|
|
მთავარი - ბიბლიოთეკის შესახებ - ელ.რესურსები |
 |
სამეცნიერო-პოპულარული სტატიები |
|
|
| საბიბლიოთეკო ჩანაწერი: |
| ავტორ(ებ)ი: თევზაძე გელა |
| თემატური კატალოგი სტატიები სხვადასხვა ჟურნალებიდან |
| კოლექციის შემქმნელი: სამოქალაქო განათლების განყოფილება |
| აღწერა: გელა თევზაძის სტატიები გაზ. "24 საათი"-დან |
|
|
1 2010 წლის ნობელის პრემია ფიზიოლოგია-მედიცინაში: ხელოვნური განაყოფიერების უდიდესი მნიშვნელობის აღიარება როგორც მეცნიერული, ასევე ჰუმანიტარული თვალსაზრისით. |
▲ზევით დაბრუნება |
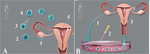
2010 წლის ნობელის პრემია ფიზიოლოგია-მედიცინაში მიენიჭა რობერტ ედვარდსს (Robert G. Edwards) in vitro განაყოფიერების მეთოდოლოგიის განვითარებისათვის (for the development of in vitro fertilization [IVF]). პრემიის ლაურეატის ვინაობის გამოცხადებისას აღინიშნა, რომ ეს პირველი შემთხვევაა, როცა ნობელის პრემია განაყოფიერებისა და გამრავლების დარგშია მინიჭებული. ის გარემოება, რომ in vitro (სიტყვასიტყვით „მინაში“, ანუ სინჯარაში, ე.ი., ხელოვნური) განაყოფიერება (შემდგომში - IVF) ნობელის პრემიისათვის 2010 წელზე ადრე არ იქნა განხილული, ალბათ იმით აიხსნება, რომ საბოლოოდ მხოლოდ ახლახანს დამტკიცდა IVF-ის სრული უვნებლობა ადამიანის ორგანიზმის განვითარებასა და ჯანმრთელობისათვის.



წინამდებარე სტატია მიზნად ისახავს IVF-ის მეცნიერული საფუძვლების აღწერას, ისევე როგორც მასთან დაკავშირებული ეთიკური პრობლემების განხილვას. სტატიის საფუძვლად აღებულია ინფორმაცია, რომელიც ნობელის კომიტეტის ვებგვერდზე ამ თემის შესახებ „დეტალური ინფორმაციის“ (Advanced Information) განყოფილებაშია მოთავსებული:
http://nobelprize.org/nobel_prizes/medicine/laureates/2010/adv.html
(აირჩიეთ „Advanced Information [pdf]“ ბმული).
უნაყოფობა, სამწუხაროდ, მეტად გავრცელებული პრობლემაა - მსოფლიოში 10%25-მდე ოჯახი უშვილოა. ისტორიულად, უნაყოფო ადამიანებისათვის კვალიფიციური სამედიცინო დახმარება თითქმის არ არსებობდა, რის გამოც ისინი იძულებულნი იყვნენ, განკურნების მიზნით საკუთარი ჯანმრთელობისა და ზოგჯერ სიცოცხლის ფასად საეჭვო ღირებულების საშუალებათათვის მიემართათ. ქალების უნაყოფობა უმეტესად უკავშირდება ფალოპის მილების (სადაც კვერცხუჯრედი მომწიფებისა და ოვულაციის შემდეგ ხვდება) დაზიანებას, კაცებისა კი - მცირერიცხოვან ან განუვითარებელ სპერმას.
IVF-ის კვლევის ადრეული ეტაპები.
IVF-ის პირველი ცდებისათვის არჩეული იყვნენ ის ორგანიზმები, რომელთა განაყოფიერება ორგანიზმის გარეთ ხდება. მაგალითად, პირველი მსგავსი ექსპერიმენტი ასკარიდებზე ჩატარდა ჯერ კიდევ XIX საუკუნეში. 1935 წელს კი ხელოვნურ პირობებში მიაღწიეს ბოცვერის ოოციტების (უჯრედები, რომლებისგანაც კვერცხუჯრედები ვითარდება) მომწიფებას გვიანდელ სტადიამდე. 1959 წელს ნაჩვენები იქნა, რომ შესაძლოა in vitro მომწიფებული ოოციტების ასევე in vitro განაყოფიერება და სიცოცხლისუნარიანი ჩანასახის მიღება. მდედრებში გადანერგული ასეთი ჩანასახები ნორმალურად ვითარდებიან და შედეგად ცოცხალი შთამომავლობა იბადება.
ადამიანის IVF - ურთულესი ამოცანა.
მიუხედავად იმისა, რომ XX საუკუნეში ცხოველთა ხელოვნურ განაყოფიერებაში მნიშვნელოვანი წარმატებები იქნა მიღწეული, 1960-იანი წლების დასაწყისამდე ადამიანის ოოციტების IVF-ის მხრივ თვალსაჩინო წინსვლა არ იგრძნობოდა. ამ მიზნის მისაღწევად რამდენიმე როგორც მეცნიერული აღმოჩენა, ასევე ტექნიკური ამოცანის გადაწყვეტა უნდა მომხდარიყო, სახელდობრ: ოოციტების მომწიფების კონტროლირება; IVF-ისათვის შესაფერის სტადიაზე მყოფი ოოციტების შეგროვების მეთოდიკის დამუშავება; სპერმის აქტივაცია (განაყოფიერებისათვის აუცილებელი პროცესი) in vitro პირობებში; IVF-ისა და ჩანასახის ადრეული განვითარებისათვის ოპტიმალური პირობების დადგენა; და ბოლოს, ჩანასახის საშვილოსნოში გადატანის მეთოდის შემუშავება.
რობერტ ედვარდსმა ამ პრობლემებზე მუშაობა 1950-იანი წლების ბოლოს დაიწყო. პირველი პრობლემა, რომელიც მას უნდა გადაეჭრა, იყო IVF-ისათვის გამოსადეგი მომწიფებული ოოციტების შეგროვების მეთოდი. 1965 წელს მან აღმოაჩინა, რომ 24-საათიანი in vitro ინკუბაციის შედეგად ოოციტები მომწიფების პროცესს იწყებდნენ და აღწევდნენ სტადიას, რომელზეც ისინი IVF-ისათვის გამოსადეგნი ხდებოდნენ. 1969 წელს ედვარდსმა ასევე გამოარკვია, რომ ადამიანის აქტივირებულ სპერმატოზოიდებს შეეძლოთ in vitro მომწიფებული ოოციტების განაყოფიერება. ამ მნიშვნელოვანი აღმოჩენით გზა გაეხსნა ადამიანის უნაყოფობის განკურნების მეთოდის დამუშავებას.
გარდატეხა.
ედვარდსის დაკვირვებით, ხელოვნურად განაყოფიერებული ოოციტები ვერ სცდებოდნენ კვერცხუჯრედის ერთხელ გაყოფის სტადიას. მისი ვარაუდით, ეს აიხსნებოდა ორგანიზმის გარეთ გატარებული დროის ხანგრძლივობით. ამ დაბრკოლების გადასალახად, ედვარდსმა მომდევნო ექსპერიმენტებში in vivo (ანუ ორგანიზმში) მომწიფებული ოოციტები გამოიყენა იმ მოსაზრების საფუძველზე, რომ ორგანიზმში სრულად მომწიფებული (და ოვულაციის ზღვარზე მისული) ოოციტები კიდევ უფრო შესაფერისნი იყვნენ როგორც IVF-ისათვის, ასევე განაყოფიერებული ჩანასახის შემდგომი განვითარებისათვის. თაგვების გამოკვლევებზე მიღებული შედეგების საფუძველზე, ისევე როგორც ქირურგიული მეთოდების სრულყოფის შედეგად, კიდევ ერთი მნიშვნელოვანი შედეგი იქნა მიღებული: ოოციტების კონტროლირებადი მომწიფება და ბოჭკოვან-ოპტიკური „თვალის“ დახმარებით მომწიფების ერთ-ერთ ბოლო სტადიაზე ოოციტების უშუალოდ საკვერცხეებიდან ამოღება. 1970 წელს ედვარდსმა და მისმა კოლეგამ - გინეკოლოგმა სტეპტოუმ (Steptoe) - აჩვენეს, რომ შესაძლებელი იყო უნაყოფო ქალებისაგან მომწიფებული და ნაყოფიერი ოოციტების მიღება საკვერცხეებზე მოქმედი ჰორმონით (გონადოტროპინით) მკურნალობის შედეგად (NB: აქ აუცილებელია აღინიშნოს, რომ ასეთი მკურნალობა თვით უნაყოფობას არ კურნავდა, არამედ მხოლოდ გარკვეულ ეტაპამდე განვითარებული ნაყოფიერი ოოციტების შესაძლებლობას იძლეოდა). 1971 წელს კი მიიღეს ხელოვნურად განაყოფიერებული კვერცხუჯრედი, რომელიც ოთხი გაყოფის განმავლობაში (ანუ 16-უჯრედიან სტადიამდე) ნორმალურად ვითარდებოდა in vitro პირობებში. ამ შედეგების ერთობლიობა უმნიშვნელოვანეს მიღწევას წარმოადგენს, რომელმაც საფუძველი დაუდო ადამიანის IVF-ის მეთოდის შემუშავების საბოლოო ეტაპებს.
1970-იანი წლების დასაწყისში ედვარდსი და სტეპტოუ იწყებენ განვითარების ადრეულ ეტაპზე მყოფი ჩანასახების დაბრუნებას დედის საშვილოსნოში. ასზე მეტი წარუმატებელი ცდის შემდეგ და ჰორმონული მკურნალობის ოპტიმიზაციის შედეგად, პირველი ფეხმძიმობა 1976 წელს იქნა მიღწეული. სამწუხაროდ, ეს ფეხმძიმობა უნდა შეწყვეტილიყო, რადგან ჩანასახი ფალოპის მილში (საშვილოსნოს გარეთ) იყო მოთავსებული. ამის შემდეგ ედვარდსმა და სტეპტოუმ გადაწყვიტეს, ჰორმონულ სტიმულაციაზე უარი ეთქვათ: ოოციტების შეგროვების მეთოდის შემდგომი სრულყოფის გამო, მათ შესძლეს, ბუნებრივი ციკლის შედეგად მომწიფებული ოოციტები მიეღოთ და 1978 წლის 25 ივლისს მათი მრავალწლიანი შრომა წარმატებით დაგვირგვინდა: ლუიზ ჯოი ბრაუნი (Louise Joy Brown), ჯანმრთელი და ნორმალურად განვითარებული ბავშვი, დაიბადა ოოციტების IVF-ის შედეგად. ამ შესანიშნავმა მოვლენამ სათავე დაუდო უნაყოფობის მკურნალობის ახალ ერას.
IVF-ის მეთოდის განვითარება.
ლუიზ ჯოი ბრაუნის დაბადების შემდეგ, ედვარდსმა და სტეპტოუმ გახსნეს კლინიკა კემბრიჯში, სადაც დაიბადა მსოფლიოში მეორე და მესამე ბავშვი, რომლებიც IVF-ის შედეგად იყვნენ ჩასახული. აქვე სრულჰყვეს საკვერცხეების ჰორმონული სტიმულაციის და ჩანასახის კულტივირების მეთოდები, რომელთა გამოყენებითაც როგორც კემბრიჯის კლინიკაში, ასევე მის ფარგლებს გარეთაც 1983 წლისათვის 139, ხოლო 1986 წლისათვის უკვე 1000-მდე ბავშვი დაიბადა. დღეისათვის, მსოფლიოში IVF-ით 4 მილიონამდე ბავშვია დაბადებული. განსაკუთრებული აღნიშვნის ღირსია ის გარემოება, რომ ამ ბავშვების პირველი თაობა (ლუიზ ჯოი ბრაუნის ჩათვლით) უკვე ზრდასრულია და ზოგიერთ მათგანს უკვე საკუთარი შვილებიც ჰყავს, რომლებიც IVF-ის გარეშე გააჩინეს.
მეთოდის შემდგომი სრულყოფა.
ედვარდსის აღმოჩენამ მრავალი სხვა მკვლევარი მიიზიდა IVF-ის სფეროში, რამაც ამ პროცესის არაერთი ეტაპის გაუმჯობესება მოიტანა. შესაძლებელი გახდა ოოციტების შეგროვება ულტრაბგერის გამოყენებით, ისევე როგორც ემბრიონების შენახვა გაყინვის გზით. 1994 წელს დაამტკიცდა, რომ შესაძლებელია in vitro მომწიფებული ოოციტების ხელოვნური განაყოფიერება. არანაკლებ მნიშვნელოვანი იყო მომწიფებულ კვერცხუჯრედში ცალკეული სპერმატოზოიდის ინექციის მეთოდის დამუშავება, რის გამოც მამაკაცთა უნაყოფობის მრავალი ფორმა უშვილობას აღარ იწვევდა. ედვარდსის შრომებმა გავლენა მოახდინეს ღეროვანი უჯრედებისა და ზოგადად უჯრედების დიფერენციაციის შესწავლაზე, ისევე როგორც იმპლანტაციისწინა გენეტიკურ დიაგნოსტიკის (preimplantation genetic diаgnostics, PGD) განვითარებაზე. ეს დიაგნოსტიკა საშუალებას იძლევა, IVF-ის შედეგად მიღებულ ჩანასახებიდან გამოაცალკევოს ისინი, რომლებშიც სერიოზული მემკვიდრეობითი დაავადებების განვითარებაა შესაძლებელი, რითაც მნიშვნელოვნად მცირდება ამ დაავადებების შემთხვევები IVF-ის შედეგად გაჩენილ ბავშვებში.
IVF-ით დაბადებულ ბავშვთა ჯანმრთელობა, მრავალი გამოკვლევის საფუძველზე, არ განსხვავდება ბუნებრივი გზით დაბადებული ბავშვების ჯანმრთელობისაგან. IVF-თან დაკავშირებული პრობლემა - მრავალი ჩანასახის ერთდროული განვითარება - უმეტესად გაპირობებულია ერთზე მეტი ხელოვნურად განაყოფიერებული ჩანასახის მოთავსებით საშვილოსნოში, რაც პროცესის ეფექტურობის გაზრდის მიზნით კეთდება. თუ საშვილოსნოში მხოლოდ ერთ ჩანასახს ათავსებენ, ეს პრობლემა თავისთავად ბათილდება. ასევე, ხელოვნურად განაყოფიერებული ჩანასახების მატარებელი ქალების ფეხმძიმობის შეწყვეტის სიხშირე ბუნებრივისაზე ოდნავ მეტია, რაც იოლად იხსნება იმ გარემოებით, რომ ამ პროცესს უმეტესად უფროსი ასაკის ქალები ირჩევენ, და მათი რეპროდუქციული შესაძლებლობები შედარებით დაქვეითებულია.
ეთიკური პრობლემები.
