითი ინბრიდინგი. მოქმედებას იწყებს გენთა დრეიფი. ინბრიდინგს თან სდევს ინსერციული მუტაგენეზი, რაც მობილურ დისპერგირებულ გენეტიკური ელემენტების გადაადგილებით არის გამოწვეული. სწრაფად წარმოიქმნება მემკვიდრული ცვალებადობის ახალი „სამობილიზაციო რეზერვი“ ანუ მიმდინარეობს ახალი მემკვიდრული ცვლილებების დაგროვება და განახლება (იხ. §5.3; 7.2 და 7.4) ბუნებრივი გადარჩევის მოქმედებით სელექციურად მნიშვნელოვანი ღირებულების ცვლილებები გროვდება და ვრცელდება პოპულაციაში, რის შედეგადაც მას უყალიბდება სრულიად განსხვავებული ბალანსირებული გენეტიკური სტრუქტურა - გენოფონდი.
ე. მაირის შეხედულებით, ჭეშმარიტი გენეტიკური რევოლუციისათვის, მემკვიდრული მასალის გარდაქმნის ან ნაწილის დაკარგვის გამო, ნიშანდობლივია გენეტიკური ჰომეოსტაზის დარღვევა. თუ პოპულაცია ამდაგვარ რევოლუციას გადაურჩა (მათი რაოდენობა ძალზე მცირეა) ახალ ჰომეოსტაზს იყალიბებს.
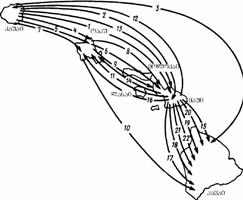
სურ. 12.5. ჰავაის კუნძულებზე ბინადარი დროზოფილების ხუთი ჯგუფის მიერ ჰავაის კუნძულების მიმართული კოლონიზაცია (ციფრებით აღნიშნულია განმეორებითი კოლონიზაციის რაოდენობა) (ლ. კაიდანოვი, 1996).
კვანტური ევოლუციის გზით არის ჩამოყალიბებული ჰავაის არქიპელაგზე გავრცელებული დროზოფილას გვარში გაერთიანებული ზოგიერთი სახეობა (იხ. სურ. 12.5). სურათზე მოცემულია დროზოფილას გავრცელების გზები. ჰავაის კუნძულებზე გვხვდება 500 სახეობის დროზოფილა. ფილოგენეზური კავშირი დადგენილია ეთოლოგიური, მორფოლოგიური და ციტოგენეტიკური (პოლიტენური ქრომოსომები) ნიშნების ანალიზით. ექვს დიდ კუნძულზე ჩამოყალიბებულია დროზოფილების უნიკალური სახეობები. ცალკეულ კუნძულზე, ჩვეულებრივ, ენდემური სახეობები სახლობს. იშვიათია ისეთი სახეობა, რომელიც სხვა კუნძულებზე ბინადრობს. სახეობათა დიდი ნაწილის დამფუძნებელია ერთი განაყოფიერებული მდედრი. ფიქსაციას თან ახლდა სწრაფი დივერგენცია. ასე მაგალითად, კუნძულ ჰავაიზე ახლად ჩამოყალიბებული სახეობები D. heteroneura და D. silvestris საერთო წინაპრიდან (D. planitibia) კვანტური ევოლუციის გზითაა ჩამოყალიბებული.
კვანტური ევოლუციის სანიმუშო მაგალითია ე. მაირის მიერ შესწავლილი გვინეური ალკანი (თანისიპტერა გალატეა). ახალი გვინეა მსოფლიოს ერთ-ერთი უდიდესი კუნძულია. მის ტყეებში გავრცელებულია ალკანის სამი განსხვავებული რასა. ისინი საკმაოდ დიდი ტერიტორიით არიან გამიჯნულნი ერთმანეთისაგან. ცენტრალური კუნძულის გარშემო მრავალი მცირე კუნძული მდებარეობს. ამ მცირე კუნძულებზე გვხვდება ხუთი რასისა და ერთი სახეობის ფრინველი. ეს ფორმები ცენტრალური კუნძულიდან შემთხვევით მოხვედრილი ფრინველთა მცირე ჯგუფიდანაა ჩამოყალიბებული კვანტური ევოლუციის შედეგად.
კვანტური ევოლუციის გზით ჩამოყალიბდა სიერა-ნევადას (კალიფორნია, აშშ) მთიანეთში გავრცელებული ყვავილოვანი მცენარე კლარკია (ოჯახი Onograceae). იგი ერთწლიანი ბალახოვანი მცენარეა. ფართოდ არის გავრცელებული სახეობა ჩლარცია ბილობა. ქრომოსომული ანალიზითა და ეკოლოგიის შესწავლით გამოირკვა, რომ მისგან კვანტურად ჩამოყალიბდა ვიწრო ენდემური სახეობა C. lingulata. ორივე სახეობა მორფოლოგიური ნიშნებით მსგავსია, მხოლოდ უმნიშვნელო განსხვავება აღენიშნებათ გვირგვინის აგებულებაში; მაგრამ ისინი განსხვავდებიან ქრომოსომული ნაკრებით. აქვთ ერთი ქრომოსომული ტრანსლოკაცია და რამდენიმე პარაცენტრული ინვერსია. C. lingulata-ს მოეპოვება დამატებითი ე.წ. B ქრომოსომა, რომელიც ჩ. ბილობა-ს ორი ქრომოსომული ნაწილის ჰომოლოგიურია.
|
|
14.6 12.6. სახეობათა სიმპატრიული წარმოქმნა |
▲ზევით დაბრუნება |
სახეობათა სიმპატრიული წარმოქმნის დროს ახალი სახეობის ჩამოყალიბების პროცესი ძველი სახეობის არეალში მიმდინარეობს. ამ პროცესის სარეალიზაციოდ სულ მცირე ორი პირობის დაცვაა აუცილებელი; 1. პოპულაციის შემადგენელი ჯგუფების სივრცობრივად ან დროში ერთმანეთისგან გამიჯვნა. 2. განცალკევებულ ჯგუფებში რეპროდუქციული იზოლაციის ჩამოყალიბება. ჯგუფების დამოუკიდებლად დიფერენცირების (დამოუკიდებლად ევულირების) მიზეზი არის საარსებო გარემოს ჰეტეროგენულობა (ეკოლოგიური ფაქტორები).
სიმპატრულ სახეობათა წარმოქმნაზე მეტყველებს ერთიან ტერიტორიაზე (ან აკვატორიაში) საერთო წინაპრიდან ჩამოყალიბებული ახლომონათესავე სახეობების არსებობა. დიდ ტბებში (ჩრდ. ამერიკა), ბაიკალში (აზია), ტანგანიკასა (აფრიკა) და სხვა დიდ წყალსატევებში მონათესავე ენდემურ სახეობათა სიმრავლე მათ სიმპატრიული გზით ჩამოყალიბებაზე მიუთითებს (იხ. §11).
ჩრდილოეთ ამერიკაში გავრცელებულ ფუტკრების გვარ Perdita-ში გაერთიანებულია 500 სახეობა. ყველა მათგანი არის მონოფაგი. მათ სიმპატრიულ ევოლუციის პროცესში ჩამოუყალიბდათ ვიწრო კვებითი სპეციალიზაცია. კონკრეტული სახეობა სარჩოს ერთი ან რამდენიმე მონათესავე სახეობის მცენარიდან მოიპოვებს. ფუტკრების ამ გვარში სახეობათა მრავალფეროვნება ნებისმიერი სივრცობრივი გამიჯვნის გარეშე კვებითმა სპეციალიზაციამ გამოიწვია. სიმპატრიულ სახეობათა ჩამოყალიბების სანიმუშო არგუმენტია ვიწროდ სპეციალიზებული მონოფაგი მავნებლის მიერ საკვები ასორტიმენტის შეცვლა. ასე, მაგალითად, სამხრეთ ამერიკაში შემთხვევით მოხვედრილ ვაშლის ნაყოფჭამია მწერში რამდენიმე ათეული წლის შემდეგ ჩამოყალიბდა კაკლისა და მსხლის მონოფაგი მავნებელი რასები. ცნობილია მცენარეთა, ცხოველთა და მიკროორგანიზმთა პარაზიტული ფორმები, რომლებიც კონკრეტულ მასპინძელში ბუდობენ.
ე. მაირის შეხედულებით ვიწრო კვებითი სპეციალიზაციის მფლობელი რასების ჩამოყალიბება წარმოადგენს ერთადერთ მნიშვნელოვან არგუმენტს, რაც სიმპატრიულ სახეობათა წარმოქმნის საწყისი ეტაპის დაწყებაზე მიუთითებს. მათი ახალ სახეობრივ ფორმებად საბოლოო სტაბილიზაცია სივრცობრივი ბარიერების მეშვეობით ხორციელდება. ჩვენს მიერ ზემოთ განხილული ჰავაის კუნძულებზე გავრცელებული დროზოფილას სახეობის 70%25 ვიწრო სპეციალიზებული მონოფაგებია (იხ. სურ. 12.5). ისინი სარჩოს კონკრეტული სახეობის მცენარიდან ან მისი რომელიმე ორგანოდან მოიპოვებენ; მიუხედავად იმისა, რომ ყოველი მათგანი სოკოებისა და მიკროორგანიზმების მეტაბოლური პროდუქტებით იკვებებიან. ასე, მაგალითად, კუნძულ ჰავაიზე გავრცელებული დროზოფილას ორი გენეტიკურად ახლომდგომი (D. silvestris და D. heedi) მკაცრად ჰომოსეკვენტური (პოლიტენურ ქრომოსომებში დისკოების მსგავსი განლაგება აღენიშნებათ) სიმპატრიული სახეობებია. პირველი სახეობის მდედრები კვერცხს დებენ ხის (Myoporum sandwicens) ზედა ნაწილებიდან გამოყოფილ წვენში, ხოლო მეორენი ამავე ხის გამონადენ სითხეში, ოღონდ მიწაზე. ძალზე რთულია ამდაგვარი ადაპტაცია, ალოპატრიულ სახეობათა წარმოქმნის კონცეფციით აიხსნას.
პოპულაციაში სეზონური რასების თანაპოვნიერება სიმპატრიული ევოლუციის მიმდინარეობის სარწმუნო არგუმენტს წარმოადგენს. მცენარეებში ცნობილია მკვეთრად გამოხატული გენეტიკურად დეტერმინირებული ფენოლოგიური რასები. ეს მოვლენა კარგად არის შესწავლილი ნ. ცინგერის მიერ ნახევრად პარაზიტ მცენარე ხრიალაზე (Alectorolophus majon) დაკვირვების გზით (იხ. §7.5). იგი ყამირ მდელოზე მთელი ზაფხულის განმავლობაში ყვავილობს. პოპულაციაში გვხვდება ადრე და გვიანმოყვავილე ფორმები. ეს ნიშანი გენეტიკურად არის დეტერმინირებული. ქვესახეობა A.m. montanus საწყის ფორმადაა მიჩნეული. სათიბ მდელოზე ადამიანის უნებური მოქმედებით (გათიბვა) შენარჩუნებულ იქნა ადრემოყვავილე ინდივიდები, საიდანაც ჩამოყალიბდა ადრემოყვავილე ზაფხულის ფორმა (A. m. aestivalis), მას შემდეგ, რაც წელიწადში მდელოს ორჯერ გათიბვა დაიწყეს, ეს უკანასკნელი თავის მხრივ ორ ფორმად გაიყო: საადრეო, საზაფხულო ფორმად (A. m. vernalis) და საგვიანო, საზაფხულო ფორმად (A. m. polycladus). ისინი ერთმანეთისგან რეპროდუქციულად იზოლირებულნი არიან. გვხვდება კიდევ ერთი ქვესახეობა A. m. apterus. იგი მხოლოდ ჭვავის ნათესებს ასარეველიანებს. ხრიალასა და ჭვავის სასიცოცხლო ციკლი და მორფოლოგიური ნიშნები ერთიმეორის მსგავსია. ისინი ერთდროულად მწიფდებიან. მათი თესლები წონითა და ფორმით ერთმანეთს გვანან. სარეველა ხრიალა სხვა ქვესახეობებისაგან განსხვავებით იკეთებს უხსნად კოლოფს და უფრთო თესლს. ჭვავისა და ხრიალას თესლები ლეწვისა და განიავების დროს ძნელად სცილდება ერთმანეთს. ადამიანი სარეველას მოსავალთან ერთად მკიდა, ლეწავდა, ხოლო შემდეგ თესავდა. ამრიგად, ხრიალაში ერთი ფორმიდან გადარჩევის შედეგად რამდენიმე ფორმა ჩამოყალიბდა. თუ გადარჩევის პირობები შენარჩუნებული იქნება, მაშინ შესაძლებელია თითოეული მათგანისაგან ახალი სახეობები ჩამოყალიბდეს (იხ. სურ. 7.6).
„საგაზაფხულო“ და „საშემოდგომო“ ფორმები გვხვდება ცხოველებშიც (ზოგიერთი მწერი, სალამურა, თევზი და სხვ.). ეს ფორმები მორფოლოგიურად მსგავსია, გენეტიკურად კი განსხვავდებიან. სახეობათა სიმპატრიული წარმოქმნისათვის დამახასიათებელია ძველი სახეობიდან მორფო-ფიზიოლოგიურად მსგავსი ახალი სახეობის წარმოქმნა.
სახეობათა წარმოქმნის სიმპატრიული კონცეფციის მიხედვით საწყისი პოპულაციის ეკოლოგიურ რასებში მიმდინარეობს დიფერენცირების პროცესი და ყალიბდება რეპროდუქციული იზოლაცია. საიზოლაციო მექანიზმების ჩამოყალიბების შემოთავაზებული ახსნა ნაკლებ დამაჯერებელია და ამ კონცეფციის სუსტ მხარეს წარმოადგენს. მოიძებნა კომპრომისული გზაც. გამოჩენილმა ევოლუციონისტმა და ზოოლოგმა მ. უაიტმა წამოაყენა მოსაზღვრე - სიმპატრიული სახეობათა წარმოქმნის კონცეფცია (სინონიმებია: სტასიმპატრიული, პარასიმპატრიული). მან შეისწავლა გვარ ავსტრალიურ კუტკალიაში (Morabinae) გაერთიანებული 160 სახეობა და ქვესახეობა. ამ ჯგუფში გამოვლენილია ევოლუციის პროცესში დაფიქსირებული 34 განსხვავებული ტიპის ტრანსლოკაცია. ინტერესს აღძრავს ორი კომპლექსური სახეობა Moraba viatica და M. scurra. ქვესახეობები ჰომოკარიოტიპებია (ჰომოზიგოტური სხვადასხვა ტრასლოკაციების მიხედვით) და უკავიათ მომიჯნავე ტერიტორიები. მათ საზღვარზე ჰიბრიდები ვიწრო ზონას ქმნიან. ჰიბრიდები ჰეტეროკარიოტიპებია (ჰეტეროზიგოტი სხვადასხვა ტრანსლოკაციების მიხედვით). ისინი სიცოცხლის უნარიანობით, ნაყოფიერებითა და მოძრაობის უნარით მნიშვნელოვნად ჩამორჩებიან საწყის ფორმებს. ამრიგად, ქვესახეობები რეპროდუქციული მექანიზმით გამიჯნულია ერთმანეთისგან.
ცნობილია ფაქტები, როდესაც ბუნებაში სახეობათშორისი ჰიბრიდიზაციისას ხშირად მიიღება სიცოცხლისუნარიანი ჰიბრიდები, მაგრამ მათში დაბალი ცხოველმყოფელობის გამეტები ფორმირდება, რის გამოც ისინი დაბალი ნაყოფიერებით ხასიათდებიან. ჰიბრიდები ჩვეულებრივ დაბალკონკურენტუნარიანნი არიან, ამიტომაც მომდევნო თობებში მათი რაოდენობა კატასტროფულად მცირდება და ელიმინირდება. მათ ადგილს ახლად ჩამოყალიბებული ჰიბრიდები იკავებენ. ორ სახეობას შორის საზღვარი იზოლირებულია „ჰიბრიდიზაციის მეშვეობით“.
ამდაგვარი ჰიბრიდული ბარიერი აბრკოლებს სახეობების შერწყმას და ერთიანი გენოფონდის ჩამოყალიბებას. როგორც უხერხემლო, ისე ხერხემლიან ცხოველებში აღწერილია „ჰიბრიდული ზონით“ იზოლირებული სახეობები. კლასიკურ ნიმუშს წარმოადგენს რუხი (Corvus cornix) და შავი (C. corone) ყვავის არეალების ჰიბრიდული ზონით შემოფარგვლა (იხ. სურ. 12.6).
კავკასიის ტყიან სარტყელში გავრცელებული მურა ბაყაყი (ღანა მაცროცრემის) პოლიტიპური სახეობაა. მცირე კავკასიონზე ბინადრობს ორი ქვესახეობა (R. m. macrocremis და R. m. camerani) რომელიც ჯავახეთის ზეგანის ჩრდილოეთ ნაწილში ვიწრო ჰიბრიდული ზოლით არის გამიჯნული (თარხნიშვილი, 2004).
სიმპატრიულ სახეობათა წარმოქმნის პრობლემა არსებით რაკურსს იძენს სისტემურ მუტაციებთან დაკავშირებით. სალტაციონიზმის მიხედვით სისტემური მუტაციის (სალტაციის) მფლობელი ინდივიდი ან ინდივიდთა მცირე ჯგუფი რეპროდუქციულადაა იზოლირებული საწყისი სახეობის ინდივიდებისაგან. ამ სახის მუტაციის ინდუქციით შესაძლებელია ერთი სახეობა მყისიერად გარდაიქმნას მეორე სახეობად (იხ. §12.1). ობიექტურობა მოითხოვს, აღინიშნოს, რომ გარკვეულ ხელსაყრელ პირობებში, ამდაგვარი სიტუაციის რეალიზაცია შესაძლებელია, მაგრამ სიტუაციის ალბათობა საკმაოდ დაბალია.
სურ. 12.6. ახლომონათესავე სახეობის რუხი Corvus cornix (1) და შავი C. corone (2) ყვავის გავრცელება. A. მთლიანი არეალი. B. არეალის ფრაგმენტი. სახეობათა შეხვედრის საზღვარზე წარმოქმნილი ჰიბრიდული ზონა მოცემულია შავი ზოლით. საწყის ფორმებთან
|
|
14.7 12.7. სახეობათა მყისიერი წარმოქმნა |
▲ზევით დაბრუნება |
|
|
14.7.1 პოლიპლოიდია. |
▲ზევით დაბრუნება |
მცენარეთა ევოლუციაში სისტემურმა მუტაციებმა არსებითი როლი შეასრულა. ამ კატეგორიას მიეკუთვნება პოლიპლოიდია. ბუნებაში პოლიპლოიდია ორი ფორმით გვხვდება: ავტოპლოიდია (პროპორციულად გადიდებულია ერთი და იგივე გენომი) და ალოპლოიდია (ჯერადად გადიდებულია სხვადასხვა სახეობის გენომი).
სურ. 12.7. სახეობათა წარმოშობა პოლიპლოიდიის გზით. ხორბლისა (A) და ქრიზანთემას (B) ქრომოსომული კომპლექტები I მეიოზური გაყოფის მეტაფაზური ფირფიტების მიხედვით. 1 - Triticum aegilopoides (N=7), 2 - T. dicoccum (n=14), 3 - T. aestivum (n=21), 4 - Chrisanthemum makinoi (n=9), 5 - Ch. indicum (n=18), 6 - Ch. japonense (n=27), 7 - Ch. ornatum (n=36), 8 - Ch. gezoensis (n=45), 9 - Ch. pacificum (45). (a. miutcingi, 1967).
პოლიპლოიდიით ხორციელდება სახეობის მყისიერად ჩამოყალიბება. იგი გენომის უეცარ გარდაქმნას იწვევს, რაც ავტოპლოიდიის დროს ხდება. პოლიპლოიდია ფართოდაა გავრცელებული მცენარეებში. ცხოველებში კი ერთობ იშვიათია. მცენარეთა ზოგიერთ გვარში გაერთიანებული მონათესავე სახეობები ქრომოსომთა ჯერადი რიცხვით განსხვავდებიან. იგი საწყის სახეობაში მომხდარი ქრომოსომთა რიცხვის პროპორციულად გადიდების შედეგია. ასეთ გვარებში გაერთიანებული სახეობები პოლიპლოიდურ რიგს ქმნიან. მონათესავე სახეობათა ჯგუფს, რომლის ქრომოსომთა ჰაპლოიდური ნაკრებით (გენომით) იქმნება ჯერადად გაზრდილი აღმავალი მწკრივი პოლიპლოიდური რიგი ეწოდება. ასე მაგალითად, ძაღლყურძენას გვარში (Solanum) ქრომოსომთა ჰაპლიდური კომპლექტი არის N=6, პოლიპლოიდური რიგი წარმოდგენილია 12, 24, 36, 48, 60, 72, 96, 108, 144 ქრომოსომიანი სახეობით. ბუნებრივი პოლიპლოიდური რიგები გვხვდება ვარდის, მარწყვის, ქრიზანთემას, ჭანგას, ხორბლის, შვრიის და სხვა გვარებში (იხ. სურ. 12.7). ცხოველებიდან პოლიპლოიდური რიგები ცნობილია ჰერმაფროდიტ ცხოველებში (მაგ., ჭიაყელა), მდედრსქესიან ჰერმაფროდიტ სახეობებში (ზოგიერთი ხოჭო, პეპელა, ბაღლინჯო, კიბოსნაირი, თევზი, სალამანდრა და სხვ.).
მცენარეებში პოლიპლოიდია ფართოდ და არათანაბრად არის გავრცელებული. პოლიპლოიდური სახეობები დიდი რაოდენობით გვხვდება ყვავილოვან მცენარეებში, მათი სიხშირე 47%25-ია (ორლებნიანებში - 43%25, ერთლებნიანებში - 58%25). შიშველთესლოვანებში იშვიათია - პოლიპლოიდური სახეობის სიხშირე 1,5%25-ს შეადგენს. საგოვანებში საერთოდ არ გვხვდება პოლიპლოიდური სახეობები, მაშინ როდესაც გვიმრანაირებში მათი სიხშირე 95%25-ს აღწევს. ბუნებაში გაცილებით დიდი რაოდენობა ალოპლოიდები გვხვდება. პოლიპლოიდებისთვის, განსხვავებით საწყისი დიპლოიდებისგან, ჩვეულებრივ ნიშანდობლივია სხეულის მძლავრი განვითარება: ეზრდებათ დიდი ზომის ფოთლები, ყვავილები, ნაყოფი და თესლები. აქვთ საკმაოდ ძლიერი და უხეში ღეროები. მეტწილად დიდი ზომისაა პოლიპლოიდური უჯრედი და ბირთვი. უმეტესად პოლიპლოიდები გამოირჩევიან გარემოს ფლუქტუირებადი ფაქტორებისადმი მედეგობით, საწყის ფორმებთან შედარებით. ამიტომაც ისინი ეგუებიან მკაცრ საარსებო პირობებს. არქტიკისა და მაღალმთიანი რაიონების ფლორა მდიდარია პოლიპლოიდური ფორმებით (იხ. სურ. 12.8).