ედვარდსი ხვდებოდა, რომ მისი აღმოჩენა მრავალ ეთიკურ პრობლემას წამოჭრიდა. ის თვითონ იყო მომხრე, რომ გამოკვლევები ადამიანის როგორც სასქესო, ასევე ღეროვან უჯრედებზე მკაცრად ყოფილიყო კონტროლირებული. ედვარდსის ასეთი შეხედულებების მიუხედავად, მას დიდი წინააღმდეგობის გადალახვა მოუხდა: ეკლესიის მამათაგან, რომელნიც ამტკიცებდნენ, რომ ამგვარი კვლევა ამორალური იყო; სახელმწიფო მოხელეთაგან, რომელთა აზრით, პილიტიკური თვალსაზრისით გაცილებით აქტუალური იყო ნაყოფიერების შეზღუდვა და არა უნაყოფობის განკურნვა; კოლეგებისაგან, რომლებიც თვლიდნენ, რომ ედვარდსი ჩანასახის უსაფრთხოების ყველა პირობას არ იცავდა. ამ უკანასკნელი მოსაზრების საფუძველზე, დიდი ბრიტანეთის სამედიცინო კვლევის საბჭომ (Medical Research Council, UK) 1971 წელს უარი განუცხადა ედვარდსსა და სტეპტოუს IVF-ის კვლევის უფლებაზე. ზემოთქმულიდან ცხადია, რომ ედვარდსმა გასაოცარი სიმტკიცე გამოიჩინა, როცა ამ კრიტიკის ფონზე შესძლო, თავისი მეცნიერული თვალთახედვის ერთგული დარჩენილიყო და თავისი ექსპერიმენტების წარმატებული დასასრულისათვის მიეღწია.
დასკვნა.
რობერტ ედვარდსმა განავითარა ადამიანის უნაყოფობის მკურნალობის მეთოდი. ეს აღმოჩენა უმნიშვნელოვანესი მიღწევაა, რომელიც კაცობრიობისათვის უდიდეს დახმარებას წარმოადგენს. ადამიანის ხელოვნურმა განაყოფიერებამ რადიკალური გავლენა მოახდინა: მრავალ ქვეყანაში, ახალშობილების 2-3%25 IVF-ის გზითაა ჩასახული, და ბევრ უნაყოფო წყვილს დახმარების რეალური იმედი უჩნდება. IVF-ის გზით მამაკაცთა უნაყოფობის მრავალი ფორმის დაძლევაც მოხერხდა. IVF-ის განვითარებამ, რომლისათვის 2010 წელს იქნა მინიჭებული ნობელის პრემია ფიზიოლოგიასა და მედიცინაში, შეცვალა მილიონობით უშვილო ადამიანის ცხოვრება და მათ ბავშვების ყოლის სიხარული მიანიჭა.
გელა თევზაძე
24 საათი 01.01.11
|
|
2 2010 წლის ნობელის პრემია ქიმიაში: პალადიუმით კატალიზებული შეწყვილების რეაქციები ორგანულ სინთეზში |
▲ზევით დაბრუნება |
წლევანდელ ნობელის პრემიას ქიმიაში სამი მკვლევარი ინაწილებს: რიჩარდ ჰეკი (Richard F. Heck), ეი-იჩი ნეგიში (Ei-ichi Negishi) და აკირა სუზუკი (Akira Suzuki). ამათგან პირველი ორი ამერიკის შეერთებული შტატებში მოღვაწეობს, მესამე კი - იაპონიაში. პრემიის მინიჭებით ნობელის კომიტეტმა აღიარა ამ მეცნიერების აღმოჩენილი და გამოკვლეული „პალადიუმის მეშვეობით კატალიზებული ჯვარედინი შეწყვილების რეაქციები ორგანულ სინთეზში“ (palladium-catalyzed cross couplings in organic synthesis). ამ კვლევების შედეგებმა დიდი გავლენა მოახდინა როგორც ორგანული ქიმიის განვითარებაზე, ასევე ახალი ნივთიერებებისა და მედიკამენტების შექმნაზე. წინამდებარე მიმოხილვაში მოცემულია ამ რეაქციების არსის ახსნის მცდელობა და მათი პრაქტიკული მნიშვნელობის აღწერა.

ნახ. 1. 2010 წლის ნობელის პრემიის ლაურეატები ქიმიაში:
რიჩარდ ჰეკი, ეი-იჩი ნეგიში და აკირა სუზუკი.
ტერმინების განმარტება
თავდაპირველად, რამდენიმე საკვანძო ტერმინის განმარტებაა აუცილებელი: 1) ორგანული ნივთიერებები ის ქიმიური ნაერთებია, რომლებიც მხოლოდ ცოცხალი ორგანიზმებისთვისაა დამახასიათებელი, და მათ ორგანული ქიმია შეისწავლის (შესაბამისად, იმ ნივთიერებებს, რომლებიც არაცოცხალი ბუნებისთვისაცაა დამახასიათებელი, არაორგანული ქიმია შეისწავლის). 2) ორგანული ნაერთების საფუძველს წარმოადგენს ნახშირბადის ატომების ერთობლიობა, რომლებიც ერთმანეთთან ქიმიური ბმულებით არიან დაკავშირებული; ამ ბმულების წარმოქმნა ქიმიური რეაქციების გზით ხდება. ასეთი რეაქციების ერთ-ერთი მაგალითია ჯვარედინი შეწყვილების (cross coupling) რეაქციები, რომელთა შედეგადაც ნახშირბადის ორი ატომი ერთმანეთს ებმება. 3) კატალიზი პროცესია, რომელიც რომელიმე რეაქციის ჩასატარებლად აუცილებელი არაა, მაგრამ ამ რეაქციას გაცილებით უფრო ეფექტურს ხდის. კატალიზატორი კი ის ნივთიერება ან ნაერთია, რომელიც კატალიზს წარმართავს, და თანაც რეაქციის პროცესში არ იხარჯება, ანუ სუბსტრატისაგან (რეაქციის საწყისი პროდუქტისაგან) განსხვავებით, რეაქციის მიმდინარეობის პროცესში განახლებას არ საჭიროებს. ხშირ შემთხვევაში კატალიზატორი ლითონის ატომია. ამ მიმოხილვაში განხილული რეაქციის კატალიზატორია პალადიუმი, რომელიც პლატინის ჯგუფის ლითონებს განეკუთვნება.
რეაქციის მექანიზმი
პალადიუმით კატალიზებული რეაქციის დროს ნახშირბადის ორი სხვადასხვა ატომი ჯერ ლითონის (პალადიუმის) ერთსა და იმავე ატომთან ქმნის ქიმიურ ბმას, რის შედეგადაც ნახშირბადის ატომები ერთმანეთთან ახლოს აღმოჩნდებიან. სწორედ ამის გამო ხდება უფრო ეფექტური ნახშირბადის ამ ატომებს შორის ბმის წარმოქმნა.
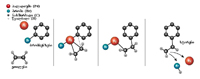
ნახ. 2. ჰეკის რეაქცია ოლეფინსა (ეთილენი) და ნახშირწყალბადის რგოლის ბრომიდს (ბრომბენზენი) შორის: რეაქციის კატალიზატორია პალადიუმი, რომელიც წარმოქმნის ნაერთს როგორც ბრომბენზენთან, ასევე ეთილენთან. ეთილენში ნახშირბადის ატომები ერთმანეთს ორმაგი ბმით უკავშირდებიან, პალადიუმთან ნაერთის წარმოქმნის შემდეგ მათ შორის ერთმაგი ბმა რჩება, მეორე კი პალადიუმთან დასაკავშირებლად გამოიყენება. ამ ნაერთის წარმოქმნის შედეგად ბრომბენზენისა და ეთილენის ნახშირბადის ატომები ერთმანეთს უახლოვდებიან, და მათ შორის ახალი ბმა წარმოიქმნება. საბოლოოდ, პალადიუმის და ბრომის გამოცალკევების შემდეგ, წარმოიქმნება ნივთიერება სტირენი, რომელიც უმნიშვნელოვანესი კომპონენტია პლასტმასების წარმოებისათვის.
წლევანდელი ნობელის პრემიის სამი ლაურეატის წვლილი ამ რეაქციის გამოკვლევაში შემდეგნაირად განისაზღვრება: რიჩარდ ჰეკს ეკუთვნის პიონერული კვლევები ამ დარგში: 1968-1972 წლებში მან პირველმა განსაზღვრა, რომ ნახშირწყალბადების გარკვეული მოლეკულების (ე.წ. ოლეფინების, ანუ ალკენების) შემადგენლობაში მყოფი ნახშირბადის ატომები ჩვეულებრივზე უფრო აქტიურად წარმოქმნიდნენ ქიმიურ ბმებს ნახშირბადის სხვა ატომებთან. მანვე აღმოაჩინა ახალი რეაქცია - ოლეფინებისათვის (მაგ., ნახშირწყალბად ეთილენისათვის) არილის (მაგ., ფენოლის ან ბენზინის) ჯგუფის დამატება. ეს რეაქცია, რომლის კატალიზატორი პალადიუმია, ახლა ჰეკის რეაქციის სახელითაა ცნობილი. 1977 წელს ეი-იჩი ნეგიშიმ მოახდინა ამ რეაქციის ოპტიმიზაცია იმ აღმოჩენის საფუძველზე, რომ ჰეკის რეაქციაში თუთიის ნაერთების გამოყენება ზრდიდა როგორც რეაქციის სპეციფიურობას, ასევე მის გამოსავალს (ანუ პროდუქტიულობას). ამ მოდიფიცირებულ რეაქციას უკვე ნეგიშის სახელი ეწოდა. აკირა სუზუკიმ 1979 წელს თუთია შეცვალა ბორის ნაერთებით, რამაც რეაქცია მრეწველობის მიზნებისათვის ბევრად უფრო პრაქტიკული გახადა: ბორი არატოქსიკურია, რეაგირებს ბევრ ნახშირწყალბადთან, და რეაქციის ჩატარების პირობები შედარებით მარტივია. ეს მოდიფიცირებული რეაქცია, რომელსაც სუზუკის სახელი ეწოდა, ხშირად და წარმატებულად გამოიყენება ფარმაკოლოგიურ მრეწველობაში.
აღმოჩენის პრაქტიკული მნიშვნელობა
ჰეკის, ნეგიშისა და სუზუკის რეაქციები გამოიყენება 100-ზე მეტი ქიმიური ნაერთის სინთეზისათვის, მათ შორისაა: ტაქსოლი, მორფინი, სტეროიდები, სტრიქნინი, ისევე როგორც მრავალი რთული ქიმიური ნაერთი ან მედიკამენტი. ამ რეაქციების მეშვეობით ხელოვნურად სინთეზირებული ნაერთების მაგალითებია: 1) დისკოდერმოლიდი - ნაერთი, რომელიც ბუნებაში ზღვის ღრუბელის ორგანიზმში არსებობს და ანტიკანცეროგენული თვისებებით გამოირჩევა; 2) დიაზონამიდი, რომელიც ეფექტურ საშუალებადაა მიჩნეული სწორი ნაწლავის კიბოს წინააღმდეგ; 3) დრაგმაციდინი F, რომელიც გამოიყენება ჰერპესის ვირუსის და შიდსით დაავადებულთა მკურნალობისას.
ჰეკის, ნეგიშისა და სუზუკის რეაქციები ასევე მნიშვნელოვანია ბუნებაში არსებული მედიკამენტების ეფექტურობის გაზრდისათვის ქიმიური მოდიფიკაციის გზით. ამგვარი მოდიფიკაციის მაგალითია ვანკომიცინი - ანტიბიოტიკი, რომლის საწყისი ფორმა ნიადაგიდან იყო გამოყოფილი კუნძულ ბორნეოზე. შემდგომ რამდენიმე ბაქტერიამ ამ ანტიბიოტიკისადმი მდგრადობა გამოიმუშავა, და სასურველი გახდა ვანკომიცინის მოდიფიკაცია. ჰეკის, ნეგიშისა და სუზუკის რეაქციებით მოდიფიცირებული ვანკომიცინი უკვე იმ ბაქტერიებსაც სპობს, რომლებიც საწყისი ფორმის მიმართ მგრძნობიარენი არ არიან.
პალადიუმით კატალიზებული შეწყვილების რეაქცია ასევე გამოიყენება კომპიუტერების წარმოებაში, კერძოდ, ორგანული სინათლის გამომსხივებელი დიოდებისათვის (OLED, organic light emitting diodes). ასეთი - სინათლის წარმომქმნელი ორგანულ მოლეკულების შემცველი - დიოდები გამოიყენება უაღრესად წვრილი (რამდენიმე მილიმეტრი სისქის) ეკრანების დასამზადებლად. მათ მიერ გამოსხივებული ლურჯი სინათლის ოპტიმიზაცია სწორედ ჰეკის რეაქციის გამოყენებით ხდება.
დასკვნა
ჰეკის, ნეგიშისა და სუზუკის რეაქციების შემდგომი ოპტიმიზაცია და გაუმჯობესება, როგორც სჩანს, კიდევ დიდი ხნის განმავლობაში მოხდება. ასეთი გაუმჯობესების ერთ-ერთი მაგალითი დაკავშირებულია 2010 წლის ნობელის პრემიასთან ფიზიკაში: ამ ნაშრომის ერთ-ერთი მნიშვნელოვანი შედეგია გრაფიტისა და პალადიუმის შეკავშირება, რის შედეგადაც შესაძლებელი გახდა სუზუკის რეაქციის ჩატარება წყლის გარემოცვაში.
ამრიგად, თუმცა ჰეკის, ნეგიშისა და სუზუკის რეაქციებს უკვე უდიდესი მნიშვნელობა აქვთ მოპოვებული, მაინც სავსებით შესაძლებელია, რომ ამ ქიმიური რეაქციების როლი ფარმაკოლოგიისა და ტექნოლოგიის განვითარებაში მომავალში კიდევ უფრო გაიზარდოს.
ავტორი მადლობას უხდის ირაკლი მჭედლიშვილს (საერთაშორისო ურთიერთობების კვლევის ცენტრი) და დავით მანაგაძეს (NIH) შენიშვნებისა და კომენტარებისათვის.
გელა თევზაძე
24 საათი 30.01.11
|
|
3 FoldIt - სტრუქტურული ბიოლოგიისა და კომპიუტერული თამაშების სიმბიოზი. |
▲ზევით დაბრუნება |
კომპიუტერული თამაშებით გატაცება დღეს სერიოზულ პრობლემას წარმოადგენს. ერთადერთი გამართლება, რომელიც ამ მოვლენას შეიძლება მოენახოს, ისაა, რომ რომ თამაშების მოცემულობასა თუ წესებში ზოგჯერ „შეპარულია“ ინფორმაცია, რომელიც მოთამაშეს ზოგადი ცოდნის დაგროვებაში უწყობს ხელს (მაგალითად, ეხმარება ინგლისური ტერმინების შესწავლაში, ისტორიული ან გეოგრაფიული ფაქტების გაცნობაში და ა.შ.). სწორედ ამ მოვლენის გამოყენებას ცდილობს კომპიუტერული თამაში FoldIt, რომელიც მოთამაშეს სთავაზობს ბიოლოგიის ერთ-ერთი მნიშვნელოვანი დარგში - ცილის სტრუქტურის ამოხსნაში - მონაწილეობის მიღებას.