ბოლო მონაცემებით დადასტურებულია, რომ პოლიპლოიდიამ საწყის ეტაპზე მნიშვნელოვანი როლი ითამაშა ხერხემლიან ცხოველთა ევოლუციაში. სახეობათა პოლიპლოიდური ფორმები, გვარები და მთელი ოჯახებიც კი, გამოვლენილია თევზებში, ამფიბიებსა და რეპტილიებში. ასე მაგალითად, თევზებში ოჯახი ორაგულისებრნი, კაპოეტასებრნი და ქაშაყისებრნი პოლიპლოიდური წარმოშობისაა. პოლიპლოიდური ფორმები საკმაო რაოდენობითაა გამოვლენილი თევზების სხვა ოჯახებშიც (მაგ. კობრისებრნი). სამხრეთ ამერიკაში გავრცელებული ბაყაყის ოჯახ Caratophrididae-ში გაერთიანებულია ავტოპლოიდური (ტეტრა - და ოქტაპლოიდური) სახეობები. პართენოგენეზურ ხვლიკებს (გვარი Lacerta და Cnemidophoris) ქრომოსომთა კომპლექტების სხვადასხვა რაოდენობა მოეპოვებათ.

სურ. 12.8. ყვავილოვან მცენარეთა პოლიპლოიდური სახეობების გავრცელება ევრაზიის სხვადასხვა რაიონის ფლორაში (პროცენტებში სახეობათა საერთო რაოდენობასთან შედარებით, შედგენილია სხვადასხვა ავტორების მონაცემების მიხედვით, ნ. ტიმოფეევ - რესოვსკი და სხვ., 1977).
მაშასადამე, პოლიპლოიდია იწვევს გენომის კარდინალურ ცვლილებას, რამაც შექმნა წინაპირობა როგორც მცენარეთა, ისე ცხოველთა სამეფოში დაწყებულიყო ფართო მასშტაბიანი ევოლუციური პროცესები. პროტისტებს, კერძოდ, ინფუზორიებს პოლიპლოიდიის გზით უყალიბდებათ რთული მრავალგენომიანი ბირთვი (მაკრონუკლეუსი). პოლიპლოიდური სახეობები აღწერილია ეუკარიოტულ მიკროორგანიზმებშიც (მაგ., სოკოები და ერთუჯრედიანი წყალმცენარეები). ე. მაირის დასკვნით, პოლიპლოიდია არის სახეობათა მყისიერად წარმოქმნის ერთადერთი უნივერსალური გზა.
|
|
14.7.2 ჰიბრიდოგენეზი. |
▲ზევით დაბრუნება |
ზოგჯერ ახალი სახეობა ორი წინარე განსხვავებული სახეობის გენომის გაერთიანებით ყალიბდება. ჰიბრიდოგენეზი არის მყისიერ სახეობათა წარმოქმნის ერთ-ერთი გზა. სახეობათა ამდაგვარი წარმოქმნა ძირითადად მცენარეებისათვის არის დამახასიათებელი, მაგრამ აღწერილია სხვა (ცხოველები, სოკოები) ფორმებშიც. მართალია, სახეობათაშორისი ჰიბრიდიზაცია საიზოლაციო მექანიზმებით იზღუდება, მაგრამ ეს პროცესი ბუნებაში მაინც ხორციელდება. რეკომბინაციული მექანიზმების მოქმედების შედეგად ჰიბრიდული შთამომავლობა კოლოსალურ ცვალებადობას განიცდის. სახეობათაშორის ჰიბრიდებში ირღვევა ძველი სახეობრივი კოადაპტირებულ გენთა სისტემები და გენოტიპთა ნაირგვარი ვარიანტები წარმოიქმნება. ზოგიერთი საარსებო გარემოსადმი იმდენად კარგადაა ადაპტირებული, რომ მათგან ახალი სახეობრივი ფორმები იღებენ სათავეს.
ჰიბრიდოგენეზი არის წინარე სახეობის გენომთა ან მათი ნაწილის გაერთიანების გზით ახალ სახეობათა ფორმირების პროცესი. იგი „ბადებრივი“ ევოლუციის ერთ-ერთი გამოვლენაა, კერძოდ, ახალი სახეობის ჩამოყალიბება ადრე არსებული გენომების რეორგანიზაციის (კომბინირების) შედეგია.
მცენარეებში სახეობათაშორისი ჰიბრიდები თითქმის ყველა დიდ ტაქსონში გვხვდება. ამ სახის ჰიბრიდები დიდი რაოდენობით ხემცენარეებიდან ტირიფში, მუხასა და ევკალიპტში, ხოლო ბალახოვანი მცენარეებიდან იაში, მზესუმზირასა და წყალიკრეფიაში გვხვდება. გვიმრანაირებიდან სახეობათაშორისი ჰიბრიდიზაციითაა ჩამოყალიბებული გვარი Pteris. სახეობათაშორისი შეჯვარება ზოგჯერ ინტროგრესიული ჰიბრიდიზაციის მიმართულებისაა. ინტროგრესიული ჰიბრიდიზაცია განაპირობებს აღმავალი შეჯვარებისას (ჰიბრიდების შეჯვარება მშობელ ფორმებთან) ცალკეული გენების, გენთა ბლოკების, ქრომოსომების მონაკვეთის ან გენომის ნაწილის ერთი სახეობიდან მეორეში გადატანას. ეს პროცესი თავისუფლად დამტვერვად მცენარეებში (ფიჭვი, მუხა, ტრადესკანცია და მისთ.) ახალი რასების ჩამოყალიბებაში გადამწყვეტ როლს ასრულებს.
ამერიკელი ევოლუციონისტის ვ. გრანტის განმარტებით, ჰიბრიდოგენულ სახეობათა წარმოქმნა არის ახალი სახეობის უშუალო ჩამოყალიბება რომელიმე წინარე ბუნებრივი ჰიბრიდიდან. მცენარეებში ეს გზა პოლიპლოიდიითა და აპომიქსისის მეშვეობით რეალიზდება. ცნობილია, რომ სახეობათაშორისი ჰიბრიდები გენეტიკური საიზოლაციო მექანიზმების მოქმედების გამო სტერილურებია (იხ. §7.5). ალოპლოიდია და აპომიქსისი ჰიბრიდებში ფერტილობის აღდგენას უზრუნველყოფენ, რითაც მყისიერი სახეობათაწარმოქმნის წინაპირობა იქმნება. განსხვავებული სახეობების შეჯვარების გზით წარმოქმნილ სტერილურ ჰიბრიდებში, ქრომოსომათა რიცხვის გაორმაგებით (ე.წ. ალოპლოიდია) მიიღწევა ფერტილობის აღდგენა. ზოგიერთი მონაცემებით მცენარეთა სახეობის დაახლოებით 50%25 ალოპლოიდიის გზით არის ჩამოყალიბებული.
XXს. 20-იან წლებში ექსპერიმენტულად პირველი ალოპლოიდი ე.წ. ამფიდიპლოიდი მიიღო ცნობილმა რუსმა გენეტიკოსმა გ. კარპეჩენკომ. მან შეძლო გვართა შორის ჰიბრიდებში (კომბოსტო და ბოლოკის ნაჯვარი) პოლიპლოიდიის მეთოდით ფერტილური ჰიბრიდული (ღაპჰანობრასიცა) შთამომავლობის მიღება.
გამოჩენილმა გენეტიკოსმა ა. მიუტცინგმა გამოავლინა, რომ ჩრდილოეთ ევროპასა და აზიაში გავრცელებული ერთწლიანი ბალახოვანი მცენარე თავცეცხლა Galeopsis tetrahit (2n=32) alotetraploidia (amfidiploidi), რომელიც დასაბამს: G. pubescens (2n=16) და G. speciosa (2n=16) დიპლოიდი მცენარეებიდან იღებს. ჰიბრიდოგენული გზითაა ჩამოყალიბებული ხორბალში (Triticum) გაერთიანებული პოლიპლოიდური სახეობები. პოლიპლოიდური რიგი წარმოდგენილია 14, 28 და 42 ქრომოსომიანი ფორმებით. ველური ცალმარცვალა ხორბალი T. monococcum (2N=14) ბუნებრივ პირობებში შეეჯვარა ველურ მარცვლოვან egilopss Aegilops speltoides (2N=14) და inducirda alotetraploidi (2N=28; მაგ., თავთუხი T. durum) ხორბლები. ამ უკანასკნელს ბუნებრივ პირობებში შეეჯვარა ეგილოპსის სრულიად სხვა სახეობა E. squarosa (2N=14) და ჩამოყალიბდა ალოჰექსაპლოიდი (2N=42 მაგ. T. aestivum) ხორბლები. ასევე კულტურული ქლიავი Prunus domenstica (2ნ=48) ბუნებრივად წარმოიშვა კვრინჩხის (P. spinosa, 2n=32) და ალუჩის (P. divericata, 2n=16) ჰიბრიდიზაციით და შემდგომ ქრომოსომთა გაორმაგებით.
მცენარეებს ევოლუციის პროცესში ჩამოუყალიბდათ მეორადი უსქესო გამრავლების ფორმა აპომიქსისი (იგი ძირითადად ორი ფორმით ვლინდება: პართენოგენეზი - კვერცხუჯრედიდან ჩანასახის ჩამოყალიბება; აპოგამია-ვეგეტატიური უჯრედიდან ჩანასახის ჩამოყალიბება). აპომიქტურად მრავლდებიან გენეტიკურად გამორჩეული ჰიბრიდული (არა ყოველთვის) წარმომავლობის სახეობები. აპომიქტურ სახეობებში ფიქსირდება ნებისმიერი არაბალანსირებული ქრომოსომების (ტრიპლოიდი, ანეუპლოიდი) შემცველი კარიომორფები. აპომიქტური კომპლექსური სახეობები ბუნებრივი ჰიბრიდიზაციისა და აგამური გამრავლების პროცესთა შეთანაწყობის საფუძველზე ყალიბდებიან. ასეთი სახეობების სიუხვით გამოირჩევიან გვარები: Poa, Potentilla, Rubus, Hieracium. ისინი ფლობენ ვრცელ არეალს და ითვისებენ არახელსაყრელ კლიმატურ რაიონებს. გამოირჩევიან სახეობათა და მიკროსახეობათა დიდი მრავალფეროვნებით. სქესობრივი გამრავლების უნარის დაკარგვის გამო აპომიქტური ფორმები ევოლუციურ პერსპექტივას მოკლებულნი არიან. ამ თვალთახედვით ყველა აგამური კომპლექსი ქმნის დახშულ გენეტიკურ სისტემას და ევოლუციურ „ჩიხს“ წარმოადგენს.
ცხოველებიდან ბუნებრივი სახეობათშორისი ჰიბრიდიზაცია გამოვლენილია როგორც უხერხემლო (მაგალითად, ზოგიერთი პეპელა, ციკადები), ისე უმდაბლეს ხერხემლიან (თევზები, ამფიბიები, რეპტილიები) ცხოველთა ზოგიერთ რიგში.
ჰიბრიდოგენული კომპლექსები გამოვლენილი და შესწავლილია თევზებში - რიგ კობრკბილიანებში (Cyprinodontiformes). ამ რიგში გაერთიანებული კვერცხცოცხალმშობი თევზი Poecilia formosa ბინადრობს ტეხასის შტატისა (აშშ) და ჩრდილოეთ მექსიკის წყალსატევებში. იგი გინოგენეზური სახეობაა. პოპულაციები მხოლოდ მდედრებისგან (2C=46) შედგება. მათ ანაყოფიერებს სხვა მონათესავე სახეობა, რომლის სპერმა ქვირითზე მხოლოდ მასტიმულირებელ გავლენას ახდენს. გამოვლენილია ზოგიერთ პოპულაციაში გინოგენეზური ტრიპლიოიდური (2C=69) კლონები. ისინი P. formosa-სა და P. mexicana-ს შეჯვარებითაა წარმოქმნილი.
თევზების ამავე რიგში გაერთიანებული გვარ Foeciliopsis-ში გვხვდება ბისექსუალი დიპლოიდური (2N=48) ფორმა, ასევე ცალსქესიანი დიპლოიდური (2N=48) და ტრიპლოიდური (3N=72) ჰიბრიდული წარმოშობის სახეობები. დიპლოიდური მდედრები პართენოგენეზურად მრავლდებიან. ცალსქესიანი სახეობიდან დიპლიდური მდედრების ჩამოყალიბება კრედიტოგენეზის გზით ხორციელდება, რაც ხერხემლიან ცხოველებში იშვიათი მოვლენაა. კერძოდ მდედრებში (განვითარებულია ჰიბრიდული ზიგოტისაგან) მიმდინარე ოოგენეზის პროცესში შერჩევით ელიმინირდება მამისეული ქრომოსომები. მომწიფებული კვერცხუჯრედები მხოლოდ დედისეულ ჰაპლოიდურ გენომს შეიცავს. ინდივიდში მამისეული გენომი მხოლოდ ერთი თაობის სიცოცხლეშია შენარჩუნებული.
ჰიბრიდული წარმოშობის პართენოგენეზური და გინოგენეზური სახეობები შესწავლილია კუდისნაირ ამფიბიებში (ჩრდილო-ამერიკული ამბისტომები). სტაბილური ჰიბრიდული ფორმაა ჯგუფი მწვანე ბაყაყები (Rana esculenta). მის ჩამოყალიბებაში მოქმედებს კრედიტოგენეზის მექანიზმი.
ჰიბრიდოგენეზის შედეგია ამიერკავკასიაში გავრცელებული კლდის ხვლიკის ოთხი სახეობა: დალისეული ხვლიკი (Lacerta dahli), თეთრმუცელა ხვლიკი (L. unisexualis), როსტომბეკოვისეული ხვლიკი (L. rostombekovi) და სომხური ხვლიკი (L. armenica). პირველი სამი სახეობა საქართველოშია გავრცელებული. მამრები ამ სახეობებში არ გვხვდება, ან ძალიან იშვიათია. ისინი ჩამოყალიბებულია ორსქესიანი მშობლებიდან ბუნებრივი ჰიბრიდიზაციის გზით და პართენოგენეზურად მრავლდებიან (იხ. სურ. 12.9).
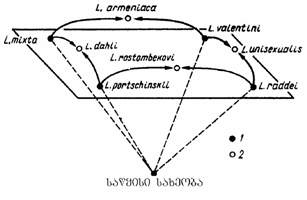
სურ. 12.9. გვარი Lacerta-ში (Darevskia) გაერთიანებულ კავკასიური კლდის ხვლიკების ჰიბრიდოგენური სახეობათა წარმოქმნა. 1 - დიპლიოლური ბისექსუალური საწყისი სახეობები. 2 - ჰიბრიდული წარმოშობის დიპლოიდური ერთსქესიანი სახეობები (ლ. კაიდანოვი, 1996).
ყველა განხილულ შემთხვევაში შორეული ჰიბრიდიზაციის შედეგად წარმოშობილი სახეობების ფორმირება და შენარჩუნება განხორციელებულია სქესობრივი გამრავლების არარეგულარული ტიპების - პართენოგენეზის, გინოგენეზისა და კრეტიდოგენეზის მეშვეობთ.
|
|
14.7.3 სახეობათა ქრომოსომული წარმოქმნა |
▲ზევით დაბრუნება |
ბუნებრივი პოპულაციებისათვის ნიშანდობლივია ქრომოსომული ცვალებადობის ფართო სპექტრი. იგი შიგაპოპულაციური ქრომოსომული პოლიმორფიზმის სახით ვლინდება (იხ. § 9.2). გვარ Drosophila-ში შესწავლილ სახეობების 75%25-ს ინვერსიული პოლიმორფიზმი აღენიშნება. თევზებში ქრომოსომული პოლიმორფიზმის სიხშირე 1%25-ს, ხოლო რეპტილიებში 2%25-ს შეადგენს. ძუძუმწოვრების პოპულაციებს მოეპოვებათ შემდეგი სახის ქრომოსომული მუტაციები: რობერტსონული ტრანსლოკაციები, პერიცენტრული ინვერსიები, ჰეტეროქრომატინული ბლოკების მატება ან კლება. ძუძუმწოვრების კლასში გაერთიანებულ 4000-მდე სახეობისა და ქვესახეობის შესწავლის საფუძველზე დადგენილია, რომ შიგაპოპულაციური ქრომოსომული პოლიმორფიზმის სიხშირე 10.9%25-ს შეადგენს.
კითხვა ისმის რა როლს ასრულებს ქრომოსომული პოლიმორფიზმი ევოლუციის პროცესში? პასუხი არაერთგვაროვანია. ევოლუციის სინთეზური თეორიის თვალსაზრისი ტრადიციულია. ამ თეორიის თანახმად (პოპულაციონისტური კონცეფციიდან გამომდინარე), შიგაპოპულაციური გენეტიკური პოლიმორფიზმი, მათ შორის ქრომოსომული არის გენეტიკური შემადგენლობის ადაპტური დიფერენცირების გამოხატულება. ალოპატრიულ სახეობათა წარმოქმნის პროცესში მცირე მუტაციების მეშვეობით მიმდინარეობს, ძირითადად, გენეტიკური სტრუქტურის თანდათანობითი გარდაქმნა. გენეტიკური სტრუქტურის დიფერენცირება რეპროდუქციული საიზოლაციო მექანიზმების ჩამოყალიბებითა და ახალი სახეობის ფორმირებით სრულდება.
ამავე შეკითხვაზე თანამედროვე სალტაციონიზმსა და ყველა იმ მიმართულებას, რომელიც ტიპოლოგიურ კონცეფციას ეფუძნება, ასევე ტრადიციული პასუხი მოეპოვება. კერძოდ, ქრომოსომული პოლიმორფიზმი (ასევე შიგაპოპულაციური პოლიმორფიზმის ნებისმიერი ფორმა) ყალიბდება პოპულაციის საჭიროებისათვის. მას სახეობათა წარმოშობასთან არავითარი კავშირი არა აქვს. სახეობათა ფორმირების პროცესი ინიცირდება ცალკეულ ინდივიდში ან ინდივიდთა მცირერიცხოვან ჯგუფში ქრომოსომული გარდაქმნით (სისტემური მუტაციებით), რომელიც მაშინვე სრულ ან ნაწილობრივ იზოლაციას აყალიბებს. ამ მექანიზმებით მათი მფლობელი ინდივიდები იზოლირდებიან საწყისი ფორმებისგან.
მაკრომუტაციას (სისტემური მუტაცია) რამდენიმე შთამომავალი ფლობს, რომელთა შეწყვილებითაც იგი შთამომავლობაში ფიქსირდება. ამრიგად ინდივიდთა საკმაოდ მცირერიცხოვანი ჯგუფი ემიჯნება პოპულაციის დანარჩენ წევრებს და ახალ სახეობრივ ფორმებს აძლევს დასაბამს. ამ თეზით იხსნება დიდი წინააღმდეგობა ერთეული უნიკალურ სისტემური მუტაციის (სალტაციის) მემკვიდრეობით გადაცემის შესაძლებლობაზე. რადგან, მაკრომუტაციის მფლობელ ინდივიდს მცირე ჯგუფებშივე მოეპოვება ანალოგიური ცვლილების მქონე მეწყვილე და მათი ურთიერთშეჯვარებით სალტაცია შთამომავლებს მემკვიდრეობით გადაეცემა.
მ. უაიტი თავის ფუნდამენტურ მონოგრაფიაში „სახეობათა წარმოქმნის საშუალებები“ (1978), აღნიშნავს: „ქრომოსომათა სტრუქტურული გარდაქმნები, როგორიცაა ინვერსიები და ტრანსლოკაციები მთავარ, შესაძლოა პირველხარისხოვან როლსაც კი ასრულებენ მრავალი, ალბათ უმეტეს სახეობათა წარმოქმნაში“. მან სახეობების ამ გზით ჩამოყალიბების პროცესს სტასიპატრიულ სახეობათა წარმოქმნა უწოდა. ანალოგიურ თვალსაზრისს ავითარებდა გამოჩენილი ევოლუციონისტი და ზოოლოგი ნ. ვორონცოვი (2004). მისი შეხედულებით, სახეობათაწარმოქმნა ორი გზით ხორციელდება: 1. „ჩვეულებრივი“ - დაკავშირებულია პოპულაციათა შორის განსხვავების თანდათანობით დაგროვებასთან, რომელიც რეპროდუქციული იზოლაციის ფორმებით სრულდება. 2. „გენეტიკური“ - იწყება რეპროდუქციული იზოლაციით (რომელიც ქრომოსომული გარდაქმნებით ყალიბდება), მხოლოდ ამის შემდეგ მიმდინარეობს გენური და ეკოლოგიური განსხვავებების ფორმირება.
იაპონელი ციტოგენეტიკოსი ს.ონო მიიჩნევს, რომ ქრომოსომულ ევოლუციაში მნიშვნელოვან როლს გენეტიკური მასალის დუპლიკაცია იძენს. გრანდიოზული ევოლუციური ნახტომები (აროგენეზი) როგორიცაა მრავალუჯრედიანების, ხერხემლიანების, ბოლოს კი ძუძუმწოვრების წარმოშობა, დუპლიკაციის გზით ახალი გენების ჩამოყალიბებისა და მისი შემდგომი დივერგენციის გარეშე შეუძლებელი იქნებოდა.
ამჟამად ხელთ გვაქვს მრავალი არგუმენტი, რომლებიც მიუთითებს, რომ ქრომოსომათა სტრუქტურული გარდაქმნითა და მათი კომბინაციით არის ჩამოყალიბებული მრავალი სახეობა. ასე მაგალითად დროზოფილას გვარში გაერთიანებული სახეობა D. melanogaster და D. simulans ორი ინვერსიული ცვლილებით ჩამოყალიბდა, ასევე ამავე გვარში ორი სახეობა D. pseudoobscura და D. miranda მრავალი ინვერსიული გარდაქმნის შედეგად ფორმირდება. დროზოფილას გვარში გაერთიანებული ვირილის ჯგუფის სახეობის ევოლუცია ტრანსლოკაციების, ინვერსირების, დუპლიკაციების ინდუქციამ და ჰეტეროქრომატინული ბლოკების კლებამ გამოიწვია (იხ. სურ. 12.10).

სურ. 12.10. გვარ Drosophila-ში გაერთიანებული ჯგუფ ვირილის-ის ჰაპლოიდურ ქრომოსომული კომპლექტები. ლათინური ანბანით აღნიშნულია გენთა ბლოკები, ასოთა შეცვლილი თანამიმდევრობა ინვერსიას გამოხატავს; X, 2-3 კი ქრომოსომებს (ჯ. პეტერსონი, 1952).
ქრომოსომულ ევოლუციაში მნიშვნელოვან როლს რობერტსონული ტრანსლოკაციები იძენს. ზოგიერთ ძუძუმწოვართა სახეობების (მღრღნელები) კარიომორფები რობერტსონულ მარაოს ქმნიან. მაგალითად იტალიაში - სახლის თაგვი, აფრიკაში - ჩია თაგვი, პამირ-ალტაიში - ბრუცუნა (დაწვრილებით განხილულია § 9.2). ნ. ვორონცოვი ბრუცუნას (Ellobius talpinus) ფართო სპექტრის კარიოტიპების ფორმირებას - ერთგვარ აფეთქებას, სეისმოაქტიობას უკავშირებს. ბრუცუნა არის ზესახეობა, რომელშიც მრავალი ფორმირებადი in situ nascendi სახეობაა გაერთიანებული. ასეთ რეგიონში ინდივიდზე ნაირგვარი მუტაგენური ფაქტორი მოქმედებს (ၧ - დასხივება, რადონის შემცველი წყალი, მძიმე მეტალთა ჭარბი თანაპოვნიერება გარემოში და მისთ.) ამ სახის ბუნებრივი მუტაგენები არასეისმურ რაიონებში მცირეა. მეორე მნიშვნელოვან ფაქტორად ინფექციური ვირუსული დაავადებები სახელდება (იხ. §5.3).