1. FoldIt-ის გამომგონებელი - ბიოქიმიკოსი დევიდ ბეიკერი (David Baker, University of Washington, Seattle).
მაშ ასე, რას გვთავაზობს ეს თამაში და რითი განისაზღვრება მისი უნიკალურობა? პირველ რიგში, სხვა თამაშებისაგან განსხვავებით, ამ თამაშის მონაწილეებს შესაძლებლობა ეძლევათ ჩაერთონ (და თავისი წვლილი შეიტანონ) ჭეშმარიტ სამეცნიერო პროცესში, ჯერ-ჯერობით ამოუხსნელი პრობლემების (ცილების სტრუქტურების) გადაწყვეტის გზით.
დასაწყისისათვის კი განვიხილოთ ამ თამაშის (ანუ ცილების სტრუქტურის) თეორია. ცილები ყოველი ცოცხალი ორგანიზმის საფუძველია (შედარებით ხანდაზმულებს, ამ სტრიქონების ავტორის დარად, ალბათ მოაგონდებათ ფრიდრიხ ენგელსის განსაზღვრება: „სიცოცხლე ცილოვან სხეულთა არსებობის გზაა...“, რომელიც კომუნისტების მმართველობის დროს კლასიკურად იყო მიჩნეული). ყველა ცილის შემადგენელი ნაწილებია ამინომჟავები. ამინომჟავები შედარებით მცირე ომის მოლეკულებია, რომლებიც, თავის მხრივ, აზოტის, წყალბადის, ნახშირბადის, ჟანგბადისა და ზოგიერთ შემთხვევაში გოგირდის ატომებისაგან შედგებიან. ამინომჟავების შემადგენლობაში შედის ატომების ძირითადი ჯგუფი (რომელიც ყველა ამინომჟავაში ერთნაირია), და გვერდითი ჯგუფი, რომელიც 20 არსებულ ამინომჟავას ერთმანეთისაგან განასხვავებს. ცილის მოლეკულის წარმოსაქმნელად საჭიროა ამინომჟავების ერთმანეთთან შეკავშირება ძირითადი ჯგუფების გადაბმის გზით, რის შედეგადაც წარმოიქმნება განსაზღვრული სიგრძის ჯაჭვი. ეს ცილის პირველადი სტრუქტურაა. ამ სტრუქტურის საილუსტრაციოდ წარმოვიდგინოთ ერთმანეთზე ხელჩაკიდებული ხალხის ჯგუფი, რომელიც წრფის სახით არის განლაგებული. ამ ჯგუფში ხელები ამინომჟავების ძირითადი ჯგუფის ანალოგიაა, ადამიანების თავები კი - გვერდითი ჯგუფებისა. გვერდითი ჯგუფები, იმისდა მიხედვით, თუ რა ატომებს შედის მათ შემადგენლობაში, რამდენიმე კატეგორიად იყოფა: წყალში ხსნადი ან წყალში უხსნადი, დადებითად ან უარყოფითად დამუხტული.
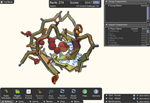
2. FoldIt თამაშის ერთ-ერთი სტადია კომპიუტერის ეკრანზე - ცილის სტრუქტურა ამოხსნის პროცესში. ნაჩვენებია წყალში უხსნადი (ნარინჯისფერი) და ხსნადი (ლურჯი) ამინომჟავები, აგრეთვე „შეჯახებული ატომები“ (წითელი წრეები).
ცილები ამ წრფივ მდგოვარეობაში არ რჩებიან: ისინი იკეცებიან, ანუ „იგორგლებიან“ (fold), რითაც პირველად სტრუქტურასთან შედარებით უფრო კომპაქტურ კონფორმაციას ქმნიან. ზოგადად, ცილების მეორადი სტრუქტურა განისაზღვრება ამინომჟავების მხოლოდ ძირითად ჯგუფებში არსებული ქიმიური ბმებით წყალბადის ატომებს შორის, ცილების მესამეული სტრუქტურა კი - ამინომჟავების როგორც ძირითადი, ასევე გვერდითი ჯგუფების ურთიერთქმედებითა და მათი განლაგებით სივრცეში. FoldIt-ის მიზანი სწორედ ცილის მესამეული სტრუქტურის გამოთვლაა, მისი შემადგენლობის (ანუ პირველადი სტრუქტურის) საფუძველზე.
მესამეული სტრუქტურის თვალსაჩინო გამოსახულებაა გრძელი ძაფი, რომელიც გორგლადაა დახვეული. დაგორგვლის პროცესში ზოგიერთი ამინომჟავა გორგალის შიგნით ექცევა, ზოგი კი გარეთ რჩება. ზოგი ამინომჟავა (პირველად სტრუქტურაში ერთმანეთისაგან დაშორებული) ერთმანეთს უახლოვდება, ზოგი კი ერთმანეთისაგან განიზიდება. ურთიერთქმედების ეს სახეები ამინომჟავების გვერდითი ჯგუფების თვისებებზეა დამოკიდებული. ყველა ცილა მისთვის დამახასიათებელ გორგლად იხვევა, და ეს სტრუქტურა მაქსიმალურად სტაბილურია ყველა შესაძლებლობიდან. თუკი ცილა რომელიმე სხვა კონფორმაციას მიიღებს, ის საბოლოოდ მაინც ყველაზე სტაბილურ მდგომარეობაში დაბრუნდება (ისევე, როგორც მთის წვერზე მოთავსებული ბურთი საბოლოოდ მთის ძირში ჩამოგორდება, სადაც ის მყარად მოთავსდება).
ცილები ასრულებენ როგორც სტრუქტურულ (ორგანიზმების „სამშენებლო მასალის“), ასევე უჯრედებში მიმავალი მრავალი ბიოქიმიური რეაქციის წამმართავ (ფერმენტულ) და დამხმარე (კატალიზურ) როლებს. ცილების ამ მრავალფეროვანი ფუნქციებისათვის აუცილებელია არა მარტო ცილის გარკვეული შემადგენლობა (ანუ ცილის შემადგენელი ამინომჟავების თანმიმდევრობა), არამედ ის სტაბილური კონფორმაციაც, რომელსაც ცილის თავდაპირველად წრფივი მოლეკულა იღებს „დაგორგვლის“ შედეგად. როგორც ზემოთ განვიხილეთ, ასეთი „დაგორგვლა“/დაკეცვა (folding) ძირითადად ამინომჟავების თვისებებსა (ხსნადობა, ელექტრული მუხტი) და მათ ურთიერთქმედებაზეა დამოკიდებული. ამრიგად, ცილის პირველადი სტრუქტურა (ცილაში შემავალი ამინომჟავები და მათი თანმიმდევრობა) ამ ცილის „გორგლის“ სტრუქტურის გამოცნობის ან, ყოველ შემთხვევაში, დასაბუთებული ვარაუდის გაკეთების საშუალებას იძლევა. მთავარი ამოცანა ამ შემთხვევაში ოპტიმალური სტრუქტურის არჩევაა, ანუ ისეთი კონფორმაციისა, რომელიც მაქსიმალურ სტაბილურობასა და, მაშასადამე, ცილის ფუნქცი(ებ)ის შესრულებას საუკეთესოდ უზრუნველჰყოფს.
ახლა კი გავეცნოთ FoldIt კომპიუტერული თამაშის ძირითად წესებს, მის შემადგენლობას/აგებულებასა და მიზნებს:
თამაშის წესები განისაზღვრება რამდენიმე ძირეული პრინციპით, რომელთა გათვალისწინება აუცილებელია ცილის „დაგორგვლის“ პროცესის განსაზღვრისას:
1. კომპაქტურად „დაგორგლეთ“ ცილა. რაც უფრო მცირე ზომისაა „დაკეცილი“ ცილა, მით უკეთესი. ამის შესასრულებლად, ცილის „ნაკეცებს“ შორის არ უნდა იყოს ცარიელი ადგილები, სადაც წყლის მოლეკულებს შეეძლებათ შეღწევა. ამიტომ ცილის შემადგენელი ატომები ერთმანეთთან მაქსიმალურად ახლოს უნდა იყვნენ მოთავსებული. ზოგიერთ შემთხვევაში, ცილის ფრაგმენტები ერთმანეთთან მიახლოებისას ამ ფრაგმენტების დამაკავშირებელ ქიმიურ ბმებს ქმნიან, რაც ცილის კომპაქტურ დაკეცვას ხელს უწყობს.
2. მოათავსეთ წყალში უხსნადი მოლეკულები ცილის „გორგალის“ შიგნით. იმის გამო, რომ ცილების უმეტესობა წყლის გარემოშია მოთავსებული, წყალში უხსნადი „გვერდითი ჯგუფების“ გორგლის ზედაპირზე მოთავსება მიზანშეწონილი არაა. ასეთი ჯგუფები FoldIt-ში ნარინჯისფრადაა აღნიშნული. ამინომჟავების წყალში ხსნადი ნაწილები (ლურჯი ფერი FoldIt-ში) კი, პირიქით, გორგლის ზედაპირზე უნდა განთავსდნენ.
3. ერიდეთ ატომთა „შეჯახებას“. ორი ატომი ერთსა და იმავე ადგილზე ვერ მოთავსდება. თუ ცილის „გორგალში“ ამინომჟავების გვერდითი ჯგუფები ერთმანეთთან ზედმეტად ახლოსაა/ერთმანეთს „გადაეფარება“, ეს თამაშის წესების დარღვევად ჩაითვლება. FoltIt-ში ასეთი სივრცის ნაკლებობა ატომებისათვის წითელი წრის სახით აღინიშნება.
ახლა კი თვით თამაშის სტრუქტურა განვიხილოთ:
FoldIt-ის პირველი ნაწილია „შესავალი თავსატეხები“ (Intro Puzzles), რომელიც განკუთვნილია დაწყებითი დონის მოთამაშეებისათვის. ამ ნაწილში მოთამაშეები [გა]ეცნობიან თამაშის იმ ძირითად კონცეფციებსა და ხელსაწყოებს, რომლებიც მათ დასჭირდებათ თამაშის მეორე ნაწილში („მეცნიერული თავსატეხები“, Science Puzzles) ცილების სტრუქტურის გამოსათვლელად, ანუ მათ „დასაკეცად“. როგორც აღვნიშნეთ, ასეთი გამოთვლა ხდება ამ ცილების შემადგენელი ამინომჟავების განლაგებასა და მათი შესაძლებელი ურთიერთქმედების საფუძველზე. თამაშის ამავე ნაწილში მოცემულია ცილების სტრუქტურის გარკვევის ძირითადი წესები (იხ. ზემოთ).
თამაშის მეორე ნაწილის (Science Puzzles) ერთ-ერთი მიზანია დაამტკიცოს, რომ ცილების ოპტიმალური სტრუქტურის ამოცნობის ზოგიერთი ასპექტის გადაწყვეტას ადამიანები გაცილებით უკეთ ახერხებენ, ვიდრე ამ მიზნისათვის შექმნილი კომპიუტერული პროგრამები. ეს თამაში ასევე მიზნად ისახავს მეცნიერებისა და ბიოტექნოლოგიური კომპანიების დარწმუნებას პროცესის ეფექტურობაში. თამაშის კიდევ ერთი მიზანია თამაშის განმავლობაში შექმნილი სტრატეგიული მიდგომების დანერგვა ცილის სტრუქტურის გამომთვლელ კომპიუტერულ პროგრამებში ამ პროგრამების ეფექტურობის გაზრდის მიზნით. ეს მიზნები ერთმანეთისაგან მეტ-ნაკლებად დამოუკიდებელია და შესაძლოა როგორც ერთ-ერთი მათგანი, ასევე ყველა განხორციელდეს.
FoldIt-ის გაცილებით უფრო საინტერესო მიმართულება, ალბათ, არა უკვე არსებული (ანუ სპეციალისტების მიერ გამოთვლილი) სტრუქტურების გამოცნობაა, არამედ სამედიცინო თუ ბიოტექნოლოგიური მიზნებისათვის გამოსადეგი ცილების „დაპროექტება“ (protein design). ამ მიდგომას გაცილებით უფრო უშუალო პრაქტიკული გამოყენება აქვს, ვიდრე სტრუქტურის გამოცნობას. აქ, ისევე როგორც საინჟინრო სფეროში, აქაც თამაშის მიზანს წარმოადგენს გარკვეული მექანიზმის დაპროექტება, მაგალითად გარკვეული ტიპის ვირუსის გაუვნებელყოფისათვის, ან ჰაერიდან ნახშირორჟანგის შთანთქმისათვის საჭირო მექანიზმის შექმნა. ამასთან ერთად, იმის გამო, რომ კომპიუტერული თამაშებით დაკავებული ადამიანების რიცხვი როგორც წესი ძალიან დიდია, ნაკლებ სავარაუდოა, რომ FoldIt-ს კონკურენცია გაუწიონ ცილების „დაპროექტებისთვის“ შექმნილმა დღეს არსებულმა სპეციალიზირებულმა სამეცნიერო კომპიუტერულმა პროგრამებმა.

3. ცილის „დაუგორგლავი“ (არასტაბილური, A) და „დაგორგლილი“ (სტაბილური, B) სტრუქტურები.
ამასთანავე, ცილის მესამეული სტრუქტურის გამოთვლას უდიდესი მნიშვნელობა აქვს როგორც მისი მოქმედების მექანიზმის გასაგებად, ასევე, ავადმყოფობის გამომწვევი ცილების შემთხვევაში, მათი საწინააღმდეგო მედიკამენტების შესაქმნელად. მცირე ზომის ცილებისთვისაც კი, ოპტიმალური სტრუქტურის მონახვა მეტად ძნელია, რადგან სხვადასხვა შესაძლო კონფორმაციების რაოდენობა ჭეშმარიტად ასტრონომიულია. ამ მრავალი კონფორმაციიდან საუკეთესოს მონახვა თანამედროვე ბიოლოგიის ერთ-ერთი ერთულესი ამოცანაა. უკვე არსებული მეთოდები, მძლავრი კომპიუტერების გამოყენების მიუხედავად, მნიშვნელოვან დანახარჯებსა და დიდ დროს მოითხოვს. ამ პრობლემის გადასაწყვეტად FoldIt-ში გამოიყენება როგორც თამაშის მონაწილე მრავალი ადამიანის ინტუიცია და ტექნიკურ ამოცანათა გადაწყვეტის უნარი, ასევე მათი ერთმანეთთან შეჯიბრების ფაქტორი: ყოველი მოთამაშის მიზანია, სხვებზე სწრაფად გამოარკვიოს ცილის ოპტიმალური სტრუქტურა.