საჭიროა აღინიშნოს, რომ პრიმატების ფილოგენიაში დაფიქსირებულია 390 ქრომოსომული მუტაცია, მათგან 120 პერიცენტრული, 20 კი პარაცენტრული ინვერსიაა. ადამიანი კარიოტიპით (2N=46) განსხვავდება ახლომონათესავე ფორმებისგან (გორილა, შიმპანზე, 2N=48). ყველა საფუძველი არსებობს ვივარაუდოთ, რომ ადამიანის, ავსტრალოპითეკისა და ადამიანის მსგავსი მაიმუნების საერთო წინაპარს 2N=48 კარიოტიპი გააჩნდა. ანთროპოგენეზის უაღრესად რთულ და მრავალფაქტორიან პროცესში ქრომოსომულმა მუტაციებმა მნიშვნელოვანი წვლილი
შეიტანა, კერძოდ, რეპროდუქციული იზოლაციის ფორმირებაში. ამ მექანიზმით ადამიანის წინაპარი გაემიჯნა ახლო მონათესავე ფორმებს. ამ უკანასკნელთ შეუნარჩუნდათ 48 ქრომოსომიანი კარიოტიპი. 48 ქრომოსომიანი კარიოტიპის 46 ქრომოსომიანად გარდაქმნის პროცესი სეისმურად აქტიურ ზონაში - დიდი აფრიკის რიფტში განხორციელდა (ვორონცოვი, 2004) (იხ. სურ. 12.11).
ხაზგასმით უნდა აღინიშნოს, რომ ქრომოსომული მუტაციებით იქმნება შემდგომი დივერგენციის წინაპირობა. სახეობათა წარმოქმნის პროცესი ყოველთვის გაწელილია დროში მაშინაც კი, როდესაც საუბარია „მყისიერ“ სახეობათა წარმოქმნაზე. ევოლუციის სინთეზური თეორიის თვალთახედვით მყისიერ სახეობათა წარმოქმნა მიმდინარეობს პოპულაციურ დონეზე და ათასობით თაობას მოიცავს. დუპლიკაციების სახით ბუნებრივ გადარჩევას მიეწოდება მასალა, რომლიდანაც ახალი გენები წარმოიშვება, ხოლო ინვერსიები და ტრანსლოკაციები ახალ სახეობრივ ფორმებში საიზოლაციო მექანიზმების ჩამოყალიბებას უზრუნველყოფენ.
სურ. 12.11. ადამიანისა (თეთრი ფერის ქრომოსომები) და გორილას (შავი ფერის ქრომოსომები) კარიოტიპების შედარება (ნ. დუბინინი, 1986).
|
|
14.8 12.8. სახეობათა ფილეტური წარმოქმნა |
▲ზევით დაბრუნება |
ფილეტურ სახეობათა წარმოქმნის დროს მიმდინარეობს საწყისი სახეობის თანდათანობითი გარდაქმნა ახალ სახეობად, პოპულაციაში თანდათანობით აღინიშნება სხვადასხვა მემკვიდრული ცვლილებები, რომლის მეშვეობითაც ხორციელდება გენეტიკური სტრუქტურის (გენოფონდის) ვექტორიზებული შეუქცევადი ცვლილება (დიფერენცირება). სახეობა როგორც ერთიანი, მთლიანი სუბსტანცია, იცვლება, რადგან ამ ცვლილების გარეშე ევოლუციური პროცესის მიმდინარეობა შეუძლებელია. ფილეტური სახეობათა წარმოქმნა (სინონიმებია: ცვლილება დროში; ცვლილება შთამომავლებში; სახეობათა გარდაქმნა) არ იწვევს გეოგრაფიული რასების ქვესახეობების ჩამოყალიბებას, შესაბამისად - სახეობათა რიცხობრივ ზრდას. ფილეტური ევოლუციის შედეგად წარმოიქმნება ერთი განუტოტავი ფილოგენეზური ხაზი. იგი შედგება დროში უწყვეტი ჯგუფების (პოპულაცია, ქვესახეობა, სახეობა) რიგებისგან. ცალკეული ჯგუფი წინარე ჯგუფებიდან იღებს სათავეს. ამრიგად, თაობათა უწყვეტ რიგში ერთი ჯგუფი მომდევნოსგან უმნიშვნელოდაა განსხვავებული, ხოლო საწყის და საბოლოო ჯგუფებს შორის მორფო-ფიზიოლოგიური განსხვავება იმდენად მკვეთრია, რომ ამ უკანასკნელს ახალ სახეობად მიიჩნევენ.
ფილეტური ევოლუციის დროს ახალი სახეობის გამოყოფა მხოლოდ იმ შემთხვევაშია შესაძლებელი, თუ უწყვეტი რიგის შემადგენელი ჯგუფების მორფოლოგიურ მაჩვენებლებს ერთიმეორეს შევადარებთ, ე.ი. განუტოტავ ფილოგენეზური ხაზის ცალკეულ მონაკვეთს ურთიერთს შევადარებთ. ამდენად ფილეტურ სახეობათა წარმოქმნის ანალიზი მხოლოდ პალეონტოლოგიურ მასალაზე დაყრდნობითაა შესაძლებელი. ამასთანავე ყოველთვის რჩება ალბათობა, რომ ევოლუციის ამა თუ იმ ეტაპზე ერთიანი ღერძი განიტოტოს. ამიტომაც „წმინდა სახით“ ფილეტური ევოლუციის გზა იდეალიზებული და გამარტივებული სახით ევოლუციის არსს გამოხატავს. გენეტიკური ცვლილება, რომელსაც ფილეტური ევოლუციას აძლევს დასაბამს, შეიძლება იყოს როგორც ავტოგენური (გამოწვეულია: მუტაციებით, რეკომბინაციით, გადარჩევით და სხვ.), ისე ალოგენური (სხვა სახეობათა გენების ჩართვით, გენეტიკური მასალის ლატერალური ანუ ჰორიზონტალური გადატანით). ყველა ევოლუციური ცვლილება ინტროგრესიითაა გამოწვეული. უნდა აღინიშნოს, რომ ფილეტური ევოლუციის დროს ცალკეულ სახეობათა შორის საზღვრის დადგენა შეუძლებელია - იგი ყოველთვის პირობითია.
|
|
14.9 12.9 სახეობათა წარმოქმნის ფორმები დროში |
▲ზევით დაბრუნება |
სახეობათა წარმოშობა დროის ხანგრძლივ პერიოდში მიმდინარე რთული პროცესია, რომელიც სხვადასხვა გზით ხორციელდება. ამჟამად გამოყოფენ დროში მიმდინარეობის მიხედვით სახეობათა წარმოქმნის ოთხ ფორმას: სტასიგენეზი, ანაგენეზი, კლადოგენეზი და სიმგენეზი (იხ. სურ. 12.12) ჩ.დარვინის მიერ ნაშრომში „სახეობათა წარმოშობა“, მოწოდებულია ევოლუციური განვითარების დივერგენციის სქემა (იხ. სურ. 12.13). მასში დემონსტრირებულია პირველი სამი გზა, ხოლო მათი აღმნიშვნელი ზემოთ დასახელებული ტერმინები შემოთავაზებულია ბ. რენშის (1954) და ჯ. ჰაქსლის (1957) მიერ.
სტასიგენეზი - არის პროგრესული ცვლილების გარეშე სახეობის (ან სხვა ტაქსონის) ხანგრძლივად შენარჩუნების პროცესი. სახეობის ისტორიული განვითარება არცერთი მიმართულებით (როგორც ანაგენეზის ისე კლადოგენეზის) არ მიმდინარეობს და არ მოქმედებს მამოძრავებელი გადარჩევა. სტასიგენეზის გზით ვითარდება ზოგიერთი ამჟამად მცხოვრები პერსისტენტული ფორმები: მოლუსკი-ნაუტილუსი, კიბოსნაირი - მახვილკუდები, ცელაკანთისნაირი თევზი - latimeria, პრიმიტიული reptilia - tuataria (hateria), ciano - და არქებაქტერიის მრავალი სახეობა. ეს არის იმ ფორმების არასრული ჩამონათვალი, რომლებიც მრავალი ათასი და მილიონი თაობის განმავლობაში თითქმის უცვლელი დარჩა. ზოგჯერ პრესაში ამ ფორმების მიმართ გვხვდება გამონათქვამი „ევოლუციის გარეშე“ ან „შეჩერდა ევოლუცია“. ეს სინამდვილეს არ შეესაბამება. მოქმედებს ბუნებრივი გადარჩევის მასტაბილიზებელი ფორმა. ამ გადარჩევის ზეწოლის მოხსნა იწვევს პოპულაციაში ნაირგვარ მუტაციათა დაგროვებას. ჩვეულებრივ კი მათ მასტაბილიზებელი გადარჩევა ცხრილავს (იხ. § 9.4 და 11.4).
სურ. 12.12. სახეობათა ფილეტური წარმოქმნის (დროში) ძირითადი ფორმები (ნ. ვორონცოვი, 2004).
სურ. 12.13. ევოლუციური განვითარების პროცესში სახეობათა და ნებისმიერი სხვა ჯგუფების დივერგენციის სქემა (მოწოდებულია ჩ. დარვინის მიერ შრომაში „სახეობათა წარმოშობა“).
ანაგენეზი არის ჯგუფის პროგრესული განვითარება, რომლის დროსაც მიმდინარეობს ძველი ტაქსონის თანდათანობითი გარდაქმნა ახალ ტაქსონად, შვილეულ ტაქსონებად დივერგირების (gantotvis) გარეშე. ანაგენეზის კლასიკურ მაგალითს ცხენის, სპილოს, მარტორქას ევოლუციური ჩამოყალიბება წარმოადგენს. ასევე ადამიანის წინაპრის ავსტრალოპითეკებიდან - მარჯვე ადამიანის (Homo habilis), და ზეამართული ადამიანის (H. erectus) გზით გონიერი ადამიანის (H. sapiens) ფორმირება ანაგენეზით განხორციელდა. ანაგენეზი ფილეტური ევოლუციის გზით მიიღწევა (იხ. § 12.8). იგი ბუნებრივი გადარჩევის მამოძრავებელ ფორმასთანაა დაკავშირებული.
სურ. 12.14. გალაპაგოსის არქიპელაგსა და კუნძულ კოქსზე გავრცელებული დარვინისეული მთიულების (Geospizinae) დივერგენცია (დ. ლეკი, 1949).
კლადოგენეზი არის ჯგუფის დივერგენციული განვითარება, როდესაც საწყისი ტაქსონი იტოტება რამდენიმე მონათესავე ჯგუფად. მხოლოდ კლადოგენეზზე (ბერძ. Klados - ტოტი) ვრცელდება ტერმინები - დივერგენციული ევოლუცია, ადაპტური რადიაცია. იგი სახეობათწარმოქმნის ყველაზე ფართოდ გავრცელებული გზაა. მისი მეშვეობით ყალიბდება, ადაპტური რადიაცია. კლადოგენეზის პოპულარული მაგალითია ავსტრალიური ჩანთოსნების დივერგენცია, რის შედეგადაც ამ კონტინენტზე ჩამოყალიბდა ჩანთოსანი მგელი, ჩანთოსანი კვერნა, ჩანთოსანი თაგვები, ჩანთოსანი თხუნელა, ჩანთოსანი მაჩვი და მისთ.
კლადოგენეზის მეორე მნიშვნელოვანი არგუმენტია საწყისი სახეობიდან არქიპელაგის სხვადასხვა კუნძულზე ჩამოყალიბებული სახეობები. ვულკანური წარმოშობის გალაპაგოსის კუნძულებზე გავრცელებულია დარვინისეული მთიულების სახეობები. მათ კონტინენტიდან შემთხვევით მოხვედრილმა ფრინველთა საკმაოდ მცირე ჯგუფმა დაუდო სათავე. ამჟამად არქიპელაგზე გავრცელებულ ფრინველებს ცალკე ოჯახად გამოყოფენ, რომელშიც 4 გვარი და 14 სახეობაა. მათი დივერგირება კვებითმა სპეციალიზაციამ და ნისკარტის სათანადო ცვლილებამ გამოიწვია (იხ. სურ. 12.14). ამავე გზით არის ჩამოყალიბებული ჰავაის კუნძულებზე ფრინველთა ოჯახი, ჰავაიური მეყვავილეები (Drepanididae), რომელშიც 19 გვარი და 40 ენდემური სახეობაა. კლადოგენეზი მამოძრავებელი და დიზრუპტული გადარჩევის ერთდროულ ან თანამიმდევნო მოქმედებასთანაა დაკავშირებული.
სიმგენეზი (სინთეზოგენეზი) - არის სახეობათა წარმოქმნის ფორმა დროში, როდესაც დამოუკიდებელი (რეპროდუქციულად იზოლირებული) სახეობების გენომების შერწყმით ყალიბდება ახალი სახეობა (ტერმინი ეკუთვნის ნ. ვორონცოვს). მასში გამოყოფენ ორ ფორმას ჰიბრიდოგენეზს და სიმბიოგენეზს. სიმგენეზი პრინციპულად განსხვავდება ანაგენეზისა და კლადოგენეზისგან. სიმგენეზში არ მოქმედებს ევოლუციის ელემენტარული მოვლენა - პოპულაციის გენეტიკური შემადგენლობის ცვლილება. ევოლუციას იწვევს და წარმართავს განსხვავებულ გენომთა გაერთიანება (შერწყმა), ანაგენეზსა და კლადოგენეზში კი ბუნებრივი გადარჩევა, იზოლაცია და გენთა დრეიფი.
სიმგენეზში მნიშვნელოვან როლს ასრულებს ბუნებრივი გადარჩევა. მისი მოქმედებით არაადაპტური (დაბალსიცოცხლისუნარიანი) ჰიბრიდული კომბინაციის გენომების ელიმინაცია ხორციელდება. განსხვავებული სახეობების შეჯვარებითა და შემდგომი პოლიპლოიდიით (ან აპომიქსისით) ჩამოყალიბებული ინდივიდები რეპროდუქციულად იზოლირდებიან საწყისი ფორმებისგან. ამრიგად სიმგენეზი განიხილება როგორც მაკროევოლუციური პროცესი, რომელიც არ არის გამოწვეული მიკროევოლუციური პროცესებით.
ეუკარიოტული უჯრედის წარმოშობის თანამედროვე სიმბიოგენეზის კონცეფციის არსი შემდეგში მდგომარეობს: 1. დიდი ზომის ამებოიდური, ანაერობული პროკარიოტული უჯრედისა და მცირე ზომის აერობი პროკარიოტულ უჯრედთან სიმბიოზმა ამ უკანასკნელის მიტოქონდრიებად ტრანსფორმაცია გამოიწვია (იხ. სურ. 12.15) 2. აერობული მიტოქონდრიების მფლობელი პროკარიოტულ უჯრედისა და სპიროქეტების მსგავსი პროკარიოტული უჯრედის სიმბიოზით შოლტები, კინეტოსომა და ცენტროსომა ჩამოყალიბდა. ამ ორგანოიდებით უჯრედში გენეტიკური მასალა ბირთვად დიფერენცირდა ე.ი. ციტოპლაზმა და ბირთვი ჩამოყალიბდა, რაც ამავე დროს ეკარიოტულ უჯრედს ნიშნავს. ასეთმა უჯრედმა დასაბამი მისცა ცხოველთა და სოკოების სამეფოს. 3. სიმბიოგენეზის კიდევ ერთი აქტით განხორციელდა ეუკარიოტულ უჯრედში პროკარიოტული უჯრედის ციანობაქტერიების ჩანერგვა, რომელიც პლასტიდებად დიფერენცირდა. აქედან სათავეს იღებს მცენარეთა სამეფო. ამ კონცეფციის სარწმუნო არგუმენტია პლასტიდებსა და მიტოქონდრიებში წრიული ბაქტერიების დნმ-ის მსგავსი სტრუქტურის თანაპოვნიერება.
სურ. 12.15. სიმბიოგენეზის გზით ძირითადი ეუკარიოტული ჯგუფების ჩამოყალიბების თანმიმდევრული ეტაპები (ლ.კაიდანოვი, 1996).
|
|
14.10 12.10. გენეტიკური მასალის ჰორიზონტული (ლატერალური) გადატანა |
▲ზევით დაბრუნება |
გენომის რეორგანიზაციის ერთ-ერთი გზა არის ერთი სახეობიდან მეორეში დნმ-ის მონაკვეთის (ბუნებრივია გენის ან შეჭიდულ გენთა ჯგუფის) გადატანა და ინტეგრაცია. როგორც უკვე აღინიშნა, ყოველი სახეობა დახშულ გენეტიკურ სისტემას წარმოადგენს. მას მოეპოვება საიზოლაციო მექანიზმები, რომელიც უზრუნველყოფს გენოფონდის დაცვას უცხო გენთა ნაკადისაგან. ამჟამად მეცნიერება ფლობს არგუმენტებს, რომელიც მიუთითებს, რომ გენოფონდი სრულყოფილად (აბსოლუტურად) არ არის დაცული უცხო გენთა ნაკადისაგან. მაშასადამე, საიზოლაციო მექანიზმები მხოლოდ ნაწილობრივ იცავს გენოფონდს უცხო გენების შეღწევისაგან.
ჯერ კიდევ გასული საუკუნის 50-იან წლებში დაადგინეს ტრანსფორმაციის (იზოლირებული დნმ-ის გადატანა ერთი უჯრედიდან მეორეში) და ტრანსდუქციის (გენების გადატანა ერთი ბაქტერიული უჯრედიდან სხვა უჯრედში ბაქტერიოფაგის მეშვეობით) გამომწვევი მიზეზი. გაირკვა, რომ ტრანსფორმაციისა და ტრანსდუქციის ფენომენი, რომლითაც რეკომინაციის პროცესი მიიღწევა, ეგზოტიკური შემთხვევა როდია, იგი ბუნებრივი პროცესია.
ბოლო წლებში გენური ინჟინერიის განვითარებასთან დაკავშირებით ფართოდ გამოიყენება პლაზმიდური ან ვექტორული ტრანსფორმაცია, კერძოდ ბუნებრივ ან ხელოვნურ პლაზმიდებში ინტეგრირებული გენები შეჰყავთ ბაქტერიულ ან ეუკარიოტულ უჯრედში. ამ მეთოდით მცენარეთა მრავალი ტრანსგენული ჯიშია მიღებული.
ჯერ კიდევ 1957 წელს კ.უოდინგტონმა ჯეროვნად შეაფასა ტრანსდუქციის ევოლუციური მნიშვნელობა. მან ივარაუდა მსგავსი მექანიზმების მეშვეობით გენთა ნაკადის მიმდინარეობა პროკარიოტულ და ეუკარიოტულ გენეტიკურ სისტემებს შორის. ბოლო პერიოდამდე ამ ფენომენს ევოლუციონისტები გვერდს უვლიდნენ. გენომის გარდაქმნა იმ მეთოდებით, რომელსაც ამჟამად გენურ ინჟინერიაში იყენებენ, ბუნებასაც შეეძლო ესარგებლა 3,5 მლრდ. წლიანი ევოლუციური განვითარების მანძილზე. კონკრეტულ შემთხვევაში, რამდენად წარმატებულად მოხდებოდა უკარიოტულ უჯრედში უცხო გენომის (სახეობის, გვარის, რიგის) მონაკვეთის ჩართვა და შენარჩუნება არ არის არსებითი. პრინციპულად მნიშვნელოვანია ის, რომ ტრანსდუქციის მექანიზმებით განხორციელებულიყო ახალი გენომის სინთეზი (ნ. ვორონცოვი, 2004).
სხვადასხვა სამეფოში გაერთიანებულ ინდივიდებს შორის ჰორიზონტული გადატანის სანიმუშო მაგალითი გამოავლინეს 1981 წელს ჯ.მარტინმა და ი.ფრიდოვიჩმა. ვერცხლმუცელა თევზის (Leiogna thidae, ოჯახი Leiognathidae) სპეციალურ ჯირკვლებში ბინადრობს სიმბიონტური ბიოლუმინესცენტური ბაქტერია Photobacter leignathi. იგი თევზში მუცლის ნათებას იწვევს. ყველა აერობულ ორგანიზმს მოეპოვება ფერმენტი მეტალოპროტეიდი - სუპეროქსიდდისმუტაზა. ფერმენტი დამცველობითი ფუნქციისა. იგი აუვნებელყოფს პეროქსიდულ რადიკალებს. ფოტობაქტერია, სხვა პროკარიოტების მსგავსად, შეიცავს სპეციფიკურ ფერმენტ Fe - superoqsiddismutazas. ამავე ბაქტერიას დამატებით მოეპოვება მეორე სახის ფერმენტიც Cu-Zn - superoqsiddismutaza. ეს უკანასკნელი გვხვდება მხოლოდ ეუკარიოტულ ორგანიზმებში, მათ შორის თევზებში. ვერცხლმუცელა თევზისა და ფოტობაქტერიის Cu-Zn - superoqsiddismutazas ამინოუმჟავური შემადგენლობის ანალიზმა ცხადყო მათი მსგავსი აგებულება. ბუნებრივია ბაქტერიამ ფერმენტის მაკონტროლირებელი გენი თევზის გენომიდან ტრანსდუქციის მექანიზმის მეშვეობით შეიძინა.
გამოვლენილია პროკარიოტებისათვის დამახასიათებელი Fe - სუპეროქსიდდისმუტაზის გენის ჰორიზონტული გადატანის ფაქტი ბაქტერიიდან (ან წყალმცენარიდან, მოეპოვება ქლოროპლასტის დნმ-ს) უმაღლეს მცენარეებში. შეისწავლეს გვიმრანაირების, შიშველთესლოვანების და ყვავილოვანი მცენარეების 43 ოჯახი. სამ ოჯახს Gინკოაცეაე (შიშველთესლოვანი) და Nymphacea (დუმფარასებრი), Cruciferae (ჯვაროსანნი; ორივე ფარულთესლოვანი) მოეპოვებათ ფერმენტი Fe - speroqsiddismutaza. სამივე ოჯახის ფერმენტები მკვეთრად განსხვავდებიან ერთმანეთისგან. ვარაუდობენ, რომ სხვადასხვა ბაქტერიიდან ან წყალმცენარიდან ტრანსდუქცირებული გენები დამოუკიდებელი გზით აღმოჩნდა ამ ოჯახებში.
ჯვაროსანი მცენარისა და ბაქტერიის Agrobacterium tumefaciens ურთიერთდამოკიდებულება საყოველთაოდ არის ცნობილი. მცენარის ფესვებსა და ბაქტერიებს შორის იმდენად მჭიდრო კონტაქტი მყარდება, რომ ბაქტერიის თი პლაზმიდან ფრაგმენტი თ-დნმ ინტეგრირდება მცენარის გენომში. მცენარეში მოხვედრილი თ-ფრაგმენტით იცვლება ზრდისა და დიფერენცირების პროცესი. მას სიმსივნური უჯრედები უყალიბდება, რომელიც იწყებს ამინომჟავების (ლიზინი, ჰისტიდინი, ორნიტინი ან არგინინი) ანალოგების - ოპინების ინტენსიურ სინთეზს. ეს ნაერთები ბაქტერიის დამატებითი აზოტოვანიწყაროა. ბაქტერიები ინტენსიურად იწყებენ გამრავლებას. თ-ფრაგმენტი ფერმენტ ლიზოპინდეჰიდროგენაზას ინდუცირებს. ვარაუდობენ, რომ ბაქტერიამ ეს გენი (აღენიშნებათ მოლუსკებს) ტრანსდუქციის შედეგად ეუკარიოტებისგან შეიძინა.
პალეობოტანიკოსები თვლიან, რომ ავსტრალიაში გავრცელებულ მცენარეთა სხვადასხვა ოჯახში ევკალიპტისებური ფორმის ფოთლის პარალელური ჩამოყალიბება (სხვა ქვეყნებში ფორმა სხვაგვარია) ვირუსული ტრანსდუქციის შედეგია. იგი წარმოადგენს ერთგვარ „მოდას“ მუტაციაზე (იხ. §5.2).
როგორც ირკვევა, ვირუსული ტრანსდუქცია (ჰორიზონტული გადატანა) გარკვეულ როლს ასრულებს სახეობათა ფორმირების პროცესში. მან გაართულა ბადებრივი (რეტიკულური) ევოლუცია.
|
|
15 § 13. მოლეკულური ევოლუციის საკითხები |
▲ზევით დაბრუნება |
|
|
15.1 გენეტიკური კოდის ევოლუცია |
▲ზევით დაბრუნება |
დნმ არის უმთავრესი ბიოლოგიური მოლეკულა, რომელიც სიცოცხლისა და ევოლუციის საიდუმლოს ფლობს. სიცოცხლის ორგანიზაციის ნებისმიერ დონეზე (უჯრედული, ორგანიზმული, პოპულაციურ-სახეობრივი.) ევოლუციურ გარდაქმნას, მოლეკულურ-გენეტიკურ დონეზე (დნმ-ის მოლეკულებში) მომხდარი ცვლილებები განაპირობებს. ცოცხალი სისტემის ევოლუცია, რომელიც დაახლოებით 3.5 მლრდ წელს მოიცავს, არსებითად დნმ-ის ევოლუირების პროცესს წარმოადგენს. ცოცხალი სისტემების გამრავლებას დნმ უზრუნველყოფს. იგი ინსტრუქტირებს ორგანიზმს, თუ როგორ განვითარდეს და იფუნქციოს. ეს პროცესი ხორციელდება დნმ-ის მოლეკულაში კოდირებული ინფორმაციის მიხედვით.
გენეტიკური კოდის გაშიფვრისთანავე მეცნიერებმა შენიშნეს მისი ერთ-ერთი მნიშვნელოვანი თვისება-უნივერსალობა. როგორც მარტივ (ვირუსები, ბაქტერიები) ისე რთულ (ფარულთესლოვანები, ძუძუმწოვრები, მათ შორის ადამიანი) ორგანიზმებში კოდი ერთიანია. თავდაპირველად გაბატონებული იყო შეხედულება, რომლის მიხედვითაც გენეტიკური კოდი უცვლელი დარჩა მას შემდეგ, რაც საერთო პროკარიოტი წინაპარი სამ დომენად (არქეები, ეუბაქტერიები, ეუკარიოტები) დივერგირდა (კრიკი, 1957). ამჟამად მეცნიერება ფლობს მასალას, რომლის თანახმადაც გენეტიკური კოდის ჩამოყალიბებაში გადამწყვეტი როლი ბუნებრივმა გადარჩევამ შეასრულა (ფრილენდი, ჰერსტი, 2004).
გენეტიკური კოდის მეორე მნიშვნელოვანი თვისება არის კოდონთა სიჭარბე (კოდი გადაგვარებულია). არსებობს 64 კოდონი, მათგან სამი (UAG, UGA, UAA) „სტოპ-სიგნალია“ (კოდირებს ტერმინაციას). დანარჩენი 61 კოდონი 20 ბლოკად იყოფა. ბლოკში გაერთიანებული კოდონები ერთსა და იმავე ამინომჟავას აკოდირებს. მხოლოდ ორ ბლოკშია თითო ტრიპლეტი (მეთიონინი, თრეონინი), დანარჩენ ბლოკებში ორი და მეტი კოდონია თავმოყრილი (იხ. სურ. 13.1). ამრიგად, გენეტიკურ კოდს მოეპოვება სინონიმური კოდონები. ერთსა და იმავე ბლოკში გაერთიანებული კოდონები ძირითადად მესამე პოზიციით განსხვავდებიან (გამონაკლისია ექვსწევრიანი ბლოკები). ბლოკში თავმოყრილი კოდონები მნიშვნელოვან თვისებებს ფლობენ. მესამე პოზიციის სპონტანური მუტაცია (სეიმსენსი) ინფორმაციას არ ცვლის. როდესაც სხვადასხვა ბლოკებში გაერთიანებულ კოდონებს პირველ ორ პოზიციაში მოეპოვებათ ერთნაირი ფუძეები, მესამე პოზიციის შეცვლა ასევე არსებით გავლენას არ ახდენს ცილის სივრცით კონფორმაციაზე. ამ შემთხვევაში ცილის მოლეკულაში საწყისი ამინომჟავის შეცვლა მსგავსი თვისებების (წარმოადგენს წინამორბედს ან ერთი მეორის პროდუქტს) ამინომჟავით ხდება. ამ სახის მუტაციები სელექციურად ნეიტრალურია (იხ. § 14.1).
სურ. 13.1. გენეტიკური კოდი, ცალკეული ამინომჟავის მადეტირმინირებელი კოდონების რიცხვი და შემადგენლობა.
ცოცხალი სისტემის ძირითად ნაწილში ფუნქციონირებს სტანდარტული ე.წ. კანონიკური კოდი. ამჟამად გამოვლენილია კოდის 16 ვარიანტი, რომელიც სხვადასხვა ფილოგენეზური რიგების წარმომადგენლებში გვხვდება. კოდირების პრინციპი ყველა მათგანში უცვლელია. თითოეულ ამინომჟავას სამი ნუკლეოტიდი - ტრიპლეტი შეესაბამება. ასე მაგალითად, მრავალ წყალმცენარეში, მათ შორის აცეტაბულარიაში სტანდარტული ტერმინაციის მიმანიშნებელი UAG და UAA ტრიპლეტებით ამინომჟავა გლიცინი კოდირდება. ზოგიერთ სოკოში, მაგალითად, გვარ ჩანდიდა-ში გაერთიანებულ მრავალ სახეობაში ტ-რნმ-ის CUG კოდონი სტანდარტული ამინომჟავა ლეიცინისაგან განსხვავებით სერინს ინტერპრეტირებს. საფუარებში, კერძოდ გვარ შაცცჰარომყცეს-ის ზოგიერთი სახეობის მიტოქონდრიაში ტრიპლეტები CUU, CUC, CUA დაCUG ლეიცინის ნაცვლად თრეონინს განსაზღვრავს. ძუძუმწოვრების მიტოქონდრიაში AGA და AGG კოდონები ტერმინაციის სიგნალია, AUU, AUC, AUA და AUG მაინიცირებელი ტრიპლეტები, ხოლო AUA იზოლეიცინის ნაცვლად მეთიონინს აკოდირებს. ამდაგვარი მაგალითების დასახელება კიდევ შეიძლება, რაც მიუთითებს, რომ ისტორიული განვითარების პროცესში გენეტიკური კოდი ევოლუირდება.
ცოცხალ სისტემათა სამივე დომენში გაერთიანებულ წარმომადგენლებში ზოგჯერ სტანდარტული „სტოპ-სიგნალი“ UGA იკითხება, როგორც არასტანდარტული (სტანდარტულად ითვლება კანონიკური კოდით დეტერმინირებული 20 სახის ამინომჟავა) დამატებითი 21-ე ამინომჟავა სელენოცისტეინი. ორ დომენში (არქეები, ეუბაქტერიები) გაერთიანებულ ზოგიერთ სახეობაში „სტოპ-კოდონი“ UAG სრულიად განსხვავებულ არაკანონიკურ 22-ე ამინომჟავას - პროლინს აკოდირებს.
გამოთქმულია მოსაზრება, რომ ადრეულ ეპოქაში თავდაპირველი ორგანიზმების ცილის შენებაში 20-ზე ნაკლები ამინომჟავა მონაწილეობდა. ევოლუციურად რთული ამინომჟავები გაცილებით გვიან, მარტივი ამინომჟავების ბიოქიმიური მოდიფიცირებით ჩამოყალიბდა. რაზედაც მიუთითებს შემდეგი ფაქტი; ზოგიერთ ბაქტერიაში ამინომჟავა გლუტამინი წინარე მეტაბოლიტ-გლუტამინის მჟავისაგან ჯერ კიდევ მაშინ წარმოიქმნება, როდესაც ეს უკანასკნელი ტრანსპორტულ რნმ-ის კომპლექსთან არის დაკავშიებული. რთულმა ამინომჟავებმა, მათი მარტივი წინამორბედების კუთვნილი ტ-რნმ და კოდონები მიითვისეს.
გასული საუკუნის 90-იან წლებში დაგროვდა მეცნიერული მონაცემები, რომელიც მიუთითებდა, რომ გენეტიკური კოდი არაა „გაქვავებული“ სისტემა; ასევე ტრიპლეტური კოდონებისა და ამინომჟავების შესაბამისობა შემთხვევითობაზე არ არის დაფუძნებული. გენეტიკური კოდი ევოლუციის პროცესში თანდათანობით შეიცვალა და გასრულყოფილდა. მემკვიდრულ აპარატში სპონტანურად ინდუცირდება სხვადასხვა სახის დარღვევები. გენებში მომხდარი ცვლილებები მუტაციებად რეალიზდება (მაგრამ არა ყოველთვის), ხოლო ტრანსლაციის პროცესში დაშვებული შეცდომები მოდიფიკაციურ ცვალებადობას აპირობებს. ეს უკანასკნელი ზოგჯერ მორფოზებად ან გენოკოპიებად ყალიბდება. ზემოთ განხილული თვისებების გამო კოდის სტრუქტურის ცვლილება (სეიმსენსის ტიპის მუტაციები) ინფორმაციას არ ცვლის. ზოგჯერ ცილის მოლეკულაში ერთი ამინომჟავა იცვლება იმავე ჰიდროფობური თვისებების ამინომჟავით. ამ შემთხვევაში ცილის სივრცითი კონფორმაცია უცვლელი რჩება. კანონიკური კოდით მინიმუმამდე მცირდება ინდუცირებულ მუტაციათა რაოდენობა, რითაც ორგანიზმი დაცულია საზიანო მუტაციების მავნე ზემოქმედებისგან. მაშასადამე, კანონიკური კოდი მეტად სიცოცხლისუნარიანია, ის ინდუცირებული ცვლილებების მინიმიზაციის უნარს ფლობს.
ვარაუდობენ, რომ განვითარების ადრეულ ეტაპზე ცოცხალ სისტემებში რამდენიმე ალტერნატიული კოდური სისტემა ფუნქციონირებდა. ამ მოსაზრების სარწმუნო არგუმენტია ამჟამად გენეტიკური კოდის 16 ვარიანტის არსებობა. კანონიკურმა კოდურმა სისტემამ კონკურენციაში (არსებობისათვის ბრძოლა) უპირატესობა მოიპოვა. მას სხვებისაგან განსხვავებით მოეპოვებოდა არსებითი უპირატესობა - როგორც გენში, ისე ტრანსლაციის პროცესში ინდუცირებული შეცდომების მინიმუმამდე დაყვანის უნარი (ე.წ. მინიმიზაცია). იგი მიიღწევა როგორც სინონიმურ კოდონთა დაჯგუფებებით, ისე იმ კოდონებით, რომლებიც მსგავსი ბიოქიმიური თვისებების ამინომჟავებს კოდირებენ. მინიმიზაციის უნარი გაცილებით მნიშვნელოვანია, ვიდრე დაშვებულ შეცდომებზე კონტროლი.
კანონიკური კოდი არის ბუნებრივი გადარჩევის პროდუქტი. იგი წარმოადგენს ეფექტურ სისტემას რომელმაც საგრძნობლად გაზარდა ევოლუციის სიჩქარე. კონკრეტულ გენში ინდუცირებული შეცდომების მინიმიზირება უზრუნველყოფს ცილის მოლეკულის სრულქმნას. ამდაგვარი კოდის მფლობელ ორგანიზმებს დიდი უპირატესობა გააჩნდათ არსებობისათვის ბრძოლაში სხვებთან შედარებით, რომელთაც იგი არ მოეპოვებოდათ.
|
|
15.2 გენომის ზომის ევოლუცია |
▲ზევით დაბრუნება |
ევოლუციის პროცესში იცვლებოდა როგორც დნმ-ის ნუკლეოტიდური შემადგენლობა, ისე მისი საერთო რაოდენობა. ვარაუდობენ, რომ პირველ ორგანიზმებს, რომლებმაც დასაბამი მისცეს დნმ-ის შემცველ ცოცხალ არსებებს რამდენიმე გენი გააჩნდათ. ბუნებრივია მათში დნმ-ის რაოდენობა ძალზე მცირე იყო (აიალა, კაიგერი, 1988). დღეს არსებული ევოლუციის სხვადასხვა საფეხურზე მდგომი ორგანიზმები დნმ-ის რაოდენობის მიხედვით მკვეთრად განსხვავდებიან. ამ ნიშნის მიხედვით უჯრედული ორგანიზაციის მფლობელი ორგანიზმები ოთხ კლასად შეიძლება დაიყოს. პირველ კლასში აერთიანებენ ბაქტერიებს. მათ უჯრედში საშუალოდ 4თ106 ნუკლეოტიდური წყვილი (ნ.წ.) გააჩნიათ. მეორე კლასში ათავსებენ სოკოთა სამეფოს წარმომადგენლებს. ბაქტერიებთან შედარებით მათი უჯრედი 10-ჯერ მეტ დნმ-ს შეიცავს. სოკოების ყოველ ერთ უჯრედზე დაახლოებით 4თ107 ნ.წ. მოდის. დნმ-ის საერთო რაოდენობის მომატება გენომში ძირითადად განმეორებადი ნუკლოტიდური თანმიმდევრობის ხარჯზეა გაზრდილი. უნდა აღინიშნოს, რომ სოკოებს გაცილებით მეტი სტრუქტურული გენი გააჩნიათ ვიდრე ბაქტერიებს. ამასთანავე მათი გენები მეტ მსგავსებას ავლენენ ბაქტერიების გენებთან ვიდრე უმაღლეს ეუკარიოტებთან (ზახაროვი, 1984).
სურ. 13.2. უჯრედში დნმ-ის შემცველობის მიხედვით ორგანიზმთა კლასიფიკაცია. დნმ-ის რაოდენობა მოცემულია წონით ერთეულში (1პგ=10-12 გ.) და ნუკლეოტიდური წყვილების რაოდენობით (ფ. აიალა, ჯ. კაიგერი, 1988).
მესამე კლასში თავს იყრის მცენარეთა და ცხოველთა სახეობების დიდი უმრავლესობა. ამ კლასის წარმომადგენელთა ერთ უჯრედზე საშუალოდ 2თ109 ნ.წ. მოდის. სოკოებისგან განსხვავებით ისინი დიდი რაოდენობით ფლობენ დნმ-ის, როგორც უნიკალურ ისე განმეორებად ნუკლოეტიდურ თანამიმდევრობებს. ამ კლასის წარმომადგენლებს გამოუმუშავდათ ეფექტური სისტემა, რომელიც გენომში დნმ-ის დაგროვებას აპირობებს. მეოთხე კლასში შიშველთესლოვან და ყვავილოვან მცენარეთა მნიშვნელოვანი რაოდენობის სახეობებია გაერთიანებული. ცხოველებიდან ამ კლასში იმყოფება სალამანდრები და უძველესი თევზები. მათ ერთ უჯრედზე გადაანგარიშებით 1010ნ.წ. გააჩნიათ (იხ. სურ. 13.2).
ევოლუციის პროცესში გენომში დნმ-ის რაოდენობის თანდათანობითი გაზრდა ყველა ორგანიზმში ხდებოდა მარტივი პროკარიოტებიდან მოყოლებული უმაღლესი ეუკარიოტების ჩათვლით. რთული ორგანიზაციის ორგანიზმები საჭიროებენ გაცილებით მეტ დნმ-ს. მათში გენომის ევოლუირებისას დნმ-ის რაოდენობის გაზრდა ხორციელდებოდა როგორც პოლიპლიდიის, ისე ტანდემური დუპლიკაციის მეშვეობით.
|
|
15.3 გენის ევოლუცია |
▲ზევით დაბრუნება |
პროკარიოტებსა და ეუკარიოტებში, ასევე მათ ვირუსებში გენომის ანალიზის საფუძველზე გამოტანილია მნიშვნელოვანი დასკვნა, რომლის თანახმადაც გენეტიკური ინფორმაციის ელემენტარული ერთეული - გენი ცოცხალი სისტემის ისტორიული განვითარების პროცესში ევოლუირებდა.
ბაქტერიებში გენეტიკური მასალა ერთ ქრომოსომადაა ორგანიზებული და წარმოდგენილია ოპერონებად (ტრანსკრიპციის მარეგულირებელი ერთეული). ოპერონში გაერთიანებული სტრუქტურული გენები ბაქტერიულ ქრომოსომაში გარკვეული წესით არიან განლაგებული. ისინი აკოდირებენ ფერმენტებს, რომლებიც თანამიმდევრულად სინთეზის ან დეგრადაციის მეტაბოლურ რეაქციებს წარმართავენ. სტრუქტურული გენების ექსპრესია საერთო მარეგულირებელი სისტემით კონტროლდება. მათი ჩართვა და გამორთვა კოორდინირებულად ხორციელდება. გენეტიკური მასალის ოპერონული ორგანიზაციის სანიმუშო მაგალითია სალმონელაში ჰისტიდინური ოპერონი. მასში ცხრა სტრუქტურული გენია გაერთიანებული, რომლებიც ჰისტიდინის ბიოსინთეზის თანამიმდევრულ რეაქციებს აკონტროლებენ. კარგად არის შესწავლილი E. coli-Si laqtozis operoni, რომელშიც გაერთიანებული სამი სტრუქტურული გენი რომელიც ლაქტოზის უტილიზაციის პროცესს წარმართავს და სხვ.
პროკარიოტებში ცალკეულ გენში წარმოდგენილი გენეტიკური ინფორმაცია მარტივად რეალიზდება. ტრანსკრიპტი (i-rnm) ერთიან ინფორმაციას ფლობს და მთლიანად ტრანსლირდება. უჯრედის მეტაბოლიზმში ბაქტერიული გენი წესისამებრ, მხოლოდ ერთ ფერმენტულ რეაქციას აკონტროლებს (გვხვდება გამონაკლისი, მაგალითად, I დნმ-პოლიმერაზა სამ ფერმენტულ რეაქციაში მონაწილეობს).
რთული ბაქტერიული ვირუსების (ბაქტერიოფაგები) გენის სტრუქტურა და გენეტიკური მასალის ორგანიზაცია ბაქტერიების გენომის მსგავსია. მაგალითად λ და თ-წყვილ ბაქტერიოფაგების დნმ-ის მოლეკულაში ხაზობრივად განლაგებული გენები ოპერონადაა ორგანიზებული. ბაქტერიის ინფიცირების შემდეგ რეგულატორული მექანიზმით ხორციელდება სტრუქტურული გენების ექსპრესია. იგი ფაგის ნორმალური განვითარებისა და მორფოგენეზისთვისაა საჭირო.
საკმაოდ მარტივადაა ორგანიზებული რნმ-ის შემცველი ბაქტერიოფაგების (R17, F2, MS2 და მისთ.) გენეტიკური აპარატი, მათი გენეტიკური მასალა რნმ-ის ერთი ჯაჭვისაგან შედგება. ერთ ჯაჭვიან დნმ - და რნმ - შემცველი ფაგებში გამოვლენილია გადახურული გენები (ერთი გენი მეორეშია ლოკალიზებული იხ. სურ. 13.3). ამ სახის გენების ევოლუცია შეუღლებულად მიმდინარეობს. მათი ევოლუირება რამდენადმე გართულებულია. იგი წარმოადგენს ე.წ. „საზღაურს ევოლუციისათვის“, კერძოდ ინფორმაციის ტევადობის გაზრდის ტენდენციისადმი. მსგავსი ფენომენი გამოვლენილია რთული ბაქტერიოფაგების, ბაქტერიებისა და ეუკარიოტების მიგრირებად ელემენტებში.
სურ. 13.3. ბაქტერიოფაგ φX174 გენეტიკური რუკა. ყურადღება მიაქციეთ, რომ B გენი A გენშია ლოკალიზებული, E გენი კი D - გენის შემადგენელი მონაკვეთია (ს. ინგე-ვეჩტომოვი, 1989).
ეუკარიოტებში ერთი და იმავე მეტაბოლიზმის გზის სხვადასხვა ეტაპის მაკონტროლებელი გენები ოპერონისმაგვარ დაჯგუფებას არ ქმნიან. ისინი, ჩვეულებრივ შემთხვევით არიან გაბნეული გენომში. ეუკარიოტებში ხშირად გვხვდება მულტიენზიმური კომპლექსის მაკონტროლებელი გენები ე.წ. გენი-კლასტერები. ეუკარიოტულ ორგანიზმებში გენი არის მოზაიკური ანუ ინტრონ-ექზონური სტრუქტურის. ამრიგად გენი შეიცავს ინფორმაციულ (ექზონი) და არაინფორმაციულ (ინტრონი) თანამიმდევრობებს.
ეუკარიოტთა ვირუსების გენეტიკური მასალის ორგანიზაცია ნაირგვარია. ამ ნიშნით ისინი ემსგავსებიან როგორც ბაქტერიული ვირუსების, ისე ეუკარიოტული უჯრედის გენომს. ბაქტერიოფაგების მსგავსად მათი გენომის ევოლუირება გენეტიკური მასალის ეფექტურად გამოყენების მიმართულებით ხორციელდებოდა. რის შედეგადაც ეუკარიოტთა ვირუსებს ჩამოუყალიბდათ გადახურული გენები. მაშასადამე, გადახურული გენების თანაპოვნიერება ვირუსების - მცირე გენომიანი სისტემების, დამახასიათებელი ნიშანია. იგი მნიშვნელოვან ადაპტაციას წარმოადგენს.
ვირუსების გამრავლება და ცხოველმყოფელობა დამოკიდებულია უჯრედ-მასპინძლის მეტაბოლიზმზე. ამის გამო მათ ჩამოუყალიბდათ გენეტიკური მასალის იმგვარივე ორგანიზაცია, რაც ეუკარიოტულ უჯრედს ახასიათებს. ეუკარიოტთა ვირუსის გენი ინტრონ-ექზონურ სტრუქტურას ფლობს. უჯრედ-მასპინძელთან მსგავსება გენეტიკური მასალის რეალიზაციაშიც შეინიშნება. რნმ - შემცველ ვირუსში სხვადასხვა ცილები, ტრანსლაციის შედეგად მიღებული - პირველადი ერთიანი პროდუქტის პროტეოლიტური გახლეჩვით მიიღება. ამდენად, მათი ი-რნმ შეიძლება განვიხილოთ როგორც გენი - კლასტერი.