და ბოლოს, რამდენიმე კონკრეტული მაგალითი იმის საილუსტრაციოდ, თუ რა პრობლემების გადაჭრა ძალუძთ FoldIt-ის მოთამაშეებს:
შიდსი: შიდსის ვირუსი ადამიანის ორგანიზმში შეღწევისას წარმოქმნის ორ ცილას, რომლებიც მის გამრავლებასა და ადამიანის ინფიცირებას უწყობს ხელს. შიდსის საწინააღმდეგო პრეპარატები სწორედ ორი ცილის სტრუქტურის გამორკვევის საფუძველზეა შექმნილი, და ამ პრეპარატების გაუმჯობესების ერთ-ერთი გზა ისევ სტრუქტურის დაზუსტებაშია.
კიბო: შიდსისგან განსხვავებით, კიბოს იწვევს თვით ადამიანის ორგანიზმში არსებული (და არა გარედან შემოტანილი) ცილები, რომლებიც გავლენით ადამიანის უჯრედების უკონტროლო გაყოფა ხდება. თუ ასეთი გაყოფის შემაჩერებელი ცილა დაზიანებულია, კიბოს განვითარება გარდაუვალია. ამ ცილის სტრუქტურის გამოკვლევაც მნიშვნელოვანია მისი გაუმჯობესებული ანალოგების შექმნისათვის.
ალცჰაიმერის დაავადება: არსებული თეორიის მიხედვით, ალცჰაიმერის დაავადების მიზეზია ერთ-ერთი (ჯანმრთელ უჯრედებშიც არსებული) ცილის დანაწევრების შედეგად წარმოქმნილი ფრაგმენტების შეწებება (აგრეგაცია). არაა გამორიცხული, რომ ამ აგრეგატების დაშლა შესაძლო გახდება „დაპროექტების“ (protein design) გზით შექმნილი ცილების დახმარებით.
თამაშის დეტალებით დაინტერესებულთათვის, ან მასში მონაწილეობის მიღების მსურველთათვის საჭირო ინფორმაცია შემდეგ ბმულებზეა განთავსებული:
http://fold.it/portal/ - FoldIt-ის ვებგვერდის მთავარი გვერდი.
http://fold.it/portal/info/faq - ზოგადი ინფორმაცია FoldIt-ის შესახებ: თამაშის წესები, მიზნები, ქულების დარიცხვის სისტემა, ტექნიკური პრობლემები, შესაძლებელი სირთულეები.
http://fold.it/portal/info/science - ცილების სტრუქტურის თეორიული საფუძვლები, FoldIt-ის თემაზე გამოქვეყნებული სტატიების სია.
http://www.facebook.com/pages/Foldit/95793463004 - FoldIt-ის გვერდი Facebook-ზე.
http://www.nature.com/news/2010/100804/full/466685a.html - სტატია FoldIt-ზე ჟურნალში „Nature“.
გელა თევზაძე
24 საათი 13.02.11
|
|
4 ცოცხალი ორგანიზმების კლონირება: შთამბეჭდავი შედეგები და სერიოზული პრობლემები |
▲ზევით დაბრუნება |
14 თებერვალი, ყველასათვის ცნობილი დღესასწაულის გარდა, კიდევ ერთი მნიშვნელოვანი მოვლენის დღეა: 2003 წლის ამ დღეს დოლი (Dolly) - პირველი ცხვარი (და, ზოგადად, პირველი ძუძუმწოვარი), რომელიც კლონირების შედეგად დაიბადა, დაიღუპა 6 წლის ასაკში ფილტვების ინფექციის შედეგად. რასაკვირველია, ამ ფაქტის მნიშვნელობა არა დოლის დაღუპვით, არამედ მისი დაბადებით განისაზღვრება: 1997 წელს დოლი დაიბადა, როგორც ითქვა, კლონირების შედეგად, ანუ იყო იმ ცხვრის ზუსტი გენეტიკური ასლი, რომლის დნ მჟავა გამოიყენეს კლონირებისათვის (დაწვრილებით იხ. ქვემოთ). უნდა აღინიშნოს, რომ ძუძუმწოვრების ჩვეულებრივი (სქესობრივი) გამრავლებისას შთამომავლობა არასდროს არაა თავისი მშობლების ზუსტი ასლი. ეს სტატია მიმოიხილავს კლონირების ექსპერიმენტების მოკლე ისტორიას (რომლებიც სწორედ დოლის დაბადებით დაგვირგვინდა) და ამ ექსპერიმენტების შედეგად წამოჭრილ პრობლემებს.

ნახ. 1: დოლი (Dolly) - ცხვარი, რომელიც დაიბადა როზლინის ინსტიტუტში (Roslin Institute, Scotland) ზრდასრული ცხვრის სარძევე ჯირკვლებიდან მიღებული ბირთვის ტრანსპლანტაციის შედეგად.
დასაწყისში რამდენიმე მნიშვნელოვანი განსაზღვრებაა აუცილებელი: უჯრედი ყოველი ცოცხალი ორგანიზმის ერთეულია, რომელიც ორგანიზმის ყველა თვისების განმსაზღვრელ ინფორმაციას ატარებს. ეს დებულება ვრცელდება როგორც ერთუჯრედიან ორგანიზმებზე, ასევე მრავალუჯრედიანებზეც, რომლებიც შემადგენლობაში შესაძლოა არსებობდეს რამდენიმე ასეულიდან (102) რამდენიმე ათეულ მილიარდამდე (1014) უჯრედი. დნ მჟავა, ანუ დნმ, ორგანიზმის გენეტიკური მასალაა, რომელიც ყველა ნიშან-თვისების განმსაზღვრელ კოდს შეიცავს. ნებისმიერი ორგანიზმის თითოეულ უჯრედში არსებული დნმ ამ ორგანიზმის ზრდისა და განვითარების სრულ გენეტიკურ კოდს შეიცავს. ორგანიზმების მრავალფეროვნება სწორედ მათი დნმ-ის (გენეტიკური კოდების) განსხვავებებით განისაზღვრება. ბირთვი უჯრედის ნაწილია, რომელიც დნ მჟავას შეიცავს, ციტოპლაზმა კი - ბირთვის გარეთ მყოფი უჯრედის შიგთავსი. კლონები ისეთი ორგანიზმებია, რომლების დნმ ერთმანეთის იდენტურია. კლონირება კი ასეთი ორგანიზმების მიღების პროცესია. ზოგიერთ შემთხვევაში, კლონირება ასევე გამოიყენება გენეტიკური ინჟინერიის ერთ-ერთი მეთოდის აღსაწერად, როცა ხდება დნ მჟავის ერთი ფრაგმენტის მიერთება მეორესთან, ამ ფრაგმენტის სტაბილური შენარჩუნებისა და გამრავლების მიზნით. ამ სტატიაში, ტერმინი „კლონირება“ მხოლოდ პირველი მნიშვნელობით (იდენტური ორგანიზმების მიღება) იქნება გამოყენებული.
კლონირების საფუძვლები მეცხრამეტე საუკუნის ბიოლოგიაში
როგორ ხდება უჯრედების სპეციალიზაცია განვითარების დროს, ანუ ერთი საწყისი უჯრედიდან (განაყოფიერებული კვერცხუჯრედიდან) სხვადასხვა მრავალნაირი ფუნქციის მქონე (კანის, ღვიძლის, ტვინის, ძვლის) უჯრედის წარმოქმნა? XIX საუკუნის მოერე ნახევარში არსებობდა ჰიპოთეზა, რომ განაყოფიერებული კვერცხუჯრედის, ანუ ზიგოტის, გაყოფის შედეგად წარმოქმნილ უჯრედებს შორის ხდება მემკვიდრეობითი ინფორმაციის დანაწევრება. ამ ჰიპოთეზის თანახმად, უჯრედების სპეციალიზაცია (ანუ მათი დიფერენციაცია) განისაზღვრება მემკვიდრეობითი ინფორმაციის იმ ნაწილით, რომელიც მათ გადაეცათ საწყისი უჯრედიდან. XIX საუკუნეშივე ამფიბიების ჩანასახებზე ჩატარებულმა ექსპერიმენტებმა უარჰყო ეს ჰიპოთეზა და აჩვენა, რომ მემკვიდრეობითი ინფორმაცია არ მცირდებოდა ან იყოფოდა ზიგოტიდან წარმოქმნილ უჯრედებში, არამედ უცვლელი სახით გადაეცემოდა თითოეულ შვილეულ უჯრედს. 1894 წელს ჟაკ ლობმა (Jacques Loeb) აღმოაჩინა, რომ შესაძლებელი იყო განაყოფიერებული კვერცხუჯრედის „გადანასკვა“ იმგვარად, რომ ბირთვი კვერცხუჯრედის ერთ ნაწილში მოექცეოდა. „ბირთვიანი“ ნაწილის რამდენიმე გაყოფის შემდეგ წარმოქმნილი ერთ-ერთი უჯრედის ბირთვი ზოგჯერ უბრუნდებოდა „უბირთვო“ ნაწილს. ამის შედეგად ეს ნაწილიც გაყოფას იწყებდა და ნორმალურ ჩანასახს წარმოქმნიდა. მსგავსი ცდა ჩატარდა 1914 წელს ჰანს სპემანის (Hans Spemann) მიერ. ეს ექსპერიმენტები შესაძლოა ბირთვის ტრანსპლანტაციის (ერთი უჯრედიდან მეორეში გადატანის) და, მაშასადამე, კლონირების პირველ წარმატებულ ცდად ჩაითვალოს.
ბირთვის ტრანსპლანტაციის თეორიული საფუძვლებისა და ტექნოლოგიის განვითარება
1952 წელს რობერტ ბრიგსმა და თომას კინგმა (Robert Briggs, Thomas J. King) აჩვენეს, რომ ბაყაყის ჩანასახიდან მიღებული ბირთვის გადატანის შედეგად უბირთვო უჯრედებში ხდებოდა თავკომბალების განვითარება. როგორც უბირთვო უჯრედების მიღება, ასევე ბირთვის გამოცალკევება და მისი ტრანსპლანტაცია ხდებოდა ავტორების მიერ შემუშავებული მიკროქირურგიული მეთოდებით. შემდეგ აღმოჩნდა, რომ ჩანასახის განვითარებასთან ერთად, ბირთვების დიფერენცირების პოტენციალი მცირდებოდა, ანუ ძნელდებოდა ასეთი ბირთვების ტრანსპლანტაციის შედეგად ორგანიზმის ნორმალური განვითარება. ამ დაბრკოლების გადალახვა ნაწილობრივ მოხერხდა ტრანსპლანტაციის პროცედურაში რამდენიმე ცვლილების შეტანით (ტემპერატურის დაწევა ბირთვის მიმღები უჯრედის ზრდის შენელებისათვის და გარკვეული ქიმიური რეაგენტის დამატება, რომელიც დიფერენციაციის ხელისშემწყობი ცილების მოქმედებას ზღუდავდა). ზრდასრული ამფიბიების ორგანიზმებისაგან კი საერთოდ ვერ მოხერხდა ისეთი ბირთვების მიღება, რომელთა ტრანსპლანტაციის შედეგად ნორმალური ჩანასახის განვითარება იქნებოდა შესაძლებელი. მოგვიანებით ასევე გაირკვა, რომ ტრანსპლანტაციის წარმატებისათვის ბირთვები არა უჯრედების გაყოფის დროს, არამედ მათი „დასვენების სტადიაში“ უნდა შეგროვდეს. ამფიბიებზე მიღებული შედეგები საფუძვლად დაედო ბირთვის ტრანსპლანტაციის ექსპერიმენტებს სხვა ორგანიზმებში: 1960-იან წლებში უკვე ჩატარდა მწერებისა და თევზების კლონირების რამდენიმე წარმატებული ცდა, 1980-იან წლებში კი ეს ექსპერიმენტები თაგვებზეც შესრულდა. სხვა ძუძუმწოვრების კლონირების ცდებიც წარმატებით სრულდებოდა იმ შემთხვევაში, თუ ბირთვი ჩანასახის ადრეულ სტადიაზე იყო მიღებული: ორგანიზმის განვითარების გვიან სტადიაზე კლონირების ეფექტურობა საგრძნობლად კლებულობს. სწორედ ამ თვალსაზრისით იყო შესანიშნავი დოლის დაბადება - პირველი კლონირებული ცხვრისა, რომლის შესაქმნელად აუცილებელი გენეტიკური მასალა (ბირთვი) მიღებული იყო ზრდასრული ცხოველის - 6 წლის ცხვრის სარძევე ჯირკვლებიდან.
რითი გამოირჩეოდა „დოლის ექსპერიმენტი“?
როგორც ზემოთ აღვნიშნეთ, ზრდასრული ორგანიზმიდან მიღებული ბირთვის ტრანსპლანტაციის უნარი მეტად შეზღუდულია. მეორე მხრივ, „დასვენების სტადიაში“ მყოფი უჯრედებიდან მიღებული ბირთვისათვის ეს პროცესი გაცილებით ეფექტურია. იენ უილმუთი (Ian Wilmut) და მისი კოლეგები როზლინის ინსტიტუტში (Roslin Institute, Scotland) სწორედ ამ ორ გარემოებას განიხილავდნენ ცხვრის კლონირების სტრატეგიის შემუშავებისას. მათ ივარაუდეს, რომ წარმატებული ტრანსპლანტაციისათვის აუცილებელი იყო, რომ ამ პროცესში მონაწილე ორივე უჯრედი (როგორც უბირთვო, ასევე ის, რომელიც ბირთვის მისაღებად იყო არჩეული) ერთსა და იმავე სტადიაში ყოფილიყო. ამასთან ერთად, 6 წლის ცხვრის უჯრედები საკვები ნივთიერებებით ღარიბ გარემოში მოათავსეს. სავარაუდოდ, ამის შედეგად ზრდასრული უჯრედის ბირთვში არსებულ დნ მჟავას ის ცილები მოშორდა, რომლებიც მის „სპეციალიზაციას“ უზრუნველჰყოფდნენ. ამ მნიშვნელოვანი მოდიფიკაციების შედეგად (და სუროგატი ცხვრის ორგანიზმში განვითარების შემდეგ) 1996 წლის ივლისში დაიბადა დოლი.

ნახ. 2: იენ უილმუთი (Ian Wilmut) და დოლი.