ევოლუციის პროცესში გენომში ყალიბდება ახალი გენები. ისინი გენეტიკური მასალის გარდაქმნით მიიღება. ახალი გენის ფორმირების ერთ-ერთ გზას დუპლიკაცია წარმოადგენს. ახლად დუპლიცირებულ გენზე აღარ მოქმედებს ბუნებრივი გადარჩევის ზეწოლა. მასში ფიქსირდება სხვადასხვა სახის მუტაციები.
გენში მომხდარი გარდაქმნებით, კონტროლირებად ცილას ეცვლება აქტიური ცენტრი
ან ინვარიანტული უბანი რაც კონფორმაციას ცვლის. შეცვლილი ფერმენტი შეიძლება
სურ. 13.4. ჰემოგლობინის მაკოდირებელი გენების ევოლუცია ადამიანში. პრო-α - საწყისი ჰემოგლობინი, წრეები აღნიშნავს გენის დუპლიკაციას (ს. ინგე-ვეჩტომოვი, 1989).
ახალ სუბსტრატს მიესადაგოს. ბუნებრივი გადარჩევის მოქმედებით, რომელიც მასალად მცირე მუტაციებს იყენებს, მისადაგების პროცესი ღრმავდება და სრულყოფილდება. რის შედეგადაც ფორმირდება ჰომოლოგიურ ცილათა ოჯახი. მათი შემადგენელი წევრები განსხვავდებიან როგორც ფუნქციით, ისე პირველადი სტრუქტურით. გენთა დუპლიკაციითა და დივერგენციის გზით არის ჩამოყალიბებული ადამიანში ჰემოგლობინის მაკოდირებელი გენები (იხ. სურ. 13.4).
ეუკარიოტების გენომს მოეპოვება ნორმალური გენების ისეთი ასლები, რომელიც არ ფუნქციონირებს (მათზე ტრანსკრიპცია არ მიმდინარეობს). მათ არ გააჩნიათ ინტრონები და შეიცავენ სხვადასხვა სახის (ნონსენსი, ათვლის ჩარჩოს გადაადგილება) მუტაციებს. ამდაგვარ ასლებს ფსევდოგენებს უწოდებენ. მათი არსებობით საბუთდება გენომში დუპლიცირებული გენის ფიქსაციის შესაძლებლობა. ფსევდონები არის ის მასალა, რომლითგანაც ახალი გენები ფორმირდება.
აღწერილია ახალი გენების წარმოშობის მეორე გზაც. კერძოდ იგი ხორციელდება გენების ან მათი ნაწილების შერწყმით. აგრეთვე შესაძლოა გენის შემადგენელი დომენები დაიყოს და ახალი გენი ჩამოყალიბდეს. გენების ფორმირების ზემოთ აღწერილი პროცესი განსხვავებული თანმიმდევრობით ხორციელდება, რითაც იქმნება გენების ევოლუციის ისტორია.
თანამედროვე თვალთახედვით, წერტილოვანი მუტაციები (ტრანზიცია, ტრანსვერსია, ნუკლეოტიდის ჩართვა ან ამოვარდნა) გენის ევოლუციაში მეორეხარისხოვან როლს ასრულებენ. წერტილოვანი მუტაციებით ჰომოლოგიური ცილების განსაზღვრულ ნაწილში (ე.წ. კოვარიონში) ამინომჟავათა პოზიციის ცვლილება ფიქსირდება. ამ სახის მუტაციებით იცვლება ცილის პირველადი სტრუქტურა, ხოლო მისი სტერეოქიმიური აღნაგობა უცვლელი რჩება. ჰომოლოგიური ცილების თანდათანობითი დივერგენციით ცილების (შესაბამისად, მათი მაკოდირებელი გენების) ოჯახი ყალიბდება. სხვადასხვა ობიექტებში გამოვლენილია სტრუქტურულად და ფუნქციურად ჰომოლოგიური ცილები. მიღებული ფაქტობრივი მონაცემები დამაჯერებლად არის ინტერპრეტირებული ნეიტრალური ანუ არადარვინული ევოლუციის კონცეფციაში (იხ.: §14.1).
ამრიგად, ახალი გენები, წესისამებრ, გენეტიკური მასალის ლოკალური ცვლილებით, კერძოდ, გენის დუპლიკაციით, ასევე, გენების შერწყმით და მათი ნაწილებად დაყოფით წარმოიქმნება. ხატოვნად თუ ვიტყვით, ძველი სტრუქტურული გენების შემადგენელი ნაწილების (ცალკეული დომენის მაკოდირებელი უბნის) რეკომბინირებით ახალი გენები ყალიბდება. მრავალი გენის პირველადი სტრუქტურის შედარებისას გამოირკვა, რომ უმეტეს მათგანს ჰომოლოგიური უბნები მოეპოვება. ცხადია, ევოლუციამ მრავალჯერ გამოიყენა გენეტიკური ინფორმაციის მფლობელი ერთი და იგივე ბლოკები ახალი სტრუქტურული გენების მისაღებად. გამოვლენილი ფენომენი ბიოლოგიური ევოლუციის ბლოკური ანუ მოდულური პრინციპის სანიმუშო მაგალითია.
გენების ევოლუცია მჭიდროდაა დაკავშირებული გენეტიკური მასალის სივრცობრივ ორგანიზაციასა და რეგულაციის სისტემის ევოლუციასთან. რეგულაციის ოპერონული სისტემიდან ცალკეული გენის ავტონომიურ რეგულაციაზე გადასვლას თან ახლდა პრინციპულად ახალი მარეგულირებელი სისტემის ჩამოყალიბება. იგი განაპირობა ეუკარიოტული ტიპის უჯრედული სისტემის ფორმირებამ და მემკვიდრული მასალის ქრომატინად ორგანიზებამ. ეუკარიოტებში ჩამოყალიბდა სრულიად ახალი მარეგულირებელი ელემენტები: ენჰანსერები (გამაძლიერებლები), და საილენსერები (ჩამხშობები). ეს ნუკლეოტიდური თანამიმდევრობები საკმაო მანძილით არის (ასეული და ათასეული ნ.წ.) დაცილებული სტრუქტურული გენისაგან. ქრომატინის ლოკალური განლაგების ცვლილებით ისინი სტრუქტურული გენის ექსპრესიაზე (აძლირებენ ან ახშობენ) ახდენენ გავლენას.
განვითარების სხვადასხვა საფეხურზე მდგომ ორგანიზმებში გენეტიკური მასალის მოლეკულური ორგანიზაციის შესწავლისას გამოირკვა, რომ არა მარტო სხვადასხვა ინდივიდების, არამედ ერთი და იმავე ორგანიზმის გენები სტრუქტურით და ექსრესიის ხასიათით განსხვავდებიან. გენები ზოგჯერ ორგანიზებულია ოპერონებად (არ არის ზოგადი კანონზომიერება). გენი აკონტროლებს ერთ ან რამდენიმე ფერმენტულ რეაქციას, შეიძლება ჰქონდეს ან არ ჰქონდეს ინტრონები, გენები შეიძლება ხაზობრივად იყოს განლაგებული და ერთმანეთისაგან გენთაშორისი ინტერვალით გამიჯნული, ზოგჯერ ისინი ერთმანეთს ხურავენ. ტაქსონომიურად საკმაოდ დაშორებულ ორგანიზმებში (პროკარიოტები, ეუკარიოტები და მათი ვირუსები) გენის სტრუქტურისა და ფუნქციის შედარების საფუძველზე გამოვლენილია მისი ევოლუციის შემდეგი ტენდეციები:
1. გენების ავტონომიზაცია: ეუკარიოტების გენები გაცილებით მეტი ავტონომიურობით გამოირჩევა, ვიდრე პროკარიოტებისა. ეუკარიოტებს აღარ მოეპოვებათ ტრანსკრიპციის რეგულაციის ოპერონული სისტემა. ამასთანავე, რიბოსომებმა დაკარგეს i-rnm-ის ერთსა და იმავე მატრიცაზე განმეორებით ინიცირების უნარი. მათ ტრანსლირების წარმართვა არ შეუძლიათ მაშინაც კი, როდესაც კოდონ-ტერმინატორს მოსდევს კოდონი-ინიციატორი. გენის ავტონომიზაციას ცალკეული გენის დამოუკიდებელი და სრულყოფილი რეგულაციის სისტემის ჩამოყალიბება მოჰყვა. ქრომოსომული გარდაქმნებისა და ტრანსპოზიციის, ე.ი. ცალკეული გენის გადანაწილებით გენომის ევოლუირების სრულიად ახალი გზა გამოიკვეთა.
2. გენების ოლიგომერიზაცია: ეუკარიოტებში გენების ოლიგომერიზაცია „რთული“ ანუ კლასტერული გენების (cluster-gene) ჩამოყალიბებისა და ფართოდ გავრცელების ტენდენციაში აისახა. გენის გართულებისადმი მიდრეკილება უეკარიოტების პროგრესულ ევოლუციაში მკაფიოდაა გამოხატული. წესისამებრ, დამოუკიდებელი კონკრეტული ფრაგმენტული რეაქციის მაკონტროლირებელი რამდენიმე გენი ერთ კლასტერულ გენადაა გაერთიანებული. ეს უკანასკნელი რამდენიმე რეაქციაში მონაწილე ცილა-ფერმენტს აკოდირებს. მაგალითად, სოკოში His4 გენი აკოდირებს ფერმენტს, რომელსაც ჰისტიდინის ბიოსინთეზში სამი ფერმენტული აქტივობა აქვს, arom1 გენის პროდუქტი კი ხუთ სხვადასხვა ფერმენტულ რეაქციაში მონაწილეობს. დროზოფილაში pur3 გენი პურინის ბიოსინთეზის ორ განსხვავებულ რეაქციას აკონტროლებს. კლასტერული გენის კონტროლით სინთეზირებული მულტიენზემური კომპლექსით იზრდება გარკვეული ფერმენტების კონცენტრაცია. ასევე, მეტაბოლიზმის ეტაპები სწრაფად, ეფექტურად და კოორდინირებულად მიმდინარეობს.
3. მოზაიკური სტრუქტურის გენის ჩამოყალიბება: ინტრონ-ექზონური ანუ მოზაიკური სტრუქტურის გენები ეუკარიოტებისთვისაა დამახასიათებელი. ეუბაქტერიებს ის არა აქვთ (გვხვდება იშვიათი გამონაკლისი). ინტრონები მოეპოვებათ არქებაქტერიების გენებს. ამის საფუძველზე გამოთქმულია მოსაზრება, რომ არქებაქტერიები ორ ტოტად დივერგირდა - პროკარიოტებად და ეუკარიოტებად. ამ უკანასკნელმა შეინარჩუნა და მათში გასრულყოფილდა გენის მოზაიკური სტრუქტურა. პროკარიოტების გენებმა ევოლუციის პროცესში დაკარგეს მოზაიკური სტრუქტურა. ნობელის პრემიის ლაურეატი უ. ჰილბერტი მიიჩნევს, რომ ეუბაქტერიებმა გენის მოზაიკურ სტრუქტურასთან ერთად დაკარგეს არაჰომოლოგიური რეკომბინაციით ექზონების გადაადგილების ე.ი. ამ მოქნილი მექანიზმის გამოყენებით გენების ევოლუირების უნარი. ეუკარიოტებმა წარმატებულად გამოიყენეს ევოლუციის ზემოთ აღნიშნული გზა და პროგრესულ ევოლუციაში უმაღლეს დონეს მიაღწიეს. წესისამებრ, ექზონები ერთი და იმავე ცილის სხვა და სხვა დომენებს (ნახევრად ავტონომიურ უბნებს) აკოდირებს. სწორედ ამ მექანიზმს ეფუძნება ეუკარიოტებში გენების ოლიგომერიზაციის ტენდენცია. ავტონომიური სპლაისინგის მექანიზმი ადაპტური მოდიფიკაციის ჩამოყალიბების შესაძლებლობას განაპირობებს. ტრანსკრიპტიდან ინტრონების ამოჭრა ყოველთვის არ ხდება. ის გარემო პირობებზეა დამოკიდებული. ამ შემთხვევაში ყალიბდება მუტაცია - გენოკოპია. როდესაც გენოკოპიის მაინდუცირებელი გარემო პირობები ხანგრძლივადაა შენარჩუნებული, მაშინ სპლაისინგის ერთ-ერთი ვარიანტი მასტაბილიზებელი გადარჩევის მოქმედებით ფიქსირდება. ის ხელს უწყობს შეცვლილ გარემოში ორგანიზმის ადაპტირებას.
4. გენის სპეციალიზაცია: როგორც პროკარიოტული, ისე ეუკარიოტული უჯრედების ვირუსების ვიწრო სპეციალიზაციამ და პარაზიტიზმმა მათ გენომში გადახურული გენების ფორმირება გამოიწვია. ამ გზით მცირე გენომის (როგორც დნმ-ის, ისე რნმ-ის შემცველი ვირუსების) მფლობელ ფორმებს ინფორმაციული ტევადობა ეზრდებათ.
|
|
15.4 მოლეკულურ-ფილოგენეზური ანალიზი |
▲ზევით დაბრუნება |
ბოლო წლებში მოლეკულურ ბიოლოგიასა და გენეტიკაში მოპოვებული ექსპერიმენტული მონაცემების განზოგადებისა და ახალი თეორიული მეთოდების საფუძველზე ევოლუციური თეორია ინტენსიურად იცვლება და ვითარდება. სიცოცხლის ორგანიზაციის მოლეკულურ-გენეტიკურ დონეზე ევოლუციური პროცესი დაკავშირებულია ნუკლეოტიდების შემადგენლობის (დნმ-სა და რნმ-ში) და ამინომჟავების (ცილებში) ცვლილებებთან. ეს მოლეკულები გადამწყვეტ როლს ასრულებენ ცოცხალ სისტემებში მოლეკულურ-გენეტიკური დონის ორგანიზაციასა და შენარჩუნებაში. ნებისმიერი ორგანიზმის დნმ-ში ნუკლეოტიდთა თანმიმდევრობა არის მისი ევოლუციური ისტორიის შედეგი.
სხვადასხვა მაკრომოლეკულები განსხვავებული სიჩქარით ევოლუირებენ. დიდი კონსერვატულობით წესისამებრ გამოირჩევიან ის გენები (ბუნებრივია მათ მიერ კოდირებული რნმ და ცილები), რომლებიც ცოცხალი სისტემების ადრეულ ეტაპზე ჩამოყალიბებულ საბაზისო (უმნიშვნელოვანეს სასიცოცხლო პროცესებში მონაწილე) ფუნქციებს ასრულებენ. ეს პროცესები ნიშანდობლივია უმრავლესი (შეიძლება ითქვას ყველა) ცოცხალი ორგანიზმისთვის. მათ მიეკუთვნება რიბოსომული რნმ-ის გენები, ჰისტონებისა და C ციტოქრომის მაკონტროლირებელი გენები და სხვ. შედარებით ნაკლები კონსერვატულობით გამოირჩევიან ისეთი გენები და ცილები, რომლებიც მოეპოვებათ განვითარების უმაღლეს საფეხურზე მდგომ ტაქსონებში გაერთიანებულ სახეობებს. მაგალითად ხერხემლიან ცხოველებში გლობინი და მათი განმსაზღვრელი გენები. განსაკუთრებით ვარიაბილურია რნმ-შემცველი ვირუსების გენები და ცილები. ისინი მიზანმიმართულად იცვლებიან მასპინძლის იმუნურ სისტემასთან ბრძოლის პროცესში (მაგალითად, გრიპის ვირუსები, აივ-ადამიანის იმუნოდეფიციტის ვირუსები, ონკოვირუსები და მისთ.).
ფილოგენიის რეკონსტრუქცია და გენეტიკურ განსხვავებათა შეფასება ეფუძნება შემდეგ დებულებას: გენები და მათ მიერ კოდირებული ცილები ჰომოლოგიურია ე.ი. საერთო წინაპრიდან არიან წარმოშობილი. განასხვავებენ გენთა ჰომოლოგიის ორ ტიპს: ორთოლოგიურსა და პარალოგიურს. ორთოლოგიური გენი წარმოშობილია საწყისი გენიდან. იგი მოეპოვებოდა საწყის სახეობას, რომლიდანაც შესადარებელი (გასაანალიზებელი) სახეობები ჩამოყალიბდა. ამ ტიპის გენებს აკუთვნებენ: რიბოსომული რნმ-ის გენებს, C ციტოქრომის მაკონტროლირებელ გენს და სხვ. ორთოლოგიური გენების ევოლუცია ასახავს იმ სახეობათა ისტორიულ განვითარებას, რომლებსაც ისინი გენოფონდში მოეპოვებათ.
სურ. 13.5. გლობალური ფილოგენეზური ხე, რომელიც 16S-18S რიბოსომული რნმ-ის მაკოდირებელი გენების ანალიზის საფუძველზეა შედგენილი (კ. ვუზი და სხვ., 1991).
გლობალური ფილოგენეზური ხის სანიმუშო მაგალითია 16S-18S რიბოსომული რნმ-ის ანალიზის საფუძველზე აგებული დენდროგრამა. იგი მოიცავს ცოცხალი სამყაროს ყველა დიდ ტაქსონს. შედგენილია ამერიკელი მკვლევარების კ. ვუდისა და თანაავტორების მიერ (იხ. სურ. 13.5). მას გააჩნია სამი გლობალური განტოტვა. ამრიგად, ცოცხალი სამყარო (სიცოცხლის იმპერია) სამ დიდ დომენად ე.წ. ზესამეფოდ იყოფა: ეუბაქტერიები, არქებაქტერიები და ეუკარიოტები. ისინი თანაბარი დისტანციით არიან ერთიმეორისგან დაცილებული.
ამ გამოკვლევამდე მიაჩნდათ, რომ ეუბაქტერიები და არქებაქტერიები არიან საერთო ზესამეფოს Procariota-ს შემადგენელი კატეგორიები, რომლებიც მნიშვნელოვანი დისტანციით ემიჯნება მეორე ზესამეფოს Eucariota-ს.
სურ. 13.6. C ციტოქრომის ანალიზის საფუძველზე აგებული ფილოგენეზური ხე. ციფრები მიუთითებს მინიმალურ ნუკლეოტიდურ ცვლილებას ცალკეულ მუტაციური ფილოგენეზურ ტოტში (ვ. ფიტჩი, ე. მარგოლიაში, 1967).
ვუდის მიერ შემოთავაზებული ფილოგენეზური ხე ძირითადად 16შ რიბოსომული რნმ-ის ანალიზის საფუძველზეა აგებული. ამ სახის მაკრომოლეკულები კოდირდება როგორც ბირთვული, ისე ორგანოიდებში (მიტოქონდრიები, ქლოროპლასტები)
არსებული გენებით. სიმინდის (ძეა მაყს) რიბოსომეული რნმ-ის ბირთვული ფრაქცია, როგორც მოსალოდნელი იყო, ნათესაობას ეუკარიოტებთან ავლენს. მიტოქონდრიებისა და ქლოროპლასტების რნმ-ის ფრაქცია ეუბაქტერიების მონათესავე აღმოჩნდა. ეს მონაცემები „სიმბიონტური“ ჰიპოთეზის ყველაზე მნიშვნელოვანი არგუმენტია (იხ. §12.9) ამ ჰიპოთეზის მიხედვით მიტოქონდრიები ჩამოყალიბდა სიმბიონტური მეწამული ბაქტერიებისგან, ხოლო ქლოროპლასტები სხვადასხვა თავისუფლად მცხოვრები ორგანიზმებიდან, მათ შორის ციანობაქტერიებისგან (მალახოვი, 2004).
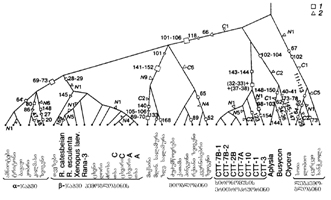
სურ. 13.7. გლობინების ზეოჯახის ფილოგენეზური ხე: 1. ფიქსირებული დელეციები. 2. ინსერცია. ციფრები მიუთითებს ცილაში მუტაციური ცვლილების ლოკალიზაციას (ვ. რატნერი, 2000).
როგორც უკვე აღვნიშნეთ, ორთოლოგიურ გენს მიეკუთვნება C ციტოქრომის მაკონტროლირებელი გენი. იგი ყველა ეუკარიოტული უჯრედის (სოკოები, მცენარეები, ცხოველები) მიტოქონდრიას მოეპოვება. უ.ფიტჩისა და ე. მარგოლიაშის მიერ მოწოდებულია C ციტოქრომის ანალიზის საფუძველზე აგებული ფილოგენეზური ხე (იხ. სურ. 13.6). მიღებული შედეგი ზოგადად ემთხვევა კლასიკური სისტემატიკის ძირითად დასკვნებს.
პარალოგიურ გენებს მიეკუთვნება წინამორბედი გენის დუპლიკაციის შედეგად წარმოქმნილი და ევოლუირებული გენები. ასეთი გენები ერთ ოჯახად ერთიანდებიან. პარალოგიური გენები ევოლუირებს ერთი და იმავე სახეობის ფარგლებში (ასევე პარალელურად სხვა სახეობებში). მათ მიეკუთვნება ადამიანში გლობინის მაკონტროლირებელი გენები (იხ. სურ; 13.3). პარალოგიური გენების ევოლუცია ასახავს იმ ცვლილებებს, რომელიც დაგროვდა მათში დუპლიცირების მომენტიდან მოყოლებული დღევანდლამდე. პარალოგიური გენების ფილოგენიით, შეიძლება თვალი გავადევნოთ დუპლიცირებული გენების ისტორიულ განვითარებას - ევოლუციას.
ცილა გლობინები (სისხლის გლობინები, ქსოვილის მიოგლობინები) და მათი მაკონტროლირებელი გენები მხოლოდ ცხოველებს მოეპოვებათ. მათი ჰომოლოგიური, მაგრამ პირველადი სტრუქტურით მკვეთრად განსხვავებული ცილები (ლეგჰემოგლობინები) გააჩნია მცენარეებს. პროკარიოტებში ამ სახის ცილები არაა გამოვლენილი. ამიტომაც გლობინები მოლეკულურ ზეოჯახს ქმნიან, რომლებიც სიცოცხლის ფილოგენური ხის გარკვეულ ნაწილს მოიცავს.
ვ. რატნერისა და თანაავტორების მიერ შემოთავაზებულია გლობინების ზეოჯახის ფილოგენეზური ხე (იხ. სურ. 13.7). გამოირკვა, რომ გლობინების მოლეკულის ევოლუციის საშუალო სიჩქარე არათანაბარი იყო. იგი 400-500 მლნ. წლის წინ მკვეთრად იყო გაზრდილი. ეს პერიოდი ხერხემლიანების მსოფლიო ოკეანიდან ხმელეთზე გადასვლას ემთხვევა. ევოლუციის პროცესში მიმდინარეობდა გლობინების ორგანიზაციული დონის გართულება და გასრულყოფილება.
თავდაპირველად ცხოველებში ფუნქციონირებდა მიოგლობინისა და ჰემოგლობინის პროტომერი (შედგება ერთი სუბერთეულისგან). იგი ამჟამად ზოგიერთ პრიმიტიულ ცხოველს მოეპოვება. ევოლუირების პროცესში მოლეკულამ დიმერის სტრუქტურა შეიძინა. ასეთი კონფიგურაციის ჰემოგლობინი აქვთ მრგვალპირიანებს (სალამურები და მიქსინები). ევოლუირების მომდევნო ეტაპზე α და β - ჯაჭვების შემცველი ტეტრამერული სტრუქტურის ჰემოგლობინი ჩამოყალიბდა.
გლობინების საწყისი გენის დუპლიცირებისა და მათი ასლების დივერგირების შედეგად ორი მონათესავე α და β ოჯახი. სახელდობრ, ამ პერიოდს აღენიშნება ევოლუციის მაღალი საშუალო სიჩქარე. მომდევნო ეტაპზე, რომელიც მრავალ მილიონ წელს მოიცავს გლობინის ევოლუციის სიჩქარე მკვეთრად დაქვეითდა და ბოლოს ნულს გაუტოლდა. მაშასადამე, როგორც კი ჩამოყალიბდა ტეტრამერული სტრუქტურის ჰემოგლობინი, ხმელეთის ხერხემლიან ცხოველებში იგი აღარ შეცვლილა. უნდა აღინიშნოს, რომ ხერხემლიან ცხოველთა ახალ ადაპტურ ზონაში შეღწევამ - (ხმელეთზე გადასვლამ) დიდი მორფო-ფიზიოლოგიური ცვლილებები გამოიწვია (აროგენეზი). ატმოსფერულ ჰაერში არსებული ჟანგბადით სუნთქვას თან ახლდა სუნთქვის სისტემის ძირეული გარდაქმნა, მათ შორის ჰემოგლობინის კონფორმაციის ცვლილება - მან ტეტრამერული სტრუქტურა შეიძინა.
მეტად ჩქარი და ლოკალური მოლეკულური ევოლუცია ნიშანდობლივია გრიპის ვირუსის H3 ჰემაგლუტინინების გენებისთვის. გრიპის ვირუსი რნმ-ს შეიცავს. დადგენილია, რომ რნმ-ში გაცილებით მაღალი სიხშირით ინდუცირდება მუტაცია ვიდრე დნმ-ში. შესწავლილია XXს. ლოკალური ეპიდემიებისა და პანდემიების გამომწვევი გრიპის პათოგენური შტამები. პირველი პანდემია ცნობილია „ისპანკის“ (1918-1919წ.წ.) სახელწოდებით. მომდევნო - „ჰონკონგის“ გრიპის და ა.შ. შეგროვებული და დაცულია კოლექციებში. გრიპის პათოგენური შტამები. სექვენირებულია ამ შტამების შემცველი რნმ, რომლის ანალიზის საფუძველზე აგებულია ფილოგენეზური ხე. ეპიდემიური შტამი წესისამებრ მომდევნო პანდემიის გამომწვევი შტამების უშუალო წინაპარი არ არის. არაეპიდემიური გრიპის ვირუსი ცირკულირებს ადამიანთა ამა თუ იმ ლოკალურ (მცირერიცხოვან) პოპულაციაში. პოპულაციის წევრები ფლობენ ვირუსისადმი მდგრად იმუნიტეტს. ახალი ეპიდემიური აფეთქება ხდება მაშინ, როდესაც ფორმირდება გრიპის ვირუსის ახალი ვარიანტი. ადამიანის პოპულაციას არ გააჩნია ანტისხეულები. ე.ი. ადამიანები ამ ვირუსის მიმართ არ არის იმუნური. ვირუსი ფართოდ ვრცელდება მანამ ვიდრე პოპულაციაში მათ მიმართ ადეკვატური იმუნური პასუხი არ ყალიბდება. რადგან ყველა ეპიდემიები ცნობილია და მათი აღმძვრელი გრიპის ვირუსის შტამები დაცულია კოლექციებში. მათში ჩატარებული მოლეკულური ანალიზის საფუძველზე გამოთვლილია ევოლუციის საშუალო სიჩქარე, რომელიც 3-5-ჯერ მაღალი აღმოჩნდა არაპათოგენურ ფორმებთან შედარებით.
სანიმუშოდ კიდევ ერთ საინტერესო მაგალითს მოვიყვანთ. მაღალი ევოლუციური სიჩქარე ნიშანდობლივია ადამიანის იმუნოდეფიციტის ვირუსისთვის (აივ, ინგლისური აბრევიატურა HIV). იგი რნმ-შემცველი ვირუსია. იწვევს შეძენილ იმუნური დეფიციტის სინდრომს. (შიდსი, ინგლისური აბრევიატურა AIDS). HIV ვირუსის აფრიკის, ვესტ-ინდოეთსა და აშშ გამოყოფილი შტამების შესწავლის საფუძველზე აგებულია ფილოგენეზური ხე (იხ. სურ. 13.8). დადგენილია, რომ HIV ვირუსი არსებობდა ჯერ კიდევ 1960 წლამდე ცენტრალურ აფრიკაში (ზაირი), საიდანაც იგი 70-იან წლებში ვესტ-ინდოეთში (ჰაიტი) გავრცელდა, ხოლო 1978 წლამდე აშშ-ში.
HIV ვირუსის წარმოშობის კერა არის აფრიკა. ამ კონტინენტის ზოგიერთი ქვეყნის მოსახლეობის 50%25 HIV ვირუსის მატარებელია. დაავადება ყველა ადამიანში არ ვლინდება. ვირუსის გამრავლებას ადამიანი-პატრონის იმუნური სისტემა თრგუნავს. HIV ვირუსი ბიოლოგიური თვისებებით მაიმუნის ზოგიერთ ანალოგიურ ვირუსს ემსგავსება. ზოგიერთი მეცნიერი ვარაუდობს, რომ ამ ტიპის ვირუსის მოდიფიცირების შედეგად ჩამოყალიბდა HIV ვირუსი. დიდი ხანი არ არის რაც სპონტანურად მოდიფიცირებული - HIV ვირუსი მაიმუნიდან ადამიანის პოპულაციაში შეიჭრა და გავრცელდა. ამ ვირუსის ვარიბალობის (მუტაციური, რეკომბინაციური) დონე იმდენად მაღალია, რომ იგი ინფიცირებულ ადამიანში 1,5-2 წლის განმავლობაში გაცილებით აგრესიული ხდება ვიდრე შეჭრისას იყო. ავადმყოფში გამომუშავებული ანტისხეულები ვირუსზე ვერ მოქმედებს ვინაიდან იგი სწრაფად ასწრებს ცვლილებას. ვირუსის ცვლილების ტემპს მნიშვნელოვნად ჩამორჩება ადამიანის იმუნური სისტემის მიერ მათ წინააღმდეგ გამომუშავებული შესაბამისი ანტისხეულების ტემპი.
სურ. 13.8. HIV ვირუსის შტამების ანალიზის საფუძველზე აგებული ფილოგენეზური ხე (ვ. რატნერი, 2000). განშტოებაზე ციფრებით აღნიშნულია დივერგირების პროცენტი. მარჯვნივ მითითებულია რეგიონები, სადაც გამოყვეს შტამები.
სწრაფად ვითარდება მოლეკულურ დონეზე ევოლუციური პროცესის კვლევის მეორე მიმართულება. კერძოდ სხვადასხვა ფილოგენეზურ ჯგუფებში დნმ-ის პირველადი სტრუქტურის შედარებითი ანალიზი. ამ გზით ადგენენ ნათესაობის დონეს.
მაშასადამე, ნუკლეინის მჟავების პირველადი სტრუქტურის საერთო მსგავსების შეფასებით შესაძლებელია ევოლუციური ცვლილებების მოლეკულურ დონეზე შესწავლა. ერთიმეორეს ადარებენ დნმ-ის უნიკალურ (განუმეორებად) მონაკვეთებს. სხვადასხვა ფილოგენეზურ ჯგუფებში გაერთიანებული სახეობების დნმ-ის მოლეკულების „ჰიბრიდიზაციისას“ ჰომოლოგიური თანამიმდევრობები ორჯაჭვიან კომპლექსს (დუპლექსს) ქმნიან. ჰიბრიდულ დნმ-ში ისაზღვრება ჰომოლოგიური მონაკვეთების წილი. ცნობილია, რომ დნმ-ის საერთო რაოდენობიდან დაახლოებით 90%25 ცილებს არ აკოდირებს. მიუხედავად ამისა დნმ-ის შესწავლით შესაძლებელია გენების ფილოგენიის შეფასება. განსაზღვრულია შიმპანზესა და ადამიანის დნმ-ის მოლეკულებს შორის პირველადი სტრუქტურის მსგავსება. მათ შორის არაჰომოლოგიური მხოლოდ 1,2%25-ს აღმოჩნდა; ადამიანსა და გორილას შორის - 1,4%25; ადამიანსა და ორანგუტანგს შორის - 2,4%25.
ადამიანსა და მაკაკა რეზუსის დნმ-ის 66%25 ჰომოლოგიური აღმოჩნდა, ადამიანისა და ხარის დნმ-ს 28%25. ჰომოლოგიური უბნები აღმოაჩნდათ, ვირთაგვასთან შედარებისას - 17%25, ორაგულთან - 8%25, ხოლო ნაწლავის ჩხირთან კი - მხოლოდ 2%25; დნმ-ის პირველადი სტრუქტურის შედარებითი ანალიზით მომავალში ევოლუციის მრავალი მნიშვნელოვანი პრობლემა გადაიჭრება.
ამრიგად, მოლეკულურ-ფილოგენეზური ანალიზით ევოლუციის მრავალი კარდინალური საკითხების შესწავლაა შესაძლებელი. რიგ შემთხვევაში ეს მეთოდი ერთადერთია, რომლითაც შესაძლებელია ევოლუციის ისტორიულ პროცესებს ჩავწვდეთ. თანამედროვე ეტაპზე ერთიმეორეს ადარებენ ნეონტოლოგიური სახეობების მაკრომოლეკულებს. პალეონტოლოგიური მასალის გამოყენებით ამ სახის კვლევა თითქმის შეუძლებელია. უნდა აღინიშნოს, რომ შესძლეს ამ მიმართულებით გარკვეული გამოკვლევების ჩატარება. კერძოდ წარსულში მცხოვრები გაყინული ორგანიზმებიდან (მაგ., მამონტი და მისთ.). გამოყვეს დნმ და მოახდინეს მისი სექვენირება. საგანგებო ყურადღებას იპყრობს მცირე ზომის ცხოველთა (ბუზები, ტერმიტები, ფუტკრები, ხოჭო - ცხვირგრძელა) დნმ-ის შესწავლა. ისინი კონსერვირებულია ქარვაში (მცენარეთა ფისი). სადღეისოდ სექვენირებულია ქარვაში ნაპოვნი მწერების და მუმიფიცირებული ადამიანის დნმ-ის ფრაგმენტები. შესწავლილია S18 რიბოსომული რნმ-ის გენები, რის საფუძველზეც აგებულია დენდოგრამა. საჭიროა აღინიშნოს, რომ შესწავლილი ტერმიტებისა და ფუტკრების ასაკი დაახლოებით 25-40 მლნ. წელია, ხოლო ცხვირგრძელასი 120-135 მლნ. წელი.
დასკვნის სახით, საჭიროა აღინიშნოს, რომ მოლეკულურ-ფილოგენეზური ანალიზი ტაქსონომიის ერთ-ერთი მნიშვნელოვანი მეთოდი ხდება. ასე მაგალითად, პრიმატთა სისტემატიკა ამ მეთოდზეა დაფუძნებული. ამ მეთოდს ფართოდ იყენებენ უმაღლესი ტაქსონების სისტემატიკაში. ბაქტერიების მეცნიერული ტაქსონომია მოლეკულურ-ფილოგენეზურ მონაცემზეა დამყარებული. გენოსისტემატიკა ინერგება სოკოების ტაქსონომიაში. ვარაუდობენ, რომ მოლეკულურ-ფილოგენეზური ანალიზი მომავალში გაცილებით დიდ მნიშვნელობას შეიძენს.
|
|
16 § 14. ალტერნატიული კონცეფციები თანამედროვე ევოლუციურ თეორიაში |
▲ზევით დაბრუნება |
თანამედროვე ეტაპზე ევოლუციის სინთეზურ თეორიას ჯერ კიდევ არ გადაუჭრია მის წინაშე დასახული ყველა პრობლემა. მრავალ კითხვას ჯერ კიდევ არ გასცემია ამომწურავი პასუხი. ბიოლოგიის სწრაფი, აღმავალი განვითარება ყველა მისი შემადგენელი დარგის წინაშე სრულიად ახალ ორიგინალურ პრობლემას წარმოაჩენს. ნებისმიერ დასრულებულ თეორიაშიც კი, გარკვეული კორექტივების შეტანა და დაკონკრეტება შეინიშნება. ეს ტენდენცია ნიშანდობლივია ევოლუციური თეორიისთვისაც. ვინაიდან ევოლუციური თეორია განაზოგადებს ბიოლოგიის ცალკეულ დარგში მიღებულ შედეგებს, ამიტომ მის წინაშე სრულიად ახალი მნიშვნელოვანი პრობლემა ისახება. ამ პრობლემათა გადაჭრის პროცესში ზოგიერთი კანონზომიერება უფრო ღრმავდება და ზუსტდება. ზოგჯერ ზოგად კანონზომიერებად მიჩნეული მოვლენა კერძო ხასიათის აღმოჩნდება, ხოლო კერძო მოვლენა - პირიქით ზოგადი ხასიათისა. რიგ შემთხვევაში ახლად მოპოვებული შედეგების ინტერპრეტაციას ალტერნატიული პოზიციიდან ახდენენ.
|
|
16.1 14.1 ნეიტრალური ევოლუციის კონცეფცია |
▲ზევით დაბრუნება |
|
|
16.1.1 კონცეფციის არსი. |
▲ზევით დაბრუნება |
თანამედროვე ეტაპზე ევოლუციურ თეორიაში მწვავე სადისკუსიო პრობლემას ნეიტრალური ევოლუციის კონცეფცია წარმოადგენს. ამ შეხედულების ძირითადი პრინციპები 1968-1969 წ.წ. ჩამოაყალიბეს იაპონელმა მეცნიერებმა მ. კიმურამ და ამერიკელმა მკვლევარებმა ჯ. კინგმა, თ.ჯუკსმა და ჯ.კროუმ. კონცეფციის შემდგომ დამუშავებასა და განვითარებაში დიდი წვლილი შეიტანა მ.კიმურამ და მისმა თანამშრომლებმა. ნეიტრალური ევოლუციის კონცეფცია დასრულებული სახით გადმოცემულია მ.კიმურას წიგნში: „მოლეკულური ევოლუცია: ნეიტრალობის თეორია“ (1983).
ნეიტრალობის თეორია არის ახალი ევოლუციური კონცეფციის მოკლე სახელწოდება. ამ თეორიის თანახმად მოლეკულურ-გენეტიკურ დონეზე პოლიმორფიზმი და ევოლუციური ცვლილება ძირითადად განპირობებულია ისეთი მუტაციებით, რომელზედაც ბუნებრივი გადარჩევის მოქმედება თითქმის ნეიტრალურია.
პოპულაციაში ნეიტრალური მუტაციების დინამიკას, ფიქსაციას ან ელიმინაციას მუტაციური პროცესი და გენთა შემთხვევითი დრეიფი განსაზღვრავს. ამ კონცეფციის თანახმად ევოლუციური პროცესის მიმდინარეობა მოლეკულურ-გენეტიკურ და ფენოტიპურ დონეზე განსხვავებულია.
ნეიტრალური ევოლუციის კონცეფცია ორ ძირითად წყაროს, პოპულაციური გენეტიკის სტოქასტურ (შემთხვევით) თეორიასა და მოლეკულური გენეტიკის მონაცემებს ეფუძნება. სტოქასტურ თეორიაში მნიშვნელოვანია ჯ.ჰოლდეინის მიერ შემოთავაზებული დებულება „საზღაური ევოლუციისათვის“. გენეტიკური ჰომეოსტაზის შენარჩუნება განპირობებულია ჰეტეროზიგოტების სასარგებლოდ მიმართული გადარჩევით, რომელიც, მეორე მხრივ, გენეტიკური ტვირთის წარმოქმნას იწვევს. ნებისმიერი პოპულაცია დიდი რაოდენობით ფლობს ჰეტეროზიგოტულ მდგომარეობაში რეცესიულ საზიანო მუტაციებს, რომელიც ე.წ. გენეტიკურ (მუტაციურ) ტვირთს ქმნის. ამასთანავე, ახლად წარმოქმნილ თაობაში სპონტანურად ინდუცირებული მუტაციების შედეგად პოპულაციის გენოფონდი ივსება და მდიდრდება ახალი მუტაციებით, ანუ ადგილი აქვს მუტაციურ ზეწოლას. ყოველივე ამის გამო, თითოეულმა წყვილმა გაცილებით მეტი შთამომავლობა უნდა დატოვოს, ვიდრე რეალურად ყალიბდება. წინააღმდეგ შემთხვევაში პოპულაციის რიცხოვნობის საშუალო დონე მკვეთრად შემცირდება. თუ დავუშვებთ, რომ პოპულაციაში ფიქსირებულ მუტაციათა დიდი ნაწილი სელექციურად ნეიტრალურია, მაშინ ვითარება მკვეთრად იცვლება. გენეტიკური ტვირთი საკმაოდ მცირდება.
თუ V-თი აღვნიშნავთ კონკრეტულ თაობაში ცალკეულ გამეტაზე მუტაციის სიჩქარეს, N-ით პოპულაციაში ინდივიდთა რაოდენობას, მაშინ მათ მოეპოვებათ 2N ქრომოსომა (აუტოსომური ქრომოსომების ლოკუსებში ამდენივე გენი) და ყოველ თაობაში სპონტანურად 2NU მუტაცია ინდუცირდება (ჰაპლოიდებში ეს მაჩვენებელი Nვ ტოლია). ფიქსირებულ ერთეულ მუტაციათა ალბათობას თუ აღვნიშნავთ უ-თი, მაშინ პოპულაციაში ერთი თაობის განმავლობაში მუტაციის დაგროვების სიჩქარე ტოლი იქნება K=2NVU (I).
კონკრეტულ თაობაში ახლად ინდუცირებული 2NV მუტაციებიდან მხოლოდ ნაწილი ფიქსირდება პოპულაციაში, რომელიც U-ს ტოლია. ამრიგად, K გამოხატავს ევოლუციის პროცესის სიჩქარეს. იგი გამოიხატება იმ რიცხვითი მნიშვნელობით, როდესაც მუტაციის შედეგად ერთი ალელი იცვლება მეორეთი. ამდაგვარი გამოთვლა მხოლოდ ცალკეული ლოკუსისათვის არის სამართლიანი.
დავუშვათ, რომ ინდუცირებული მუტანტური ალელები სელექციურად ნეიტრალურია. ამ შემთხვევაში უ=1/2(2N). თუ ამ მაჩვენებელს (I) ფორმულაში ჩავსვამთ, მაშინ მივიღებთ K=V სიდიდეს. რაც მიანიშნებს, რომ კონკრეტულ თაობაში მუტაციის გზით ალელის შეცვლის სიჩქარე, გამეტაზე გაანგარიშებული მუტაციის სიჩქარის ტოლია. ნეიტრალური მუტაციები დამოუკიდებლად ინდუცირდება და არ არის დამოკიდებული პოპულაციის სხვა პარამეტრებზე (რიცხოვნობა, დინამიკა და მისთ.). ასევე შესაძლებელია გამოვთვალოთ თაობათა ის საშუალო რაოდენობა (თ), რომელიც ერთი საწყისი ალელის, მეორე სელექციურად ნეიტრალური ექვივალენტური მუტაციური ალელით შეცვლას სჭირდება. იგი გაოთხმაგებული პოპულაციის ეფექტური რიცხოვნობის (ე) ტოლია და გამოისახება ფორმულით T=4NE. ჰომოლოგიურ ცილებში ამინომჟავათა თანამიმდევრობის ცვლილების ანალიზის მიხედვით, თეორიული მონაცემები და რეალური შედეგი შეჯერებულია ერთმანეთთან კიმურასა და სხვა ავტორების მიერ. ნეიტრალური ევოლუციის კონცეფციის მიხედვით, სხვადასხვა ფილეტური ჯგუფების ჰომოლოგიურ ცილებში ამინომჟავური თანამიმდევრობის ცვლილება ერთნაირი მუდმივი სიჩქარით მიმდინარეობს.
ცილის უმნიშვნელო ფუნქციის საიტებში ამინომჟავების შეცვლა სელექციური მნიშვნელობით ნეიტრალურია. ამ სახის ცვლილებებზე ბუნებრივი გადარჩევის ზეწოლა არ ხდება. ამიტომ მოლეკულურ-გენეტიკურ დონეზე ნუკლეინის მჟავებისა და ცილების ევოლუცია ბუნებრივი გადარჩევის მოქმედებით არ წარიმართება. გენეტიკური მასალის ევოლუციური ცვლილება სტოქასტური არამიმართული პროცესების მეშვეობით-ნეიტრალური მუტაციების დრეიფით ხორციელდება. მუტაციების ნეიტრალობის ბუნებიდან გამომდინარე, კონცეფციის ავტორები თვლიან, რომ მათი დაგროვება გენომში მიმდინარეობს მუდმივი სიჩქარით, ამდენად სხვადასხვა ორგანიზმებში ცილის მოლეკულის აგებულებაში აღმოჩენილი განსხვავებებით შესაძლებელია საერთო წინაპრიდან მათი დაცილების (დივერგენციის) დროის გამოთვლა.
სურ. 14.1. ხერხემლიან ცხოველებში ჰემოგლობინის α - ჯაჭვის ანალიზის საფუძველზე აგებული ფილოგენეზური ურთიერთდამოკიდებულება. განხილულია ტაქსონების დივერგირების დრო (მ. კიმურა, 1985).
მოლეკულური ევოლუციის მუდმივ თანაბარი სიჩქარით მიმდინარეობაზე (ცილებში ადაპტურ-ნეიტრალური ცვლილების ფიქსაცია) დაყრდნობით ე. ცუკერკანდლმა და ლ. პოლინგმა წამოაყენეს „ევოლუციის მოლეკულური საათის“ იდეა. ამ იდეის თანახმად ცილებში ამინომჟავათა შეცვლის სიჩქარის განსაზღვრით შეიძლება გამოითვალოს კონკრეტული სახეობის არსებობის დრო ე.ი. განისაზღვროს სახეობის და სხვა ტაქსონების (გვარი, ოჯახი, რიგი, კლასი.) დივერგენციის დრო. მრავალრიცხოვანი მასალის ანალიზით დადგენილია, რომ დივერგენციის დრო და ამინომჟავათა ცვლილების სიჩქარე ერთიმეორეს კარგად ემთხვევა (იხ. სურ. 14.1). ასე მაგალითად, ჰემოგლობინის α-ჯაჭვში შეცვლის ფიქსაციის სიჩქარე წელიწადში ამინომჟაურ ნაშთზე K=0,9თ10-9-ს შეადგენს. სხვა ცილები განსხვავებული სიჩქარით ევოლუირებდნენ. ამასთანავე კონკრეტული ცილის ოჯახისათვის (შესაბამისად გენთა ოჯახისათვის) ევოლუციის სიჩქარე მუდმივია. ასე მაგალითად, მაღალი ევოლუციური სიხშირე აღენიშნება ფიბრინოპეპტიდებს (K=8,3თ10-9), მნიშვნელოვნად დაბალი კი C ციტოქრომს (K=0,25თ10-9).
|
|
16.1.2 ნეიტრალური ცვლილებები ცილებსა და დნმ-ში. |
▲ზევით დაბრუნება |
ნეიტრალურ მუტაციად მიჩნეულია გენის ისეთი საიტების ცვლილება (ცვლილება ე.წ. კოვარიონში), რომლის შედეგადაც ამინომჟავათა ნაშთების პოზიციის ცვლილება არ არღვევს ცილის აქტივობას და სივრცით კონფორმაციას (მეორეულ, მესამეულ და მეოთხეულ სტრუქტურას). ცილის აქტიურ ან ალოსტერულ ცენტრებში (ე.წ. ინვარიანტული უბანი) ნებისმიერი ამინომჟავის შეცვლამ შეიძლება ორგანიზმის დაღუპვა გამოიწვიოს.