დოლი მხოლოდ ერთი წარმატებით დაგვირგვინებული შედეგი იყო იმ 277 ტრანსპლანტირებული უჯრედიდან, რომელიც უილმუთმა და მისმა თანამშრომლებმა მიიღეს. ამ გარემოებამ არაერთი ეჭვი თუ სკეპტიკური შეკითხვა გამოიწვია, რომლებიც, პირველ რიგში, ტრანსპლანტირებული ბირთვის იდენტობას ეხებოდა: მრავალი ექსპერტი ვარაუდობდა, რომ ეს ერთადერთი უჯრედი არა სპეციალიზირებული, არამედ ღეროვანი (არადიფერენცირებული) უჯრედი (stem cell) იყო, და დოლის დაბადება სწორედ ამით აიხსნებოდა. სკეპტიციზმის მეორე საფუძველი 6 წლის შემდეგ - დოლის შედარებით ნაადრევი დაღუპვის გამო წარმოიშვა: ამბობდნენ, რომ დოლის დნ მჟავა არა 6, არამედ 12 წლისა იყო (მისი ბირთვის ასაკის გათვალისწინებით), და მისი სიკვდილი „ნაადრევი დაბერებით“ იყო გამოწვეული. დაბერების დაჩქარების სავარაუდო მექანიზმი ქრომოსომების (დნმ-ის დამოუკიდებელი ფრაგმენტების) დამოკლებაა და ეს პროცესი, შესაძლოა, უკვე დაწყებული იყო ახალდაბადებული დოლის უჯრედებში. ოპონენტები ასევე აღნიშნავდნენ, რომ დოლის „ზუსტი გენეტიკური ასლის“ სახელით მოხსენიება მთლიანად არ შეესაბამებოდა სინამდვილეს, რადგან დოლის და მის დედას მხოლოდ ძირითადი გენეტიკური მასალა (რომელიც ბირთვშია მოთავსებული) ჰქონდათ იდენტური, ციტოპლაზმა და მასში შემავალი დნმ კი - სხვადასხვანაირი. აქ აუცილებელია აღვნიშნოთ, რომ ეს კრიტიკა ზოგადად ბირთვის ტრანსპლანტაციის მეთოდს და მასთან დაკავშირებულ ტერმინოლოგიას ეხება და არა მხოლოდ დოლის კლონირების ექსპერიმენტს.

ნახ. 3: Idaho Gem („აიდაჰოს [ძვირფასი] თვალი“) - პირველი კლონირებული ჯორი. იგი წარმატებით მონაწილეობდა დოღში და რამდენიმე საპრიზო ადგილიც აქვს აღებული. 2006 წელს ჩატარდა პირველი დოღი, რომელშიც „ჩვეულებრივ“ ჯორებთან ერთად, კლონირებული ჯორიც მონაწილეობდა.
უილმუთი და მისი კოლეგები ამ მოსაზრებებს უსაფუძვლოდ თვლიან: ღეროვანი უჯრედის მოხვედრა სარძევე ჯირკვალში, მათი განმარტებით, თითქმის შეუძლებელი იყო. რაც დნმ-ის დაბერებას შეეხება, ამ პროცესის დაჩქარების საბუთი კლონირებულ ცხოველებში (და კერძოდ, დოლის ორგანიზმში) ჯერ არ არსებობს. თანაც, ფილტვების ინფექციის შედეგად 6 წლის ასაკში ცხვრის დაღუპვა უჩვეულო სულაც არაა. კლონირებულ ცხოველებში ხშირად იჩენს თავს მნიშვნელოვანი (თუმცა არა სიცოცხლესთან შეუთავსებადი) დეფექტები, მაგრამ დოლი სავსებით ნორმალურად ვითარდებოდა. ამრიგად, როგორც დოლის „წარმოშობის“, ასევე მისი დაღუპვის მიზეზების შესახებ აზრთა სხვადასხვაობა ჯერ კიდევ არსებობს.
კლონირების პერსპექტივები და მასთან დაკავშირებული პრობლემები
დოლის დაბადებას მოჰყვა კატის, ძაღლის, ბოცვრის, ღორისა და ძროხის ბირთვის ტრანსპლანტაციის წარმატებული ექსპერიმენტები. 2001 წელს კლონირებისათვის ასევე წარმატებით გამოიყენეს დაღუპული ცხოველის (გარეული ცხვრის - მუფლონის) უჯრედიც კი. 2003 წელს მოხდა ცხენისებრთა ოჯახის წარმომადგენლის პირველი კლონირება. კლონირებული იყო ჯორი, რაც ასევე ბუნებრივად სტერილური ცხოველის კლონირების პირველი შემთხვევაა. ამ ექსპერიმენტში, „დაბერებული“ დნ მჟავის პრობლემის თავიდან ასაცილებლად, ბირთვი 45 დღის ასაკის ჩანასახიდან იყო მიღებული. აქვე უნდა აღინიშნოს, რომ წმინდასისხლიან/სადოღე ცხენებზე ცხოველთა კლონირების გავრცელების შესაძლებლობამ დიდი პროტესტი გამოიწვია: კლონირების ოპონენტები ამტკიცებდნენ, რომ ამ მეთოდის დანერგვა ძირს გამოუთხრიდა ცხენების სელექციის ხელოვნებას და დოღს იდენტური კლონების შეჯიბრებამდე დაამდაბლებდა.
პრიმატების კლონირებას უდიდესი სამედიცინო მნიშვნელობა აქვს, მაგ., გენეტიკურად იდენტური მაიმუნების მისაღებად, რაც შიდსის საწინააღმდეგო ნაქცინის მისაღებად შესანიშნავი შენაძენი იქნებოდა. ჯერ-ჯერობით, ორი კლონირებული რეზუს-მაკაკას გენეტიკური მასალა ჩანასახის განვითარების ადრეული ეტაპებზეა მიღებული. ზრდასრული ცხოველის უჯრედებიდან მიღებული ბირთვის ტრანსპლანტაცია ამ შემთხვევაშიც რთული პრობლემაა.
ბუნებრივია კითხვა, შესაძლო იქნება თუ არა ადამიანების კლონირება? ეთიკური საკითხების გარდა, რომელიც ასეთ შემთხვევაში თავისთავად წამოიჭრება, არსებობს მეთოდოლოგიური პრობლემებიც: თვით იენ უილმუთი აღიარებს, რომ დოლის წარმატებული მაგალითის გარდა, არსებობს კლონირების არაერთი დამაფიქრებელი შედეგი (მაგ., დოლის დაბადებიდან ცოტა ხნის შემდეგ კლონირებული ცხვარი, რომელის არასრულყოფილად განვითარებული ფილტვებით დაიბადა; ხშირ შემთხვევებში კლონირების გზით დაბადებულ ცხოველებს არაბუნებრივად დიდი ზომის შინაგანი ორგანოები ახასიათებთ, და ა.შ.). სწორედ ასეთი შემთხვევების გამო მეცნიერების დიდ უმრავლესობას დაუშვებლად მიაჩნია ადამიანის კლონირების დაწყება არსებული თეორიული ცოდნისა თუ ექსპერიმენტული მეთოდოლოგიის საფუძველზე. ჯერ კიდევ 2001 წელს ამერიკის ეროვნულ აკადემიაში ამ თემაზე ჩატარებული კონფერენციის თითქმის ყველა მონაწილემ აღიარა, რომ ადამიანის კლონირება უსაფრთხო მეთოდად ვერ იქნება მიჩნეული. თუმცა კონფერენციის მონაწილე მეცნიერთა ნაწილი აცხადებდა, რომ ადამიანის გამრავლების შესახებ დაგროვილი ცოდნა მნიშვნელოვნად აღემატება ინფორმაციას ამ პროცესის შესახებ სხვა ძუძუმწოვრებში. ამიტომ, მათი აზრით, ადამიანის კლონირებისას შესაძლო იქნება ცხვრებისა და მღრღნელების შემთხვევაში მიღებული არასასურველი შედეგების თავიდან აცილება. ადამიანის კლონირების მომხრეთა მთავარი არგუმენტი კი ადამიანის უფლებებს ეხება, რადგან „ნებისმიერი ადამიანის ფუნდამენტური უფლებაა გამრავლდეს იმ გზით, რომელსაც იგი აირჩევს“.
დასკვნა
დოლის კლონირება უდაოდ დიდ მეცნიერულ მიღწევად უნდა ჩაითვალოს, რადგან ამ ექსპერიმენტის ავტორებმა თავი გაართვეს უაღრესად რთულ ამოცანას როგორც თეორიული თვალსაზრისით, ასევე მრავალსაფეხურიანი ცდის თითქმის უნაკლო შესრულების მხრივ. ცხოველების (და კერძოდ, ძუძუმწოვრების) კლონირების პრაქტიკული მნიშვნელობა ბიოტექნოლოგიისა და მედიცინისათვის ეჭვგარეშეა. ამასთან ერთად, ადამიანის კლონირება, რაშიც იგულისხმება კონკრეტული პიროვნების ზუსტი გენეტიკური ასლის შექმნა ბირთვის ტრანსპლანტაციის საფუძველზე, მრავალ ეთიკურ და მეთოდოლოგიურ პრობლემას ქმნის, და მათ გადაჭრას ალბათ კიდევ არაერთი წელი დასჭირდება.
გელა თევზაძე
24 საათი 27.02.11
|
|
5 გენმოდიფიცირებული ორგანიზმები: რეალური საფრთხე თუ უსაფუძვლო შიში? |
▲ზევით დაბრუნება |
გენმოდიფიცირებული ორგანიზმების (ქვემოთ - GMO) გავრცელება და მათი საკვებად გამოყენების მართებულობა ბოლო წლებში საკმაოდ მწვავე პრობლემად იქცა. GMO-ს მომხრეები ახალი ორგანიზმების შთამბეჭდავ უპირატესობებზე მიუთითებენ, მოწინააღმდეგეები კი როგორც ტრადიციული სახეობების გაქრობის საშიშროებას, ასევე „ხელოვნურად შექმნილი საკვების“ შეუფერებლობას აღნიშნავენ. ამ სტატიაში განვიხილავთ როგორც GMO-ს წარმოშობის ისტორიასა და მათ განვითარებას თანამედროვეობამდე, ასევე მათ მომხრეთა და მოწინააღმდეგეთა არგუმენტებს და მათ მართებულობას.

1. GMO-ს საწინააღმდეგო ემბლემა.
რა არის GMO? გენეტიკურად მოდიფიცირებული (ან გენმოდიფიცირებული) ორგანიზმი ისეთი ორგანიზმია, რომლის გენეტიკური მასალა (დნ მჟავა) შეცვლილი იქნა გენეტიკური ინჟინერიის მეთოდების გამოყენებით. ეს მეთოდები ასევე ცნობილია რეკომბინანტული დნ მჟავის ტექნოლოგიის კრებითი სახელით. რეკომბინანტული დნ მჟავის ტექნოლოგია საშუალებას იძლევა, სხვადასხვა ორგანიზმების დნ მჟავის მოლეკულების კომბინაციით შეიქმნას ახალი გენები. ეს ახალი გენები (რომლებიც დნ მჟავას ფრაგმენტებია) შემდეგ შეჰყავთ რომელიმე ორგანიზმში, რის შედეგადაც ეს ორგანიზმი იძენს შეცვლილ ან სრულიად ახალ თვისებებს. ტრანსგენული ორგანიზმები, რომლებიც GMO-ს კერძო შემთხვევაა, შეიცავს რამდენიმე სხვადასხვა ორგანიზმიდან მიღებულ დნ მჟავას (და, შესაბამისად, გენებს).
მოკლე ისტორიული მიმოხილვა
რადგან GMO-ს მიღების პრინციპი ორგანიზმისათვის ახალი გენეტიკური მასალის შეძენას ეფუძნება, მათი ისტორია გენეტიკურ ინჟინერიასთან ერთად იწყება. პირველი რეკომბინანტული ბაქტერია შექმნეს 1973 წელს, როცა მიიღეს ნაწლავის ჩხირის (E. coli) ბაქტერია, რომელშიც განსხვავებული ბაქტერიის (სალმონელას) გენი იყო „ჩაშენებული“. ახალი მეთოდის პერსპექტივებსა და შესაძლო პრობლემების თემაზე კალიფორნიაში 1975 წელს ჩატარდა ასილომარის კონფერენცია. ამ კონფერენციაზე მიღებული რეკომენდაციის შესაბამისად, კვლევები რეკომბინანტული დნ მჟავის ტექნოლოგიაში უნდა მოქცეულიყო მთავრობის კონტროლის ქვეშ - თუნდაც იმ დრომდე, როცა ამ ტექნოლოგიის უსაფრთხოება ეჭვქვეშ აღარ იქნებოდა. უკვე 1978 წელს კი კომპანია „გენენტექმა“ (Genentech) შექმნა E. coli, რომელშიც ადამიანის ინსულინის გენი იყო „ჩაშენებული“ და რომელიც, მაშასადამე, ადამიანის ინსულინს გამოიმუშავებდა. ადვილი მისახვედრია, რომ ეს ბაქტერია GMO-ს ერთ-ერთ პირველ მაგალითად შეიძლება ჩაითვალოს.
1986 წლისათვის შექმნილი იყო ე.წ. „ყინულ-მინუსოვანი“ (ice-minus) ბაქტერია, რომელსაც, ჩანაფიქრის მიხედვით, შეეძლო მცენარეთა მოყინვის პრობლემა გადაეჭრა. ამ ბაქტერიის ზედაპირზე (მისი „არაგენმოდიფიცირებული“ ვარიანტისაგან განსხვავებით) აღარ წარმოიქმნება ერთ-ერთი ცილა, რომელიც ჩვეულებრივ ყინულის ნაწილაკების დაგროვებისათვის ხელსაყრელ პირობებს ქმნის. ფოთლების ზედაპირის დაფარვას „ყინულ-მინუსოვანი“ ბაქტერიებით მცენარეები მოყინვისაგან უნდა გადაერჩინა. ეს პროექტი წარუმატებლად დასრულდა, რადგან ბაქტერიების საველე გამოცდისას წარმოიქმნა აზრი, რომ „ყინულ-მინუსოვანი“ ბაქტერიების გავრცელება კლიმატზე და, კერძოდ, წვიმის სიხშირეზე მოახდენდა გავლენას.
აშშ-სა და კანადაში გენმოდიფიცირებული (GM) მცენარეების პირველი მცირე გამოცდები ჯერ კიდევ 1980-იან წლებში დაიწყო. 1990-იანი წლების შუაში კი უკვე ფართომასშტაბიანი გამოცდებისა და კომერციული წარმოების ნებართვა იყო მიღებული. იმ დროიდან, GM მცენარეების გამოყენება აშშ და კანადის ფერმერების მიერ ყოველწლიურად იზრდებოდა.

2. მომხმარებელთა დამოკიდებულება GM-საკვების მიმართ 2010 წელს (NM-Incite-ის მიხედვით): 59%25 უარყოფითი, 8%25 დადებითი, ყველა დანარჩენს ან მეტ-ნაკლებად ნეიტრალური დამოკიდებულება აქვს, ან „არანაირი“ აზრი არ გააჩნია ამ თემაზე.
GMO-ს გამოყენება
GM ორგანიზმები გამოიყენება როგორც ბიოლოგიურ და სამედიცინო კვლევაში, ასევე ფარმაკოლოგიურ წარმოებასა და აგრობიზნესში. ამ მიმოხილვაში GMO-სა და აგრობიზნესის ურთიერთქმედების საკითხს დაწვრილებით განვიხილავთ.