ჰემოგლობინის ფუნქციონირებაში წამყვან როლს ასრულებს გლობინის ჰემთან დამაკავშირებელი უბანი (ე.წ. „ჰემური ჯიბე“). ამ უბანში ნებისმიერი ამინომჟავური ნაშთის შეცვლა სხვადასხვა სახის ანომალიას იწვევს. გლობინის ზედაპირული უბნები არ მონაწილეობს მეოთხეული სტრუქტურის ფორმირებაში. ამდენად, მასში მომხდარი ცვლილებები მის ფუნქციაზე გავლენას არ ახდენს. დადგენილია, რომ გლობინის α და β ჯაჭვის შიგა უბნებთან შედარებით, ზედაპირულ უბნებში ცვლილებები 10-ჯერ მეტი სიხშირით წარმოიქმნება (იხ. სურ. 14.2).
ციტოქრომები - უძველესი გლობულარული ცილებია. მათი ევოლუცია ორ მილიარდზე მეტ წელს ითვლის. იგი თითქმის ყველა ეუკარიოტულ და პროკარიოტულ უჯრედში გვხვდება და უმნიშვნელოვანეს ენერგეტიკულ პროცესებში მონაწილეობს. ყველა ძუძუმწოვარსა და მრავალ ეუკარიოტში C ციტოქრომი 104 ამინომჟავური რგოლისაგან შედგება. ზოგიერთ ბაქტერიაში ამინომჟავური ნაშთების რიცხვი 135-ს აღწევს. ევოლუციური განვითარების სხვადასხვა საფეხურზე მდგომ 90-ზე მეტ სახეობაში განსაზღვრულია C ციტოქრომის პირველადი სტრუქტურა. მიუხედავად მათი მნიშვნელოვანი ცვლილებისა სივრცითი კონფორმაციები (მესამეული სტრუქტურა) კონსერვატულია და საოცრად ჰგვანან ერთიმეორეს. ევოლუირების პროცესში სხვადასხვა ტაქსონებში გაერთიანებულ სახეობებში C ციტოქრომის შემადგენელი ამინომჟავური ნაშთების 75%25 შეცვლილია. კონკრეტულ სახეობაში შეცვლილ ამინომჟავათა რაოდენობა 5-10%25 არ აღემატება.
სურ. 14.2. ჰემოგლობინის მოლეკულის სტრუქტურა. მითითებულია α - და β - ჯაჭვი, რკინის შემცველი ჰემი; დეფექტური უბნები, რომელიც განაპირობებს ნამგლისებრუჯრედიან ანემიას (ლ. კაიდანოვი, 1996).
ადამიანის ჰემოგლობინის α-ჯაჭვისა და მწერებში მონათესავე ცილა გლობინ ერითროკრუორინის პირველად სტრუქტურის ჰომოლოგიური უბნების რაოდენობა 16%25-ს არ აღემატება. ისინი მეორადი სტრუქტურით უმნიშვნელოდ განსხვავდებიან. რაც შეეხება კონფორმაციაში ე.წ. „ჰემის ჯიბესთან“ α სპირალის განლაგებას, ისინი იდენტურია (იხ. სურ. 14.3).
ნეიტრალური ჩანაცვლება შესაძლებელია მხოლოდ გლობულის ზედაპირულ ნაწილში, თანაც იმგვარად, რომ არ უნდა დაირღვეს მოლეკულის სტერეოქიმიური აღნაგობა. მასტაბილიზებელი გადარჩევის ზეწოლით მიიღწევა ნორმიდან ნებისმიერი გადახრის წუნდება. სტრუქტურული გენების მხოლოდ განსაზღვრულ პოზიციაშია შესაძლებელი არასინონიმური ტიპის ნუკლეოტიდური ცვლილება. ამრიგად, ცალკეულ ლოკუსს მოეპოვება პოზიციათა მცირე რიცხვი, სადაც შესაძლებელია
მუტაციური ცვლილების ფიქსაცია, რომელიც შესაბამის კონტროლირებად ცილაში ამინომჟავური ნაშთების თანამიმდევრობის შეცვლას იწვევს. ასეთ უბნებს კოვარიონი ეწოდება. კოვარიონის თანაპოვნიერება რეალურს ხდის ლოკუსის მკაცრად განსაზღვრულ უბნებში ნეიტრალური მუტაციების ფიქსაციის შესაძლებლობას. ამ სახის ფლუქტუაცია შესამჩნევ ფენოტიპურ ცვლილებას არ იწვევს. სხვადასხვა ფილოგენეზური ხაზების ევოლუირებისას კოვარიონულ უბნებში ეცვლებათ ნუკლეიტიდური თანამიმდევრობა. ბუნებრივია მას თან სდევს ცილის შესაბამის მონაკვეთებში ამინომჟავური ნაშთების ლოკალიზაციის ცვლილება. უნდა აღინიშნოს, რომ ამ ტიპის მუტაციები იწვევენ მხოლოდ მეორეხარისხოვანი თვისებების ცვლილებას. მრავალი მილიონი წლის განმავლობაში ევოლუირების შედეგად ჩამოყალიბებული ძირითადი თავისებურებები პრაქტიკულად უცვლელი რჩება.
სურ. 14.3. I. ადამიანის ჰემოგლობინის α - ჯაჭვის მეორეული და მესამეული სტრუქტურა. II. მწერის ერითროკრუოინის მეორეული და მესამეული სტრუქტურა (ა. ლესკი, ც. ჩოზია, 1980).
მ. კიმურას მიერ მოწოდებულია დებულება, რომლის თანახმადაც: ევოლუციის პროცესში გაცილებით ხშირად მიმდინარეობს ისეთი მუტაციური ცვლილება, რომელიც მოლეკულის სტრუქტურისა და ფუნქციის უმნიშვნელო დარღვევას იწვევს (კონსერვატორული შეცვლა), ვიდრე ისეთი, რომელიც არსებითად არღვევს მოლეკულის სტრუქტურასა და ფუნქციას. კონსერვატორულ ცვლილებაში იგულისხმება სეიმსენსის ტიპის გენური მუტაციის ინდუქცია. ამ შემთხვევაში ერთი სინონიმური ტრიპლეტი მეორეთი იცვლება, რის გამოც ინფორმაციის შინაარსი არ იცვლება. ცილის პირველადი სტრუქტურა უცვლელი რჩება.
ამინომჟავათა უმრავლესობა დეტერმინირებულია ერთზე მეტი ტრიპლეტით. ლოკუსში სეიმსენსის ტიპის ნეიტრალური მუტაციის სიხშირე მაღალია. იგი გენური მუტაციების საერთო რაოდენობის დაახლოებით 30%25-ს შეადგენს.
საჭიროა აგრეთვე აღინიშნოს, რომ ზოგჯერ მუტაციის შედეგად ცილის მოლეკულაში მონომერი მონათესავე თვისებები ამინომჟავით იცვლება. ასე მაგალითად, ტრიპლეტი GUU, რომელიც ვალინს კოდირებს თუ შეიცვალა GCU-თი (კოდირებს ალანინს), მაშინ ცილის მოლეკულაში ვალინის ადგილს მსგავსი თვისების ამინომჟავა ალანინი იკავებს. ამდაგვარი მუტაციით თითქმის არ იცვლება ცილის მოლეკულის თვისება.
ამჟამად დადგენილია, რომ მუტაციები სწრაფად გროვდება ფსევდოგენებში. ასევე სწრაფი მუტაციური გარდაქმნები ნიშანდობლივია ინტრონებისთვის, სპეისერებისა და გენომის იმ ნაწილებისთვის, რომლებიც ფუნქციონირებადი პროდუქტის სინთეზს არ აკონტროლებენ. მუტაციის განსხვავებული ფიქსაცია აღენიშნებათ უნიკალურ და დუპლიცირებულ გენებს. ახალი გენების წარმოქმნა საწყისი გენის დუპლიცირებით ხორციელდება. დუპლიცირებულ გენებზე ბუნებრივი გადარჩევის ზეწოლა აღარ მოქმედებს. მათში გროვდება როგორც თითქმის ნეიტრალური, ისე საზიანო მუტაციები. გენოტიპში ნორმალური და მუტირებული დუპლიცირებული გენების ერთდროული არსებობით იზღუდება მუტირებული გენების მავნე მოქმედება. დუპლიცირებული გენების გარკვეული ნაწილი დივერგირდება და დამოუკიდებელ ფუნქციას იძენს. მათი დიდი ნაწილი დეგენერირდება და შემდეგ იკარგება.
|
|
16.1.3 ცილების პოლიმორფიზმი. |
▲ზევით დაბრუნება |
პოპულაციისათვის ნიშანდობლივია ცილების (ალოზიმების) მიხედვით პოლიმორფულობა. ნეიტრალური თეორიის კონცეფციის მიხედვით ცილებისა და დნმ-ის პოლიმორფიზმი არის მოლეკულური ევოლუციის გარდამავალი ფაზა. ისინი უარყოფენ პოპულაციონისტების თვალსაზრისისს, კერძოდ, რომ პოლიმორფულ ცილათა სისტემები ადაპტურ როლს ასრულებენ პოპულაციაში და ბალანსირებულ ადაპტურ პოლიმორიზმს აყალიბებენ (იხ. §9.3). სადღეისოდ არ მოიპოვება დამაჯერებელი არგუმენტი, ალოზიმების ადაპტურ განსხვავებათა შესახებ. ძნელია დასაბუთება, იმისა, რომ, სელექციური განსხვავება უშუალოდ პოლიმორფული ალელებითაა გამოწვეული, თუ შეჭიდულ ლოკუსებთან ურთიერთქმედებით (მდებარეობის ეფექტით). ასევე არ არსებობს სარწმუნო მტკიცებულება, თუ რამდენად იდენტურია ექსპერიმენტის პირობები სახეობის საარსებო პირობებთან. ალოზიმების მაკოდირებელ ალელებსა და ინვერსიებს შორის წონასწორობის დარღვევის მიზეზად კოადაპტირებულ გენთა კომპლექსების ჩამოყალიბებების დასახელება (პოპულაციონისტური პოზიცია) ნაკლებ სარწმუნოა. გაცილებით მისაღებია ამ მოვლენის დამფუძნებლის პრინციპთა და გენთა შემთხვევითი დრეიფით (ნეიტრალიზმის პოზიცია) ინტერპრეტირება. ჰეტეროზიგოტების სასარგებლოდ მიმართულ გადარჩევის მოქმედების ჰიპოთეზას ასევე ეწინააღმდეგება ჰაპლოიდ ორგანიზმებში პოლიმორფიზმის არსებობის ფენომენი. კიმურა მიიჩნევს, რომ ალოზიმების კლინურ ცვალებადობას მიგრაციები და სელექციურად ნეიტრალური გენების დრეიფი იწვევს. მისივე თვალთახედვით, პოპულაციებში მიმდინარე მოლეკულური განსხვავებები, რომელსაც სელექციონისტური პოზიციიდან ცდილობენ ახსნან, გაცილებით ლაკონურად და დამაჯერებლად ნეიტრალიზმის კონცეფციით იხსნება.
ნეიტრალური ევოლუციის კონცეფცია მოლეკულურ-გენეტიკურ დონეზე მთლიანად როდი გამორიცხავს ევოლუციის ადაპტურ ხასიათს. ზოგიერთ ცილაში ამინომჟავათა შეცვლა (მაგ., ფიბრიონოპეპტიდი) გაცილებით მაღალი სიხშირით მიმდინარეობს სხვებთან (მაგ., ჰისტონები) შედარებით. მცირე სასიცოცხლო მნიშვნელობის ცილებში (ფიბრინოპეპტიდი) ნეიტრალური მუტაციები გაცილებით მაღალ სიხშირით და ტემპით წარმოიქმნება, ვიდრე უმნიშვნელოვანეს სასიცოცხლო ფუნქციაში მონაწილე (მაგ., ჰემოგლობინი, C ციტოქრომი, ჰისტონი) ცილებში.
ნეიტრალიზმის კონცეფცია არ გამორიცხავს ასევე ბუნებრივი გადარჩევის უპირატეს როლს პროგრესულ ევოლუციაში. ე. კიმურა აღნიშნავს, რომ ნეიტრალიზმის თეორია არ ეწინააღმდეგება ბუნებრივ გადარჩევის შესახებ ჩამოყალიბებულ შეხედულებას. ახალი კონცეფცია ევოლუციის პროცესს სრულიად სხვა კუთხით წარმოაჩენს, კერძოდ, მოლეკულურ დონეზე ევოლუციის პროცესში დიდ როლს მუტაციები და გენთა დრეიფი ასრულებს.
|
|
16.1.4 ნეიტრალური ევოლუციის ძირითადი პოსტულატები. |
▲ზევით დაბრუნება |
მოლეკულური ევოლუციის ზოგადი კონცეფცია ფორმულირებულია შემდეგ ემპირიული წესებით:
1. განსხვავებულ ფილოგენეტიკურ ჯგუფებში ნებისმიერი ცილის ევოლუციის სიჩქარე (პოლინგებში) მუდმივი და ერთნაირია.
2. ფუნქციურად მცირე მნიშვნელობის ცილები ან ცილის უბნები მაღალი სიხშირით ევოლუირებს ვიდრე მნიშვნელოვანი ცილები ან ცილის უბნები.
3. მეტი ალბათობითაა ნიშანდობლივი მუტაციის მეშვეობით ისეთი ამინომჟავების შეცვლა, რომელიც ცილის სტრუქტურასა და ფუნქციას უმნიშვნელოდ არღვევს.
4. ფუნქციურად ახალი გენი დუპლიკაციის შედეგად წარმოიქმნება.
5. ევოლუციის პროცესში ნეიტრალური (ან თითქმის უმნიშვნელოდ საზიანო) მუტაციის ფიქსაცია გენომში უფრო ხშირად მიმდინარეობს, ვიდრე სასარგებლო მუტაციის.
შეიძლება დავასკვნათ, რომ ნეიტრალური ევოლუციის კონცეფციამ საზოგადოდ დადებითი როლი შეასრულა. მან გააფართოვა და გააღრმავა ევოლუციური ბიოლოგიის თვალსაწიერი. ახალი კონცეფციის საფუძველზე გადაისინჯა მთელი რიგი ადრე ჩამოყალიბებული პოსტულატები. მისი მეშვეობით გამოვლენილი და შესწავლილია მნიშვნელოვანი მოვლენები, მათ შორის ცილებსა და დნმ-ში მონომერთა ნეიტრალური შეცვლა და მისი ევოლუციური მნიშვნელობა. ამასთანავე ხაზგასმით უნდა აღინიშნოს, რომ ნეიტრალიზმის კონცეფციით ევოლუციის სინთეზური თეორიის შეცვლის მცდელობა ყოველგვარ საფუძველს არის მოკლებული. შეუძლებელია ნეიტრალიზმით აიხსნას ადაპტაციების წარმოქმნა და მათი სრულქმნა. ნეიტრალური მუტაცია აბსტრაქტულად არსებობს. კონკრეტულ საარსებო პირობებში ნებისმიერი მუტაცია შეიძლება ერთ შემთხვევაში აღმოჩნდეს საზიანო, მეორე შემთხვევაში სასარგებლო, მესამე შემთხვევაში გენოტიპის ერთიანი სისტემიდან განმხოლოებით განხილვისას, ნეიტრალური. სადღეისოდ ცნობილია ერთადერთი მიმართულების მქონე ევოლუციის ფაქტორი - ბუნებრივი გადარჩევა.
|
|
16.2 14.2. თანამედროვე სალტაციონიზმი |
▲ზევით დაბრუნება |
სალტაციონიზმის კონცეფციის თანახმად, ახალი სახეობის, გვარების და სხვა სისტემატიკური კატეგორიების ჩამოყალიბება მყისეულად, ნახტომისებური გარდაქმნის შედეგად ხორციელდება. ეს იდეა შორეული წარსულიდან იღებს სათავეს. იგი გარკვეულად სპონტანური ჩასახვის გზით ცოცხალი სისტემების წარმოქმნის შეხედულებებს უკავშირდება. მყისიერად სახეობათა წარმოქმნის იდეა სხვადასხვა ფორმით თავს იჩენდა ანტიკური და საშუალო საუკუნეების მკვლევართა ნაშრომებში. ამდაგვარ შეხედულებებს XIX საუკუნისა და XX საუკუნის დასაწყისში ანვითარებდნენ ნეოკატასტროფიზმის (ვ. ვაგნერი, ო. შინდევოლფი...) და მუტაციონიზმის (ჰ. დე ფრიზი, ვ. ბეტსონი, რ. გოლდშმიტი...) თეორიის მომხრეები.
XX საუკუნის 70 წლებში ეს კონცეფცია კვლავ აღორძინდა პუნქტირალიზმის სახით (ს. ჯოულდი, ნ. ელდრიჯი) და მოწოდებულია „წყვეტილი წონასწორობის“ სახეობათა წარმოქმნის მოდელი. პუნქტირალიზმი მკვეთრად დაუპირისპირდა გრადუალიზმს (იგივე სელექციონიზმი). იგი ძირითადად ეფუძნება პალეონტოლოგიურ მონაცემებს. საჭიროა აღინიშნოს, რომ დარვინმა თავისი კონცეფციის (გრადუალიზმი) დასასაბუთებლად მარჯვედ გამოიყენა პალეონტოლოგიური მასალა.
სურ. 14.4. პუნქტირალიზმისა და გრადუალიზმის მიხედვით სახეობათა წარმოქმნის გრაფიკული მოდელი. ასოებით აღნიშნულია სახეობები (ლ. ტატარინოვი, 1987).
პუნქტირალიზმის ავტორების თვალთახედვით, ევოლუცია ძირითადად ნახტომების გზით მიმდინარეობს. ნახტომთა შორის ევოლუციური სტაზისი ანუ სტაგნაცია შეინიშნება. სახეობის წარმოშობიდან სტაზისი დაახლოებით 1015-1017 თაობათა მანძილზე გრძელდება. ამ პერიოდში სახეობა პრაქტიკულად უცვლელია. სტაზისის შემდეგ საკმაოდ მოკლე პერიოდში (103-104 თაობა) მიმდინარეობს საწყისი სახეობიდან ახალი ფორმების ჩამოყალიბება (იხ. სურ. 14.4). სალტაციონისტები არ გამორიცხავენ გრადუალიზმის გზით სახეობათა ფორმირებას. მათი შეხედულებით ამჟამად არსებული სახეობების 95%25 პუნქტირალიზმის გზით არის ჩამოყალიბებული.
სახეობათა ფორმირება გაცილებით მაღალი სიხშირით ხორციელდება დაკავებული არეალის პერიფერიულ ნაწილში. სახეობათა წარმოქმნა მიმდინარეობს ერთი პოპულაციის ფარგლებში. კერძოდ, პერიფერიული იზოლატების მცირე რიცხოვან ჯგუფებში გენოფონდის სწრაფი (სალტაციური) რეორგანიზაციის შედეგად. ამასთანავე საწყისი ფორმები ელიმინირდებიან და განთავისუფლებულ არეალს ახალი სახეობები იკვებენ.
სახეობათა წარმოქმნის პროცესში არსებით როლს სტოქასტური პროცესები ასრულებენ. მათგან მთავარია გენთა დრეიფი და სისტემური მუტაციები (სალტაციები). სისტემური მუტაციებიდან სახეობათა ფორმირებაში არსებით როლს ასრულებს ისეთი სალტაციები, რომლებიც ფართომასშტაბიან ფენოტიპურ ეფექტს ფლობენ. ამ თეორიის მომხრენი (ჯოულდი, 1982; სტენლი, 1978) სახეობათა წარმოქმნის პროცესში გამორიცხავენ ბუნებრივი გადარჩევის მოქმედებას. მათი მოსაზრებით, დიდი ტაქსონომიური ჯგუფების (დინოზავრები და მისთ.) ამოწყდომა გლობალურმა კატასტროფებმა (კომეტების, მეტეორების ჩამოვარდნა, რადიაციული ფონის გადიდება, ვულკანური ამოფრქვევა და მისთ.) გამოიწვია.
სალტაციონიზმის იდეებმა თავი იჩინეს მოლეკულურ ბიოლოგიაშიც. მათი შეხედულებით, მიკროევოლუცია დაფუძნებულია სტრუქტურული გენების მუტაციაზე, მაკროევოლუცია - რეგულატორული გენების მემკვიდრულ ცვლილებებზე. რეგულატორული გენების მუტაციის შედეგად რადიკალურად იცვლება ონტოგენეზი და ფართომასშტაბიანი ფენოტიპური ცვლილებები ყალიბდება. ზოგიერთი მკვლევარი ახალ სახეობათა წარმოქმნას ვირუსების მეშვეობით უმდაბლესი ორგანიზმებიდან უმაღლეს ფორმებში გენთა ბლოკების გადატანითა და გენომში ინტეგრაციით ხსნის (გენების ჰორიზონტული გადატანა; იხ. §12.10). ამ თეორიის თანახმად, ადამიანის სწრაფი ევოლუცია მაკრომუტაციების (სალტაციები) ინდუქციის შედეგია, რაც აფრიკის ზოგიერთ რეგიონში (სეისმურად აქტიურ რიფტულ ზონაში) მაღალმა რადიაციულმა ფონმა გამოიწვია (იხ. §12.7).
პუნქტირალიზმის კონცეფციის ხელაღებით უარყოფა უმართებულოა, ვინაიდან მრავალი სარწმუნო არგუმენტი მიუთითებს, რომ შესაძლებელია სალტაციის გზით ევოლუციური ცვლილებები. მაგ., მცენარეებში სპორანგიუმისა და თესლკვირტის ჩამოყალიბება, ორლებნიანებიდან ერთლებნიანების წარმოქმნა და სხვ. მრავალი მაგალითის დასახელებაა შესაძლებელი ცხოველთა სამეფოდანაც, კერძოდ, კისრისა და ტანის მალების რიცხვის მომატება ან შემცირება, ძუძუმწოვრებში საძილე არტერიების, აორტის რკალის მარცხენა მხარეს, ფრინველებში - მარჯვენა მხარეს გადატანა და მისთ.
პუნქტირალიზმმა თავისი განვითარების აღმასვლა დაასრულა. ამ კონცეფციის მომხრე მრავალი მეცნიერი, ამჟამად მის დებულებებს აკრიტიკებს. რაც აშკარად მიუთითებს, რომ ბუნებაში გრადუალიზმის გზით სახეობათა ფორმირება ფართოდაა გავრცელელებული ვიდრე სალტაციური (ტატარინოვი, 1987).
ევოლუციის სინთეზური თეორია არ გამორიცხავს სახეობის მყისიერ წარმოქმნას, ვინაიდან სახეობის წარმოქმნის გზები და საშუალებები ნაირგვარია. არსებითია სახეობათა წარმოქმნის ადაპტური ხასიათი, რომელიც მხოლოდ ბუნებრივი გადარჩევის მეშვეობით ხორციელდება (იხ. §12.1).
|
|
17 § 15. ევოლუციის სინთეზური თეორიის ძირითადი პოსტულატები |
▲ზევით დაბრუნება |
ევოლუციური თეორიის განვითარების თანამედროვე ეტაპზე ჩამოყალიბებულია ევოლუციის სინთეზური თეორია. ამ თეორიის ფორმირებაში არსებითი როლი შეასრულა დარვინიზმის ბაზაზე ბიოლოგიის სხვადასხვა დარგის გამოკვლევების სინთეზურმა განზოგადებამ. კერძოდ - პოპულაციის სტრუქტურის გენეტიკურ-ეკოლოგიურმა შესწავლამ (გ. ტურესონი, ნ. ტიმოფეევ-რესოვსკი, ჯ. კლაუზენი და სხვ.), არსებობისათვის ბრძოლისა და ბუნებრივი გადარჩევის ექსპერიმეტულმა და მათემატიკურმა გამოკვლევებმა (ვ.სუკაჩევი, ჯ.ჰოლდეინი, ბ.ქეთლუელი და სხვ.), ექსპერიმენტული და თეორიული გენეტიკის მონაცემებმა (რ.ფიშერი, ს.რაიტი, ნ.დუბინინი, ე.ფორდი, გ.სტებინსი, რ.ლევონტინი და სხვ). ბოლო პერიოდში ევოლუციის სინთეზური თეორიის განვითარებაზე დიდ გავლენას ახდენს მოლეკულურ გენეტიკისა და მოლეკულურ ბიოლოგიაში მოპოვებული შედეგები.
სადღეისოდ ევოლუციის სინთეზურმა თეორიამ დასრულებული სახე მიიღო. ამ თეორიას ჯერ კიდევ არ გადაუჭრია ყველა პრობლემა. ადრე დამუშავებულ და გადაჭრილ საკითხებში შეინიშნება გარკვეული კორექტივების შეტანა და დაკონკრეტება. შემუშავებულია ევოლუციის სინთეზური თეორიის ძირითადი პოსტულატები, რაშიაც დიდი დამსახურება მიუძღვის ფ. დობჟანსკის, ე. ბეზინგერსა და ნ. ვორონცოვს. უნდა აღინიშნოს, რომ ზოგიერთი პოსტულატი დარვინის მიერ არის მოწოდებული. მრავალი მათგანი XXს-ში ჩამოაყალიბეს. ნ.ვორონცოვმა (2004) ამ დებულებებს თავი მოუყარა და ერთიანი სახე მისცა.
ევოლუციის სინთეზური თეორიის პოსტულატები შემდეგი სახითაა ფორმულირებული:
1. მუტაციები არის ევოლუციური პროცესის ელემენტარული მასალა. მუტაციები მემკვიდრული მასალის მცირე, მაგრამ დისკრეტული ცვლილებაა. ევოლუციის სინთეზურ თეორიაში დიდი ყურადღება ეთმობოდა გენურ მუტაციებს. ე.წ. მცირე (წერტილოვანი) მუტაციები არსებით როლს ასრულებენ. ამ თეორიის განვითარების ადრეულ ეტაპზე ქრომოსომულ მუტაციებს ნაკლები, ხოლო გენომურ მუტაციებს უმნიშვნელო როლს ანიჭებდნენ ევოლუციის პროცესში. გენური მუტაციები განაპირობებს პოპულაციაში მაღალი დონით ჰეტეროგენულობის შენარჩუნებას. ქრომოსომული მუტაციები ძირითადად ხელს უწყობს საიზოლაციო მექანიზმების ჩამოყალიბებას, ხოლო პოლიპლოიდიას ეფუძნება მყისიერ სახეობათა წარმოქმნა. ამრიგად, მუტაციური პროცესი არის ელემენტარული მასალის მომწოდებელი ელემენტარული ფაქტორი. ამ პროცესს არამიმართული და შემთხვევითი ხასიათი გააჩნია (აქედან გამომდინარე, ლ. ბერგმა ევოლუციის სინთეზურ თეორიას „ტიხოგენეზი“ ანუ ალბათობაზე დაფუძნებული ევოლუცია უწოდა).
2. ბუნებრივი გადარჩევა წარმოადგენს ევოლუციის მთავარ და წარმმართველ ფაქტორს. გადარჩევა აწესრიგებს, წარმართავს და აძლევს მიმართულებას ევოლუციურ პროცესს. იგი ახდენს შემთხვევით ინდუცირებულ მცირე მუტაციების გადარჩევას (სელექციას). აქედან წარმოდგება ევოლუციის სინთეზური თეორიის მეორე სახელწოდება - სელექციოგენეზი ანუ გადარჩევაზე დაფუძნებული ევოლუცია.
3. პოპულაცია არის ევოლუციის ელემენტარული ერთეული. ზოგიერთი მეცნიერი ვარაუდობდა, რომ ევოლუციის ელემენტარული ერთეული არის სახეობა, ზოგსაც მიაჩნდა - ინდივიდი (კერძოდ „შეძენილი ნიშანთვისების“ მემკვიდრეობის მომხრეები - ლამარკისტები). არც ერთი მათგანი არ აკმაყოფილებს ელემენტარული ერთეულის მოთხოვნებს. კერძოდ, არ უნდა ხდებოდეს მათი მარტივ ერთეულებად დაყოფა, ბუნებრივ პირობებში რეალურად და კონკრეტულად უნდა არსებობდეს, ასევე საჭიროა გააჩნდეს ბიოლოგიური თაობების მანძილზე (დროში) მემკვიდრული ცვლილების უნარი. განმარტების გარეშე ტერმინის „პოპულაცია“ მნიშვნელობა გაურკვეველია. ამ ტერმინის გენეტიკური განმარტება (პოპულაცია არის ინდივიდთა ჯგუფი, რომელთა შორის ერთი თაობის განმავლობაში რეალურად მიმდინარეობს შეჯვარება) ეკოლოგების კრიტიკის საგანი იყო. ისინი გამოყოფენ „გეოგრაფიულ“, „ლანდშაფტურ“, „ეკოლოგიურ“ და სხვა ტიპის პოპულაციებს. XXს. 60-იან წლებში პოპულარობით სარგებლობდა ტერმინი „დემი“, როგორც პოპულაციის ელემენტარული ერთეული, რომელშიც რეალურად ერთი თაობის მანძილზე მიმდინარეობს გენების მიმოცვლა. ჩამოყალიბდა სპეციალური მიმართულება „დემეკოლოგია“. მაგრამ მკაფიოდ არ არის განსაზღვრული დემის ადგილი შიგასახეობრივ იერარქიულ სტრუქტურებში. ამ პრობლემის გადასაწყვეტად საჭიროა ერთიანი მეთოდების მომარჯვებით სპეციალური გამოკვლევების ჩატარება მთელ არეალზე. ასეთი გამოკვლევები კი ძალზე მცირეა.
4. ევოლუცია ფლობს დივერგენციულ ხასიათს ე.ი. ერთი საწყისი ტაქსონის დივერგირების გზით რამდენიმე ტაქსონი ყალიბდება. ყოველ სახეობას ერთი საწყისი ფორმა მოეპოვება, საბოლოო ჯამში ერთი საწყისი პოპულაცია. ამ პოსტულატში შეინიშნება ზოგიერთი უზუსტობა. ევოლუცია ყოველთვის არ არის დივერგენციული ხასიათის (მაგალითად, სტასიგენეზი და ანაგენეზი). ზოგჯერ სახეობას ერთზე მეტი წინაპარი სახეობა მოეპოვება (მაგალითად, ჰიბრიდოგენეზი და სიმბოგენეზი). თუ დავუშვებთ, რომ ევოლუირებს პოპულაცია და არა ინდივიდი, მაშინ დივერგენციის დიქოტომიური ხის საფუძველი არის მრავალი ინდივიდი. მიკროევოლუციის დონეზე ამ წინააღმდეგობის გადაჭრას შეეცადა ფ. დობჟანსკი. მან წამოაყენა რეტიკულური ანუ ბადებრივი ევოლუციის სქემა.
5. ევოლუცია ატარებს თანდათანობით (გრადუალურ) და ხანგრძლივ ხასიათს. ევოლუციის სინთეზური თეორიის მიხედვით სახეობათა წარმოქმნა არის ევოლუციის თვისებრივი ეტაპი. პოპულაციაში თანდათანობით გროვდება ცვლილებები. თაობათა უწყვეტ რიგში თანდათანობითი (გრადუალური) თვისებრივი ცვლილებების შედეგად ერთი კონკრეტული პოპულაცია მეორედ გარდაიქმნება. (აქედან წარმოდგება ამ კონცეფციის კიდევ ერთი სახელწოდება - გრადუალიზმი).
სალტაციონიზმის (მაკროგენეზი) კონცეფციის მიხედვით ახალი სახეობის ჩამოყალიბება, ერთ ან რამდენიმე კონკრეტულ ინდივიდში, ინდუცირებული მაკრომუტაციის (სისტემური მუტაცია) შედეგია. სისტემური მუტაციის ერთ-ერთ ფორმას მიეკუთვნება პოლიპლიდია. იგი ხშირად გვხვდება მცენარეებში, ხოლო ცხოველებში იშვიათად. სალტაციონიზმის განვითარებაში დიდი წვლილი ძირითადად ბოტანიკოსებმა შეიტანეს. ევოლუციის სინთეზური თეორიის ჩამოყალიბებაში ძირითადად ზოოლოგები იღებდნენ მონაწილეობას. ამ პერიოდისათვის ცხოველებში პრაქტიკულად არ იყო ცნობილი სისტემური მუტაციები, მათ შორის პოლიპლიდია. ყოველივე ამან განაპირობა დაპირისპირებული კონცეფციების ჩამოყალიბება.
6. სახეობას პოლიტიპური სტრუქტურა გააჩნია. სახეობა მრავალი იერარქიულად დაქვემდებარებული სტრუქტურისგან შედგება. კერძოდ, მასში გაერთიანებულია მორფო-ფიზიოლოგიურად და გენეტიკურად განსხვავებული ერთეულები - ქვესახეობა, პოპულაცია და მისთ. ამ კონცეფციას ბიოლოგიური სახეობის ფართო პოლიტიპურობის კონცეფცია ეწოდება. ევოლუციის სინთეზური თეორიის ფუძემდებლებმა შესანიშნავად იცოდნენ, რომ ბუნებაში მრავალი მონოფილური სახეობა მოიპოვება. ისინი მცირე არეალს ფლობენ. მათი დანაწევრება ქვესახეობად შეუძლებელია. ასეთებია ენდემური სახეობები, მათ შორის რელიქტიები. ისინი მხოლოდ ერთი პოპულაციითაა წარმოდგენილი. რელიქტიური სახეობის ევოლუციური ბედი ხანმოკლეა.
7. სახეობა არის გენეტიკურად ერთიანი და დახშული სისტემა. პოპულაციათა შორის ალელთა მიმოცვლით - „გენთა ნაკადით“ ხორციელდება სახეობის, როგორც ევოლუციის ეტაპის მდგრადობის შენარჩუნება. ალელთა მიმოცვლა (გენთა ნაკადი) მხოლოდ სახეობის შიგნით მიმდინარეობს. როდესაც მუტაციას დადებითი სელექციური ფასეულობა გააჩნია, მაშინ ის გენთა ნაკადის გზით არეალის მთელ ტერიტორიაზე განსახლებულ ინდივიდებში (ქვესახეობებსა და პოპულაციებში) ვრცელდება. აქედან გამომდინარეობს სახეობის სხარტი და საკმაოდ ტევადი განმარტება: სახეობა არის გენეტიკურად ერთიანი და დახშული სისტემა.
სახეობის ერთიანობას და მდგრადობას პანმიქსია და გენთა ნაკადი უზრუნველყოფს. ნაირგვარი რთული საიზოლაციო მექანიზმებით განპირობებულია, სახეობის გენეტიკურად დახშულ სისტემად არსებობა. იზოლაცია ორგვარ როლს ასრულებს. მისი მეშვეობით სახეობის გენეტიკური სისტემა უცხო (სხვა სახეობების) გენოფონდების გენთა ნაკადისგან არის დაცული (უნდა აღინიშნოს, რომ არა აბსოლუტურად). იზოლაცია განაპირობებს სხვადასხვა სახეობების გენოფონდების დამოუკიდებლად არსებობასა და ევოლუირებას. მეორე მხრივ, იზოლაციით სახეობის შიგნით, პანმიქსია მიიღწევა. პოპულაციათა შორის მიმდინარე გენთა ნაკადით სახეობის ერთიანი გენეტიკური სისტემის შენარჩუნება ხორციელდება.
8. სახეობები ერთიმეორისგან რეპროდუქციულად არის გამიჯნული. ბიოლოგიური სახეობის ერთ-ერთი მნიშვნელოვანი კრიტერიუმია - რეპროდუქციული იზოლაცია. იმ სახეობებისთვის რომლებიც მხოლოდ უსქესოდ მრავლდებიან (ე.წ. აგამური, აპომიქტური, პართენოგენეზური ფორმები) ეს კრიტერიუმი თავის მნიშვნელობას კარგავს. მოწოდებული კრიტერიუმის ფარგლებს გარეთ რჩება პროტისტები და უმდაბლესი ეუკარიოტების მრავალი სახეობა. კერძოდ, ყველა ის ფორმა რომლებიც მხოლოდ უსქესოდ მრავლდება (გვხვდება გამონაკლისი - ზოგჯერ სხვადასხვა სახეობები ეჯვარებიან და ნაყოფიერ შთამომავლობას წარმოქმნიან). ყველა ნამარხი ფორმა სახეობის ბიოლოგიური კონცეფციის მიღმა რჩება. სახეობის ნაცვლად პალეონტოლოგიაში გამოიყენება ეკვივალენტური ტერმინი „ქრონოსპეციესი“ ან „ფრატრია“. ერთი ტერმინის ნაცვლად მეორის გამოყენება ვითარებას არ ცვლის. პალეონტოლოგიური სახეობების რაოდენობა გაცილებით მეტია ვიდრე ნეონტოლოგიურისა.
9. ტაქსონის მონოფილეტური წარმოშობა. ნებისმიერ რეალურ ტაქსონს (გამონაკლისია კრებსითი ტაქსონები) გააჩნია ერთფუძიანი, მონოფილეტური წარმოშობა. მონოფილეტური წარმომავლობა - ტაქსონის არსებობის აუცილებელი პირობაა.
10. ყველა განხილული პოსტულატებიდან გამომდინარეობს, ხომ ევოლუცია არაპროგნოზირებადია, საბოლოო განსაზღვრულ მიზნისკენ არ არის მიმართული ე.ი. არაფინალისტურ ხასიათს ატარებს.
|
|
18 ლიტერატურა |
▲ზევით დაბრუნება |
1. ალექსიძე ნ., ეკოლოგიური ბიოქიმიის საფუძვლები, თბ., 1999
2. გაბუნია ლ., ძველი ხერხემლიანების სამყაროში, თბ., 1987
3. გაგნიძე რ., მცენარეთა გეოგრაფია (ფიტოგეოგრაფია), თბ., 1996
4. გეგეჭკორი ა., ჩემი აფრიკული საფარი, თბ., 2002.
5. გეგეჭკორი ა., ადამიანის ევოლუცია. გაუჩინარებული და ტრაიბალური ცივილიზაციები, თბ., 2004
6. დარვინი ჩ., მოგზაურობა დედამიწის გარშემო გემ „ბიგლით“. თბ., 1951
7. დიასამიძე ა., ევოლუციური მოძღვრების საფუძვლები. ბათუმი, 2000
8. ელიავა ი., ნახუცრიშვილი გ., ქაჟაია გ., ეკოლოგიის საფუძვლები, თბ., 1992
9. თარხნიშვილი დ., კავკასიის ამფიბიები - ეკოლოგია და ევოლუცია. სადოქტორო დისერტაცია., თბ., 2004
10. თარხნიშვილი დ., გენების კალეიდოსკოპი მთიან კუნძულზე,. თბ., 2004
11. ლეჟავა თ., ადამიანის გენეტიკა, თბ., 1998
12. ნათაძე ლ., ხერხემლიან ცხოველთა შედარებითი ანატომია,. თბ., 1962
13. პაპალაშვილი გ., დარვინის მეცნიერული მემკვიდრეობა, თბ., 1961
14. ჟორდანია რ., ხერხემლიანთა ზოოლოგია, თბ., 1997
15. ჟორდანია რ., ზოოტოქსიკოლოგია, თბ., 2004
16. ტიმირიაზევი კ., ჩ, დარვინი და მისი მოძღვრება. თარგმანი და წინასიტყვაობა დ. მელაძისა, თბ., 1938
17. ქაჯაია გ., სახეობის პრობლემა ბიოსისტემატიკაში, თბ., 1984
18. ყურაშვილი ბ., უხერხემლოთა ზოოლოგია, თბ., 1996
19. შათირიშვილი ა., ცარციძე მ., ცაგარელი ს., ზოგადი ბიოლოგია, 1999
20. შათირიშვილი ა., ევოლუციის გენეტიკური მექანიზმების შესწავლა საფუარის ბუნებრივ პოპულაციებში, კრებ.: ბიოლოგია და თანამედროვეობა, თბ., 2002
21. ცაგარელი ს., ბიოსოციოლოგია, თბ., 2004
22. ცარციძე მ., ცაგარელი ს., შათირიშვილი ა., ბიოლოგიის შესავალი, თბ., 1999
23. ჭუჭულაშვილი ი., პოპულაცია ევოლუციის ელემენტარული ერთეული, ქიმია _ ბიოლოგია სკოლაში, №1, 1990
24. ჭუჭულაშვილი ი., გენთა დრეიფი - ევოლუციის ელემენტარული ერთეული, ქიმია - ბიოლოგია სკოლაში, №3, 1991
25. ჭუჭულაშვილი ი., არსებობისათვის ბრძოლა და მისი ფორმები, ბიოლოგია, №2, 1998
26. ჭუჭულაშვილი ი., შათირიშვილი ა., მიკროევოლუციის პროცესების შესწავლა საფუარის ბუნებრივ პოპულაციებში, კრებ.: თანამედროვე ბიოლოგიის აქტუალური საკითხები, თბ., 2000
27. ჯანაშვილი ა., ზოოგეოგრაფია. თბ., 1974
28. Айала Ф., Введение в попюляционную генетику, М., 1984
29. Айла Ф., Кайгер Дж., Современная генетика., т. 3, 1998
30. Алтухов Ю.П., Генетические процессы в популяциях. М., 1989
31. Воронцов Н.Н., Развитие эволюционных идей в биологии, М., 2004
32. Грант В., Эволюция организмов. М., 1980
33. Грант В., Эволюционный процесс. М., 1981
34. Георгиевский А.Б., Дарвинизм. М., 1985
35. Давиташвили Л.Ш., Эволюционное учение, т.1-2, Тб., 1977-78
36. Давиташвили Л.Ш., Теория полового отбора, М., 1961
37. Дарвин Ч., Происхождение видов путем естественного отбора, или сохранение благоприятствующих пород в борьбе за жизнь, СПб., 1991
38. Докинз Р., Эгоистический ген, М., 1993
39. Дубинин Н. П., Избранные труды, т.1-4, М., 2001
40. Захаров И.А., Экологическая генетика и проблема биосферы, Л., 1978
41. Захаров И.А., Сексуальная жизнь божьей коровки, Природа №12, 2001
42. Инге-Вечтомов С.Г., Генетика с основами селекции, М., 1989
43. Иорданский Н.Н., Эволюция жизни, М., 2001
44. Кайданов Л.З., Генетика популяций М., 1996
45. Кейлоу П., Принципы эволюции, М., 1986
46. Кимура М., Молекюлярная эволюция: теория неитральности, М., 1985
47.Левонтин Р., Генетические основы эволюции, М., 1978
48. Майр Э., Зоологический вид и эволюция. М., 1968
49. Майр Э., Популяции, виды и эволюция. М., 1974
50. Мак-Фарленд Д., Поведение животных, М., 1988
51. Малахов В.В., Великий симбиоз: происхождение эукариотической клетки. В мире науки, №2, 2002
52. Мейнрад Смит Дж., Эволюция полового размножения, М., 1981
53. Панов Е.Н., Межвидовая гибридизация у птиц: эволюция в действии, Природа, 6, 2001
54. Парамонов А.А., Дарвинизм, М., 1978
55. Северцов П., Основы теории эволюции, М., 1987
56. Солбриг О., Солбриг Д., Популяционная биология и эволюция. М., 1980
57. Татаринов Л.П., Очерки по теории эволюции, М., 1987
58. Тимофеев-Ресовский Н.В., Воронцов Н.Н., Яблоков А.В., Краткий очерк теории эволюции. М., 1977
59. Фриленд С., Херст Л., Закодированная эволюция, В мире науки, №7, 2004
60. Хедрик Ф., Генетика популяций., М., 2003
61. Хесин Р.Б., Непостоянство генома, М., 1985
62. Шварц С.С., Экологические закономерности эволюции. М., 1980
63. Шеппард Ф., Естественный отбор и наследственность, М., 1970
64. Шмальгаузен И.И., Проблемы дарвинизма, М., 1969
65. Шмальгаузен И.И., Вопросы дарвинизма М., 1990
66. Эрлих П., Холм Р., Процесс эволюции, М., 1966
67. Яблоков А.В., Юсуфов А.Г., Эволюционное учение, М., 2004
68. Яблоков А.В., Популяционная биология, М., 1987
69. Capy P., Langin T., Anxolabehere, Dynamics and evolution of transposable elements. Chapman, Hall, N.Y., 1998
70.Dobzhansky T., Genetics and the evolutionary process. Columbia univ. press, N.Y., 1970
71.Graur D., Li H., Fundamentals of molecular evolution. Suderland, Mass.: sincuer, 2000
72.Maynard Smith S., Evolutionary genetics. Oxford, 1996
ინტერნეტის ზოგიერთი საიტი.
1. http://gened. emc. maricopa. edu-www. mnhn. fz-
2. http://ibss. febras. ru/ebolut
3. http://www. pbs. org/wgbh/evolution/index. Html
4. http://www. pbs. org/wgbh/evolution/library/01/index. html № coevolution
5. http://www. talorigins. org/origins/faqs-evolution. Html
6. http://www. ucmp. berkeley. edu /history/evotheory. Html
7. .http://pespmc 1. vub. ac. be / EVOLUT. Html
8.. http://www. interactiv. com /DARWIN/ Evolution. Html
9.. http://www. becominghuman.org