ამ დროისათვის, GMO-ს ყველაზე წინააღმდეგობრივი, და ამასთანავე ყველაზე პოპულარული მაგალითი კომპანია „მონსანტო“-ს (Monsanto, USA) საკუთრება - მარცვლეულის კულტურები, რომლებსაც ახასიათებს ჰერბიციდებისადმი მდგრადობა, პესტიციდური თვისებები, ან ორივე ამ თვისებათაგანი. „მონსანტოს“ GMO მარცვლეული მსოფლიოში 1 მილიონ კვადრატულ კილომეტრზეა კულტივირებული. თუმცა, მეტად შთამბეჭდავი გავრცელებისა და გავლენის არეალის ყოველწლიური ზრდის მიუხედავად, „მონსანტოს“ პატენტი თავისი პროდუქტების უპირატეს გავრცელებაზე 2014 წელს ამოიწურება, და სხვა კომპანიებში შექმნილი GM კულტურების გავრცელებისათვის უკეთესი პირობები შეიქმნება. ერთ-ერთი შეფასებით, 2015 წლისათვის ახალშექმნილი GM მცენარეების 40%25 „აზიური წარმოშობისა“, ანუ აზიაში განლაგებული კომპანიების პროდუქტი იქნება.
GMO-ს წარმატებული გამოყენების მაგალითები ძუძუმწოვარ ცხოველებშიც არსებობს. 2009 წელს ა.შ.შ.-ში მოიწონეს და დაამტკიცეს ადამიანის ანტიკოაგულანტის (სისხლის შედედების ხელისშემშლელი ნაერთის) წარმოება, რომელსაც თხის ორგანიზმში „ჩაშენებული“ გენი წარმოქმნის და რომლის გამოყოფა ამგვარი თხის რძიდანაა შესაძლებელი.
ურთიერთსაწინააღმდეგო შეხედულებები GMO-ს თაობაზე
ზოგადი მოსაზრებები: ზოგიერთი აზრის თანახმად, GMO წარმოადგენს უხეშ ჩარევას ბიოლოგიური პროცესების ბუნებრივ მიმდინარეობაში, რაც ევოლუციის კანონების მნიშვნელოვანი დარღვევაა. ასევე არსებობს ეჭვი, რომ თანამედროვე მეცნიერებას არ ძალუძს სრულყოფილად განჭვრიტოს გენეტიკური ინჟინერიის ყველა შესაძლო (უარყოფითი) შედეგი. მეორე მხრივ, არსებობს საწინააღმდეგო მოსაზრებაც, რომელსაც ამ სტრიქონების ავტორიც იზიარებს: GMO-ს არსებობა განიხილება, როგორც იმ პროცესის გაგრძელება, რომელიც ადამიანების მიერ საუკუნეების მანძილზე მარცვლეულთა (და, ზოგადად, მცენარეთა და ცხოველთა ჯიშების) გაუმჯობესებაში მდგომარეობდა. მართლაც, გენეტიკური ინჟინერიის მეთოდების მიმართ გაუცნობიერებელ შიშს თუ უგულებელვყოფთ, GMO-ს შექმნის პროცესი არაფრით განსხვავდება მცენარეთა და ცხოველთა ხელოვნური სელექციისაგან, რომლის არსი სასარგებლო თვისებების გამორჩევა და გაძლიერებაა.
გავლენა „კვებით ჯაჭვზე“: GMO-ს მოწინააღმდეგეთა აზრით, ასეთი ორგანიზმების საკვებში გამოყენება ხელს უწყობს ახალი სახის ალერგენებისა და ანტიბიოტიკებისადმი მდგრადობის განმსაზღვრელი გენების გავრცელებას. ჯერ-ჯერობით, ამგვარი ბრალდებები საფუძველს მოკლებულია, რადგან არცერთი გამოქვეყნებული კვლევა არ ადასტურებს ჯანმრთელობის გაუარესებას GM მცენარეების პროდუქტებით კვების შედეგად. თანაც, ალერგენების წარმოქმნა გამორიცხული არაა „კლასიკური“ ხელოვნური სელექციის შედეგადაც. რაც შეეხება ანტიბიოტიკებისადმი მდგრადობის განმსაზღვრელ გენებს, GM ტექნოლოგიის საწყის ეტაპებზე ისინი მართლაც გამოიყენებოდა „გენეტიკურ ნიშნულად“ რეკომბინანტული დნ მჟავის შექმნის პროცესში, მაგრამ თანამედროვე - უფრო დახვეწილ - მეთოდებში ამ გენების გამოყენებას გვერდს უქცევენ: ძირითადი მიზეზი იმ (უაღრესად მცირე) შანსის გამორიცხვაა, რომ ასეთი გენები ადამიანის ორგანიზმში მოხვედრისას „ადგილობრივ“ მიკროორგანიზმებში (მაგალითად, მომნელებელი სისტემის მიკროფლორაში) შეაღწევდნენ და მათ გენეტიკურ ცვლილებას გამოიწვევდნენ. გამონათქვამი, რომ GM-საკვები „არაბუნებრივია და, მაშასადამე, არაა უსაფრთხო“ (unnatural and therefore unsafe) უფრო ემოციებზეა დაფუძნებული, ვიდრე ფაქტებსა და მსჯელობაზე. მიუხედავად ამისა, ამ შეხედულებების გავლენით მრავალ სახელმწიფოში მიღებულია კანონები, რომლების მიხედვითაც ადამიანთა კვებისათვის განკუთვნილი ყველა ახალი ორგანიზმის უსაფრთხოების შემოწმება აუცილებელია. რასაკვირველია, ამგვარი კანონების არსებობა დადებითი მოვლენაა, თუნდაც იმ მოსაზრებით, რომ ასეთი შემოწმების შედეგად დაგროვილი „დადებითი შედეგები“ საზოგადოებაში GMO-ს მიმართ არსებულ უნდობლობას შეამცირებს.
GM და „ბუნებრივი“ ორგანიზმების ურთიერთქმედება: GMO-ს კრიტიკის ერთ-ერთ მნიშვნელოვან ასპექტს წარმოადგენს არგუმენტი სახეობათა ბუნებრივი მრავალფეროვნების შემცირების შესახებ. ეს შემცირება, ამ არგუმენტის თანახმად, GMO-ს გავრცელების გარდაუვალი შედეგია: ჯვარედინი დამტვერვის შედეგად ხდება GM მცენარეების შეჯვარება ადგილობრივ ჯიშებთან, რაც ამ უკანასკნელთა თანდათანობით გაქრობას იწვევს და ართულებს GM-აგრობიზნესის თანაარსებობას ტრადიციულ მეურნეობასთან. ამ შემთხვევაშიც, ეს პროცესი არაფრით განსხვავდება ბუნებაში არსებულ შეჯვარებისაგან ჯვარედინი დამტვერვის მქონე მცენარეებს შორის, რისი შედეგიც ხშირად ახალი ჰიბრიდების წარმოქმნა და ზოგჯერ ტრადიციული ჯიშების გავრცელების არეალის შემცირება ხდება. ამრიგად, ეს არგუმენტიც GMO-სათვის „სპეციფიურ ბრალდებად“ არ გამოდგება და მხოლოდ GM კულტურების ფართო გავრცელების (იხ. ზემოთ) შესაძლო შედეგების პროგნოზისათვის თუ გამოდგება.
GMO კულტურების სტაბილურობისა და GM მცენარეებზე „საავტორო უფლების დაცვის“ საკითხი: კომერციული მოსაზრებით, GM მცენარეთა უმეტესობის სათესლე მასალა ე.წ. „პირველი თაობის“ (F1) ჰიბრიდია, რომლის შემდეგ თაობაში (F2) მომგებიანი თვისებების განმსაზღვრელი გენები ერთმანეთისაგან „ითიშებიან“, ანუ არაერთგვაროვან შთამომავლობას წარმოქმნიან. ამის გამო, ფერმერები იძულებულნი არიან, ყოველ წელს შეისყიდონ კომპანიისაგან (მაგ., „მონსანტო“-საგან) F1 სათესლე მასალა. ეს, უმეტეს შემთხვევაში, უფრო მომგებიანია ფერმერებისათვის, ვიდრე არაერთგვაროვანი F2 (F3, და ა.შ.) თესლის გამოყენება, რაც მოსავლიანობის შემცირებასა და ინსექტიციდებსა და პესტიციდებზე დანახარჯის გაზრდას გამოიწვევს.
GM კულტურებზე მონოპოლიის შენარჩუნების კიდევ ერთი ხერხია „მკვლელი“ („Terminator“) და „მოღალატე“ („Traitor“) გენების ჯერ-ჯერობით დაუმკვიდრებელი ტექნოლოგია. ორივე მათგანის მიზანია, შეაჩეროს GMO-ს გავრცელება ჯვარედინი დამტვერვით (იხ. ზემოთ) და, ასევე, უფრო „დამაჯერებლად“ აიძულოს ფერმერები, ყოველ წელს შეისყიდონ სათესლე მასალის ახალი პარტია კომპანიისაგან. „Terminator“ გენის ამუშავების შედეგად კომპანიისაგან შესყიდული მარცვლისაგან აღმოცენებული ყველა მცენარე უნაყოფოა, ანუ მათგან მიღებული მოსავალი მხოლოდ საკვებად გამოდგება და არა სათესლე მასალად. „Traitor“ გენის არსებობის შემთხვევაში, GM თვისებების გამოვლენა მხოლოდ მაშინ მოხდება, თუ სათესლე მასალა გარკვეული (უეჭველია, იმავე კომპანიის კუთვნილი) რეაგენტითაა დამუშავებული. ორივე მეთოდი, როგორც ითქვა, ჯერ კიდევ დამუშავებისა თუ დახვეწის სტადიაშია.
და ბოლოს, მოვიყვან სხვადასხვა ტრანსგენული/GM ორგანიზმის კიდევ რამდენიმე მაგალითს: 2009 წელს იაპონელმა მეცნიერებმა შესძლეს გენის გადატანა მარმოზეტის სახეობის მაიმუნში და პირველი სტაბილური ტრანსგენული პრიმატები მიიღეს. უკვე ნახსენები ინსულინის გარდა, რეკომბინანტული ბაქტერიები გამოიყენება ადამიანის ზრდის ჰორმონის გამომუშავებისათვის. GM თევზები (მაგ. ორაგული) გენეტიკური მოდიფიკაციის შედეგად დიდი რაოდენობით გამოიმუშავებენ საკუთარი ზრდის ჰორმონს, რაც მათი ზრდის მნიშვნელოვან დაჩქარებას იწვევს. ასევე თევზებში მიღებულია რამდენიმე სხვადასხვა ფერის ნათების (ფლუორესცენციის) უნარის მქონე ზებრასებრი თევზი (zebrafish). ფლუორესცენცია მედუზის ორგანიზმიდან გამოყოფილი მწვანე ფლუორესცირებადი ცილითაა (Green Fluorescent Protein, GFP) გაპირობებული. ასეთი „GloFish“ თევზები ერთ-ერთი პირველი GM ცხოველებია, რომელთა შეძენა უკვე შესაძლებელია.

3. GloFish - გენეტიკურად მოდიფიცირებული ზებრასებრი თევზები, რომლებშიც მედუზიდან გამოყოფილი ფერადი ნათების გამომწვევი გენია გადატანილი.
დასკვნა
GM ორგანიზმების შექმნა მეცნიერებისა და ზოგადად კაცობრიობის ერთ-ერთ უმნიშვნელოვანეს მიღწევად უნდა ჩაითვალოს. GMO-ს გავრცელების უდაო დადებით მხარეებია მაღალპროდუქტიული და დაავადებებისადმი ნაკლებად მგრძნობიარე კულტურების გავრცელება, ზოგიერთ ქვეყანაში საკვების ნაკლებობის პრობლემის შესაძლო გადაჭრა ასეთი გავრცელების შედეგად, აგრობიზნესისათვის ოპტიმიზირებული და მომგებიანი კულტურების დამკვიდრება. GMO-სთან დაკავშირებული პრობლემების უმეტესობა ტექნოლოგიურ და მეთოდოლოგიურ საკითხებთანაა დაკავშირებული და ამ საკითხების გადაჭრასთან ერთად, იმედია, პრობლემებიც გაქრება. GM-საკვების მიმართ არსებული უნდობლობაც მომავალში უნდა შემცირდეს ასეთი ორგანიზმების რაოდენობის ზრდისა და მათი უსაფრთხოების დამამტკიცებელი მაგალითების დაგროვების შედეგად.
გელა თევზაძე
24 საათი 13.03.11
|
|
6 სინთეტური ბიოლოგია: ძველი ნაწილებიდან აწყობილი ახალი სისტემები |
▲ზევით დაბრუნება |
ყველა ცოცხალი ორგანიზმი (და მისი შემადგენელი ერთეულები - უჯრედები) შესაძლოა წარმოვიდგინოთ სხვადასხვა მექანიზმის სახით, როგორც დიდი (მთელი ორგანიზმი), ასევე ძალიან პატარა (ცალკეული უჯრედი). ამ ანალოგიით დაინტერესდა სხვადასხვა სპეციალობის მრავალი მეცნიერი, რის შედეგადაც წარმოიქმნა ბუნებისმეტყველების ახალი დარგი - სინთეტური ბიოლოგია. სინთეტური ბიოლოგიის მიზნებია: ქიმიური სისტემების დაპროექტება, რომლების მუშაობის პრინციპი ცოცხალი ორგანიზმების მოქმედებას ემყარება; არსებული (ბუნებრივი) მოლეკულებისაგან ახალი ფუნქციონალური ნაერთების შექმნა; გამარტივებული (ანუ მხოლოდ სიცოცხლისათვის აუცილებელი კომპონენტების შემცველი) უჯრედების, ან ხელოვნური ეკოსისტემების აწყობა.
ბიოლოგიის ტრადიციული დარგების მიერ დაგროვილია ცოდნა უჯრედისა და მისი შემადგენელი ნაწილების ფუნქციებსა და ურთიერთქმედების შესახებ. ამ დახასიათებას შემდგომში ემატება ინფორმაცია მსგავსი სისტემების ევოლუციური „ნათესაობის“ შესახებ, ისევე როგორც მათი სტრუქტურის შესწავლა, რაც წარმოაჩენს კავშირს მოლეკულების ფორმასა და მათი ფუნქციის შორის. სინთეტური ბიოლოგია მიზნად ისახავს ბიოლოგიური სისტემების ოპტიმალურ დაპროექტებასა და შექმნას, ტრადიციულ დარგებში დაგროვილი ინფორმაციის გამოყენებით. არანაკლებ მნიშვნელოვანია, რომ ასეთი ხელოვნური სისტემები საშუალებას გვაძლევენ შევისწავლოთ მრავალი ბიოლოგიური კომპონენტის მოქმედება ახალ გარემოში. ეს კი, თავის მხრივ, ამდიდრებს ტრადიციული დარგების მიერ დაგროვილ ცოდნას.
სინთეტური ბიოლოგიის არანაკლებ მნიშვნელოვანი ასპექტია მისი კავშირი მათემატიკურ მოდელირებასთან. კომპიუტერული მოდელირების საშუალებით შესაძლებელია მრავალი ბიოლოგიური პროცესის (მაგ., გენებში კოდირებული ინფორმაციის „წაკითხვა“, ცილის სინთეზი, უჯრედის გაყოფა) მნიშვნელოვანი პარამეტრების განსაზღვრა, და ამ პარამეტრების ოპტიმალური სიდიდეების გამოანგარიშება, რომელთა საფუძველზე ამ პროცესის მართვაა შესაძლებელი.
წინამდებარე მიმოხილვაში სინთეტური ბიოლოგიის იმ ნაწილს განვიხილავთ, რომლის მიზანი ცალკეული უჯრედების შიგნით ახალი ბიოლოგიური სისტემების შექმნაა. „სინთეტური წრედი“ ან „სინთეტური ხელსაწყო“ გამოიყენება ისეთი სისტემის აღსანიშნავად, რომელიც შექმნილია რაიმე სასარგებლო ფუნქციის სტაბილურად და საიმედოდ შესრულებისათვის. ადვილი წარმოსადგენია, რომ განსაკუთრებულ ინტერესს იწვევს ასეთი სისტემების გამოყენება ბიოტექნოლოგიასა და გენურ თერაპიაში. მაგალითად, გენეტიკური „გადამრთველი“, რომელიც მოქმედებს უჯრედში შემავალი სიგნალების საპასუხოდ, შესაძლებელს გახდის სამკურნალო თვისებების მქონე ცილის სინთეზს დიდი ხნის განმავლობაში და მედიკამენტების მიღებას ააცილებს პაციენტებს. ასეთი გადამრთველი მოქმედებს ნათურის „ჩართვა-გამორთვის“ მექანიზმის მსგავსად: ნებისმიერ დროს გენი, რომელშიც კოდირებული ინფორმაციის საფუძველზე ხდება ამ ცილის სინთეზი, ან გამორთულია, ან ჩართული (ე.ი., არ არსებობს „შუალედური მდგომარეობა“, როცა ცილის სინთეზი სიგნალის მიღებაზე არაა დამოკიდებული, იხ, ქვემოთ).
გენეტიკური კონსტრუქციების აწყობისას სინთეტურ ბიოლოგიაში გამოიყენება რამდენიმე საინჟინრო პრინციპიც, მათ შორის ე.წ. ურთიერთცვლადი ფუნქციების სამ დონეზე არსებული იერარქია (three-level hierarchy of interchangeable functions). სიგნალის (მაგ., რომელიმე მცირე ზომის მოლეკულა) ამოცნობა ხდება სენსორის მიერ, შემდეგ ეს სიგნალი გადამუშავდება „წრედში“, რის შედეგადაც წარმოიქმნება სიგნალის საპასუხო მოქმედება. ბიოლოგიურ „წრედებში“ ეს, უმეტეს შემთხვევაში, რომელიმე გენის ექსპრესიაში (ანუ მასში კოდირებული ინფორმაციის „წაკითხვაში“) გამოიხატება. ასეთი გადაცემის მექანიზმი შეიძლება განისაზღვრებოდეს მრავალი გენითა და ცილით თითოეულ დონეზე, ან ერთადერთი მოლეკულით, რომელსაც სამივე დონეზე აკისრია სპეციფიური ფუნქცია.

ნახ. 1: სინთეტური ბიოლოგიის შემადგენელი ნაწილები და ამ დარგის მიღწევების გავლენა ჩვენი ცხოვრების მრავალ ასპექტზე.
სინთეტური ბიოლოგიის მიღწევათა უმეტესობა ბაქტერიულ სისტემებთანაა დაკავშირებული, რაც ადვილად აიხსნება როგორც ბაქტერიების შედარებითი სიმარტივით, ასევე მათი უჯრედების შემადგენლობის თითქმის სრულყოფილი შესწავლით. ამ სისტემებში მიღწეული წარმატებების მაგალითებია სინათლის მგრძნობიარე სიგნალის გადამცემი სისტემები, და რხევით პრინციპზე აგებული „წრედები“, რომლებიც გენების ექპრესიას მართავენ. ამის მიუხედავად, ბოლო წლებში მნიშვნელოვნად განვითარდა სისტემები, რომლებიც გაცილებით უფრო რთული აგებულების უჯრედებში (საფუარის, ცხოველების, მცენარეების) მოქმედებს.
უჯრედის სასიგნალო „წრედების“ ოპტიმიზაცია. უჯრედის სასიგნალო სისტემებში მოქმედებს ე.წ. კასკადური პრინციპი, როდესაც გარემოდაც შემოსული სიგნალის გადაცემა ხდება ჯერ უჯრედის გარსის გავლით, შემდეგ კი უჯრედის შიგნით. ეს გადაცემა სრულდება მრავალი ცილის ურთიერთქმედების შედეგად. ბუნებრივ პირობებში, ყოველი ცილა მისი „წინამორბედი“ ცილის გავლენით ამოქმედდება. საფუარის უჯრედებში დამუშავებულ სინთეტურ სისტემაში კი მოახდინეს მნიშვნელოვანი მოდიფიკაცია, რომლის შედეგადაც ამ ჯაჭვში მოქმედი ბოლო ცილა მოქმედებდა პირველ (ჯაჭვის დასაწყისში მოთავსებულ) ცილაზე და ააქტიურებდა მას. ასეთი ცვლილების შედეგად, კასკადის ამოქმედებისათვის სიგნალის მხოლოდ ერთჯერადი მიწოდება იყო საკმარისი: ამის შემდეგ სისტემა „თვითკმარი“ ხდებოდა. ამ სისტემის გამოყენებით, შესაძლებელი გახდა არაერთი სასარგებლო პროდუქტის (ცილის) ოპტიმალური გამომუშავება როგორც საფუარის, ასევე მცენარეების უჯრედებში.
სენსორის ფუნქციის შეცვლა მკვლევართა სურვილისამებრ შესაძლებელია მოხდეს ისეთი რეცეპტორების (სიგნალის მიმღები და გადამცემი მოლეკულების) შექმნით, რომლებიც მხოლოდ კონკრეტული თვისებების სიგნალებს ცნობენ. გენეტიკური ინჟინერიის მეთოდების გამოყენებით, შეიქმნა RASSL-რეცეპტორები (receptors activated solely by synthetic ligands - რეცეპტორები, რომელთა აქტივაცია ხდება მხოლოდ სინთეტური ლიგანდების (ანუ სიგნალების) მიერ).
გენების ექსპრესიის სინთეტური რეგულაცია. გენების ექსპრესია (ანუ მათში კოდირებული ინფორმაციის „წაკითხვა“, ასევე - „ტრანსკრიფცია“) უჯრედებისათვის სიგნალის მიღებისა და გადაცემის ბოლო (და საკვანძო) ეტაპია. მრავალ შემთხვევაში, ამ ეტაპის განხორციელება ხდება ე.წ. გენეტიკური გადამრთველების მეშვეობით. ეს გადამრთველები წარმოადგენენ ცილებს, რომლებსაც აქვთ დნ მჟავასთან ბმის უნარი. ამ უნარის გამო, ასეთ ცილებს შეუძლიათ, „უშუალო კონტაქტში“ შევიდნენ გენებთან (შეგახსენებთ, რომ გენები დნ მჟავის ფრაგმენტებია) და, მიღებული სიგნალის პასუხად, ჩართონ ან გამორთონ კონკრეტული გენის ექსპრესია. ამ პრინციპის საფუძველზე, შექმნილია „ტრანსკრიფციის ხელოვნური ფაქტორები“ (ATF, artificial transcription factors), რომლებიც გამოიყენება ცალკეული გენების ექსპრესიის მართვისათვის. ზოგიერთ შემთხვევაში, ATF იმგვარადაა „დაპროექტებული“, რომ მათი ბმა დნ მჟავასთან დამოკიდებულია მცირე ზომის მოლეკულების (სიგნალის) არსებობასა თუ არარსებობაზე.
სინთეტური „წრედების“ ჩართვა ბუნებრივ სისტემებში. ასეთი მიდგომის ერთ-ერთი საუკეთესო მაგალითია თერაპიული ნაერთის (მედიკამენტის) სინთეზის იმგვარი მართვა, როდესაც უჯრედები მედიკამენტის წარმოებას მხოლოდ ავადმყოფობის შემთხვევაში იწყებენ. ამ შემთხვევაში გამოიყენება გენის რეგულატორული ფრაგმენტები, ანუ ის ფრაგმენტები, რომელთა მეშვეობითაც ხდება გენების ექსპრესიის მართვა (მაგ., ამ ფრაგმენტებს უშუალოდ ებმიან ტრანსკრიფციის ფაქტორები). ეს რეგულატორული ფრაგმენტები სინთეტურ სისტემებში სენსორის ფუნქციას ასრულებენ: ზოგიერთი მათგანისათვის სიგნალს ჰორმონები წარმოადგენენ, სხვებისათვის კი - ჟანგბადის ნაკლებობა, გარკვეული მოლეკულების კონცენტრაციის შეცვლა და ა.შ. ასეთი ხელოვნური კომპონენტების შეყვანა ბუნებრივ „წრედებში“ შესაძლოა ეკოლოგიური სისტემების მდგომარეობაზე დაკვირვებისთვისაც: თუ რეგულატორული ფრაგმენტი, გარკვეული მავნე ქიმიური ნაერთების დაგროვების პასუხად იწყებს მოქმედებას,მაშინ მის მიერ მართვადი გენის ექსპრესია ასეთი ნაერთების ნიადაგსა თუ ჰაერში დაგროვების უტყუარი საბუთი იქნება.
სინთეტური ბიოლოგიის წვლილი მოლეკულური ბიოლოგიისა და მედიცინის განვითარებაში. უპირველეს ყოვლისა, სინთეტური მექანიზმების შექმნასა და მათ გამოცდას ცოცხალ ორგანიზმებში ამოწმებს უკვე არსებული ცოდნის სისრულესა და მისი გამოყენების ეფექტურობას. ხელოვნური ნაწილების ჩართვა ბუნებრივ სისტემებში სულ ცოტა სამ მიზანს ასრულებს: 1) ამ სისტემების შესაძლო ახალი თვისებების აღმოჩენა; 2) კვლევისათვის სასარგებლო ხელსაწყოების შექმნა; 3) არსებული მოდელების შემოწმება.
სინთეტური წრედები იქმნება არა მარტო ბიოლოგიური კვლევის, არამედ საინჟინრო მიზნების მისაღწევადაც, ამიტომ ამ წრედების კომპონენტები ხშირად უნიკალური (ბუნებაში არარსებული) თანმიმდევრობით ერთდებიან. იმ შემთხვევებში, როცა ამ სისტემის ამუშავება წარუმატებელია საინჟინრო თვალსაზრისით, მაინც შესაძლოა მისი კომპონენტების ახალი ბიოლოგიური თვისებების წარმოჩენა.

ნახ. 2: უჯრედი, როგორც საინჟინრო კვლევის ობიექტი.
არსებობს მედიცინის განვითარებაში შეტანილი წვლილის რამდენიმე კონკრეტული მაგალითიც: სინთეტური ბიოლოგიის მიღწევების შედეგად, შესაძლებელი გახდა მალარიის საწინააღმდეგო მედიკამენტის წარმოება ბაქტერიებისა და საფუარის უჯრედებში, ისევე როგორც ახალი წამლების გამოცდის ტექნოლოგიის დამუშავება ადამიანის უჯრედებში.
დასკვნა.
ახალი სინთეტური სისტემების (წრედების) განვითარებისათვის დიდი მნიშვნელობა აქვს მათთვის გამოსადეგი კომპონენტების იდენტიფიკაციასა და დახასიათებას. ასეთი კომპონენტები შეიძლება გამოიყოს ბუნებრივი სისტემებიდან, ან ხელოვნურად შეიქმნას გენური ინჟინერიის მეთოდების გამოყენებით. ახალი კომპონენტების საგულდაგულო შემოწმება და ასეთი შემოწმების სტანდარტების დადგენა არანაკლებ მნიშვნელოვანი ეტაპია. თუ გავითვალისწინებთ, რომ სინთეტური ბიოლოგია სულ უფრო მეტად იყენებს რთული აგებულების (მცენარეებისა და ცხოველების) უჯრედებს, ამ უჯრედებთან მუშაობის პროცედურების გაუმჯობესებაც მნიშვნელოვანი შენაძენი იქნება.
სინთეტური ბიოლოგიის მიღწევებმა უკვე გვიჩვენა, რომ ხელოვნურ სისტემებს ძალუძთ რთულ ბიოლოგიურ გარემოში მოქმედება, და ამ მოქმედების ეფექტურობა მნიშვნელოვნად არ ირღვევა გარეშე (ზოგ შემთხვევაში უცნობი) ფქტორების გავლენით. მეტიც, მცენარეთა და ცხოველთა უჯრედებში არსებული რთული სისტემები (სიგნალის გადაცემა, ცილის სინთეზი და მისი უჯრედისშიდა განლაგება) შესაძლებელია გამოვიყენოთ სხვადასხვანაირი სინთეტური წრედების შესაქმნელად. უჯრედების ჯერ კიდევ არასრულად შესწავლილი კომპონენტების არსებობა ფართო ასპარეზს უქმნის სინთეტური ბიოლოგიის განვითარებას.
გელა თევზაძე.
24 საათი 27.03.11
|
|
7 სისტემების ბიოლოგია: ექსპერიმენტებში დაგროვილი მასალის ანალიზი და სინთეზი |
▲ზევით დაბრუნება |
მოლეკულური ბიოლოგიის (და ზოგადად, ექსპერიმენტული ბიოლოგიის) განვითარების შედეგად დაგროვდა დიდი მასალა უჯრედებისა თუ ორგანიზმების ცალკეული ნაწილების ფუნქციებსა და მოქმედების მექანიზმებზე. ამ მასალის საფუძველზე, შესაძლებელი გახდა ბიოლოგიის ახალი დარგის განვითარება - დარგისა, რომელიც შესაძლებელს გახდიდა მთელი უჯრედისა თუ ორგანიზმის სტრუქტურისა და ფუნქციის შესწავლას. ამ დარგს ეწოდა „სისტემების ბიოლოგია“ (Systems Biology). სახელწოდებაში ასახულია ის გარემოება, რომ ნებისმიერი ორგანიზმი (როგორც ერთუჯრედიანი, ასევე მრავალუჯრედიანი) განიხილება როგორც ცალკეული ნაწილებისაგან შემდგარი სისტემა. ამ სისტემის შესწავლა უკვე შესაძლებელია - პირველ რიგში იმის გამო, რომ მისი კომპონენტების შესახებ უკვე დიდძალი მასალაა დაგროვილი, და შესაძლებელია მათი ურთიერთქმედებისა და ფუნქიათა ოპტიმალური ერთობლიობის გაანგარიშება.
სისტემების ბიოლოგიის განვითარება მოითხოვს ზოგადი ბიოლოგიის შესასწავლი საგნების ახლებურად გააზრებას. ცალკეული გენებისა თუ ცილების შესწავლა კვლავინდებურად მნიშვნელოვანია, მაგრამ მთელი სისტემის სტრუქტურისა და დინამიკის შესწავლა უმთავრეს ამოცანად გვევლინება. სისტემა არ წარმოადგენს გენებისა და ცილების უბრალო ერთობლიობას, ამიტომ მხოლოდ მათი ურთიერთქმედების დიაგრამის შედგენით სისტემის თვისებების სრულფასოვანი შესწავლა ვერ მიიღწევა. უეჭველია, რომ ასეთი დიაგრამის შედგენა პროცესის პირველი მნიშვნელოვანი საფეხურია, მაგრამ ის შეიძლება მეტაფორულად შევადაროთ ქუჩის რუქას, მაშინ როდესაც სისტემის სრულყოფილი შესწავლა - ისევ მეტაფორულად - ტრანსპორტის მოძრაობის დინამიკას, ტენდენციებსა და მათ წარმომშობ ფაქტორებს, და ამ ფაქტორების კონტროლის მექანიზმების გარკვევასაც ითვალისწინებს.
ორგანიზმის ყველა გენისა და ცილის იდენტიფიკაცია თვითმფრინავის ყველა დეტალის შემცველი სიის შედგენის მსგავსია. რასაკვირველია, ასეთი სია ნაწილების სრულ აღწერას იძლევა, მაგრამ მხოლოდ მის საფუძველზე ვერ გავიგებთ თვითმფრინავის რთულ მოწყობილობას და მოქმედების მექანიზმს. ამისათვის საჭიროა კიდევ იმის ცოდნაც, თუ როგორ უკავშირდება ეს ნაწილები ერთმანეთს თვითმფრინავის კონსტრუქციაში. სწორედ ამ ცოდნის სადარია ბიოლოგიური სისტემების შემადგენელი ნაწილების მოქმედების დინამიკის შესწავლა.
ნებისმიერი ბიოლოგიური ორგანიზმის შესწავლა სისტემურ დონეზე ოთხ შემადგენელ ნაწილად შეიძლება დაიყოს:
სისტემის სტრუქტურა: გენების ურთიერთქმედება და ბიოქიმიური რექაციების ერთობლიობა; მექანიზმები, რომლებითაც ხდება უჯრედშორისი და მრავალუჯრედიანი სტრუქტურების თვისებების ცვლილება.
სისტემის დინამიკა: როგორ მოქმედებს სისტემა დროის განმავლობაში? სისტემა განიხილება, როგორც მრავალგანზომილებიანი სტრუქტურა, რომელშიც თითოეული განზომილება წარმოდგენილია ერთ-ერთი შემადგენელი ნაწილის (ბიოქიმიური ფაქტორის) გარკვეული კონცენტრაციით.
კონტროლის მეთოდი: არსებობს მექანიზმები, რომლებიც უჯრედის ზრდასა და განვითარებას მართავენ. შესაძლებელია ამ მექანიზმების რეგულირება მათი შეცდომების თავიდან აცილების მიზნით. ამ მეთოდით ასევე შეიძლება მრავალი ავადმყოფობის განკურნების სტრატეგიის შემუშავებაც, მედიკამენტების „სამიზნის“ იდენტიფიკაციის გზით.
დაპროექტების („დიზაინის“) მეთოდი: სრულყოფილი სისტემების დაპროექტება შემუშავებული პრინციპებისა და მეთოდების საფუძველზე. პირდაპირი ექსპერიმენტული მიდგომისაგან განსხვავებით, ასეთი სტრატეგია ნაკლებ შეცდომას წარმოშობს.
ასეთ კვლევაში, გენების რეგულაციისა და ბიოქიმიური სისტემების „შინაგანი ლოგიკის“ გამოაშკარავება ყველაზე ძნელად გადასაჭრელი ამოცანაა. ტრადიციული მიდგომა ითვალისწინებს მრავალი ექსპერიმენტის შედეგებისა და ლიტერატურაში არსებული ცნობების დამუშავებას. არსებობს (ან შემუშავების სტადიაშია) ცნობათა ბაზები, რომლებიც შეიცავენ ცნობებს გენების რეგულაციისა და ბიოქიმიური სისტემების შესახებ, მაგრამ ისინი სრულყოფილებისაგან შორს არიან: მრავალი სისტემა ამ ბაზების შემადგენლობის მიღმა რჩება, ან მათი მოქმედების მექანიზმი ჯერ-ჯერობით ამოუცნობია.
სისტემური ანალიზის ერთ-ერთი სტრატეგიაა ორგანიზმის ყველა გენის ექსპრესიის (ანუ მათში კოდირებული ინფორმაციის „ამოკითხვის“) დეტალური შესწავლა: რა ფაქტორები მართავენ ექსპრესიას, დროის რა მონაკვეთში ხდება იგი და როგორ შეიძლება გენების დაჯგუფება ამ კრიტერიუმების საფუძველზე? გენების დაყოფა ასეთ ჯგუფებად („კლასტერებად“) უდაოდ სასარგებლოა გენების ექსპრესიისა და ბიოლოგიური პროცესების შორის არსებული კორელაციის აღმოსაჩენად, მაგრამ ის ყოველთვის ვერ წარმოაჩენს მიზეზ-შედეგობრივ კავშირებს რეგულატორულ სისტემებში. მაგალითად, რომელიმე გენის ინფორმაციის „ამოკითხვა“ შეიძლება ხდებოდეს რაიმე საკვები ნივთიერების ნაკლებობის საპასუხოდ, და ეს ნაკლებობა შეიძლება ასევე გახდეს განსხვავებული ბიოლოგიური პროცესის წინაპირობაც (მაგ., უჯრედის გაყოფის შეჩერების ან გაყოფის მექანიზმის შეცვლისა). ამ შემთხვევაში შეცდომა იქნებოდა, რომ ხსენებული გენის ფუნქცია ამ რომელიმე პროცესთან უშუალოდ დაგვეკავშირებინა: ამ შემთხვევაში კავშირი მხოლოდ იმაში მდგომარეობს, რომ ორივე ეფექტი (გენის ექსპრესია და უჯრედის გაყოფის მსვლელობის ცვლილება) ერთი და იგივე მიზეზით (საკვები ნივთიერების ნაკლებობით) არის გამოწვეული.
როგორც კი სისტემის სტრუქტურას გავიგებთ, სისტემის დინამიკის შესწავლაც ხდება შესაძლებელი. უმეტეს შემთხვევაში, სისტემის სტრუქტურისა და დინამიკის შესწავლა ერთსა და იმავე დროს ხდება - ეს ორი პროცესი „ურთიერთგადამკვეთია“: დინამიკის ანალიზი ზოგჯერ საშუალებას იძლევა, აქამდე უცნობი ურთიერქმედებების არსებობა ვივარაუდოთ. დინამიკის შესწავლისათვის საჭიროა პროცესის მოდელირება, და ამ დროს განსაკუთრებით მნიშვნელოვანია მოდელის მიზნების, მისი მასშტაბისა და „აბსტრაგირების დონის“ (abstraction level) განსაზღვრა. ანალიზის მეთოდის არჩევანი დამოკიდებულია იმ - ბიოლოგიური ექსპერიმენტების შედეგად მიღებული - ინფორმაციის რაოდენობაზე, რომელიც მოდელში იქნება გამოყენებული.
როცა სისტემის სტრუქტურაცა და ფუნქციით განსაზღვრული თვისებებიც მეტ-ნაკლებად გარკვეულია, დგება კლასიფიცირებისა და სხვა სისტემებთან შედარებითი ანალიზის ჯერი. ასეთი ანალიზი გვაჩვენებს, რამდენად მრავალფეროვანია ამ სისტემის სხვადასხვა ვერსიები და რამდენად ცვალებადი ან კონსერვატული იყო ეს სისტემა ევოლუციის განმავლობაში. არსებობს მოსაზრება, რომ ამ საკითხის ინტენსიური ანალიზი გამოავლენს ბიოქიმიური სისტემების ევოლუციურ ოჯახს, და საფუძველს ჩაუყრის ფუნქციონალური რეგულატორული მექანიზმების „პერიოდული ტაბულის“ შექმნას.
ბიოლოგიური სისტემების ერთ-ერთ მნიშვნელოვან დამახასიათებელ ნიშანს მათი გამძლეობა (robustness) წარმოადგენს. გამძლეობის ბიოლოგიური საფუძვლების ანალიზი აუცილებელია კვლევისათვის სისტემების ბიოლოგიის ფარგლებში. გამძლეობის ფენომენოლოგიური თვისებები სამ ჯგუფად შეიძლება დაიყოს: i) ადაპტაცია, რომელიც გარემო პირობების ცვლილებების პასუხად შეგუების უნარის აღმნიშვნელია; ii) დამოუკიდებლობა გარკვეული პარამეტრებისაგან, რაც აღნიშნავს სისტემის მიერ მცირე მგრძნობიარობას ზოგიერთი ცვლილების მიმართ; iii) „მოხდენილი“ დეგრადაცია (graceful degradation), რაც გამოიხატება სისტემის ფუნქციების თანდათანობით დაქვეითებაში (უეცარი გაჩერების საპირისპიროდ). საინჟინრო სისტემებში, გამძლეობა მიღწეულია რამდენიმე ხერხით: i) ორივე მიმართულებით მოქმედი კონტროლის სისტემებით; ii) სათადარიგო კომპონენტების შექმნით, როცა რამდენიმე დეტალს ერთი და იგივე ფუნქცია აკისრია; iii) სტრუქტურული სტაბილურობა, როდესაც სისტემის შინაგანი მექანიზმები უზრუნველყოფენ მდგრადობას; iv) მოდულარული სტრუქტურა, როცა სისტემა ცალკეული მოდულებისაგან შედგება, და ერთი მოდულის ფუნქციის დარღვევა მთელი სისტემის დაშლას არ იწვევს. ცხადია, რომ ასეთივე პრინციპებით აგებული „სქემები“ უხვად არსებობს ბიოლოგიურ სისტემებშიც: ბაქტერიული ქემოტაქსისი (მოძრაობა გარკვეული ნაერთების მზარდი კონცენტრაციის მიმართულებით) გამძლეობის ზემოთნახსენებ სამივე პრინციპს აკმაყოფილებს. სათადარიგო დეტალების მაგალითების მონახვა შეიძლება როგორც ცალკეული გენების, ასევე მათი ერთობლიობის შემთხვევაშიც (შესაბამისად, უჯრედის ზრდისა და გაყოფის ციკლში მონაწილე გენები, და ბაქტერიების მეტაბოლიზმში მოქმედ რეაქციათა ჯაჭვის ალტერნატიული გზები). სტრუქტურული სტაბილურობა შეინიშნება ხილის ბუზის (დროზოფილის) სხეულის სეგმენტების წარმოქმნისას, ხოლო მოდულარული სტრუქტურა მიღწეულია როგორც ინდივიდუალური უჯრედების დონეზე, ასევე სასიგნალო სისტემებში, რომლებიც განცალკევებულნი არიან, მაგრამ ურთიერთშეთანხმებულად მოქმედებენ.
1. სისტემების ბიოლოგიის „წრედი“ - ჰიპოთეზებზე დაფუძნებული კვლევა სისტემების ბიოლოგიაში.
სისტემური ანალიზისათვის ასევე აუცილებელია რაოდენობრივი მონაცემების მნიშვნელოვანი რაოდენობაც. იმ კვლევები, რომელთა მიზანია უჯრედების მოქმედი მოდელის შექმნა, სასურველია მოდელირების წინასწარი ანალიზი (მაგ., მოდელის სიზუსტისა და თავდაპირველ მიზანთან შესაბამისობისა), რათა საბოლოო მოდელის მიღებამდე უსარგებლო გამოთვლების დაგროვება თავიდან იყოს აცილებული.
გამოთვლების სიზუსტე და მართებულობა სამი ასპექტის გათვალისწონებას მოიცავს: i) ანალიზის სრულფასოვნება, რითაც აღინიშნება იმ მოლეკულების (ნუკლეინის მჟავებისა და ცილების) რიცხვი, რომელთა რაოდენობის გაზომვა ერთდროულადაა შესაძლებელი; ii) დროის ფაქტორი - იმის გამოსარკვევად, თუ რა დრო სჭირდება მოცემული ანალიზის შესრულებას; iii) ანალიზის „ყოვლისმომცველობა“, რაც სისტემის მიერ რამდენიმე სხვადასხვა ფაქტორის სხვადასხვაგვარი ანალიზის ერთდროული ჩატარების შესაძლებლობას არკვევს. ექსპერიმენტის დაგეგმვა მოდელის საფუძველზე საშუალებას გვაძლევს შევიტყოთ, რომელ კომპონენტში/სტადიაზეა საჭირო მეტი ან ნაკლები სიზუსტე, რაც სისტემის ოპტიმიზაციის საშუალებას იძლევა.
2. ბიოლოგიური სისტემების სირთულია ამსახველი სქემა - სიგნალის გადაცემა უჯრედის გარედან უჯრედის ბირთვში.
ბიოლოგიური რეგულაციის სრული (და სრულყოფილი) სისტემური ანალიზი ჯერ-ჯერობით თანამედროვე ექსპერიმენტული მეცნიერების შესაძლებლობების მიღმა რჩება. ექსპერიმენტული მეთოდებისა და ხელსაწყოების შემდგომი სრულყოფა უმნიშვნელოვანესი ფაქტორია სისტემების ბიოლოგიის განვითარებასათვის. ასეთი მეთოდები არა მარტო გააუმჯობესებენ ინფორმაციის შეგროვების სისწრაფეს, არამედ პროცესის ავტომატიზაციასაც შეუწყობენ ხელს. არანაკლებ მნიშვნელოვანია კომპიუტერული პროგრამები, რომელთა მეშვეობით მოხდება ბიოლოგიური პროცესების მოდელირება და მიღებული შედეგების მასობრივი ანალიზი.
სისტემების ბიოლოგია მეტად მნიშვნელოვანია ფარმაკოლოგიური წარმოებისა და სამედიცინო კვლევისათვის: დღესდღეობით, ყველაზე ადვილად წარმოსადგენია უჯრედის რეგულაციის დეტალური მოდელის შექმნა, რომელიც ფოკუსირებული იქნება სასიგნალო მოლეკულებსა და მათგან შემდგარი კასკადური რეაქციების მექანიზმებზე. ასეთი მოდელის დახმარებით, ახალი მედიკამენტების სტრუქტურის განსაზღვრა მთელი სისტემის აგებულების გათვალისწინებით მოხდება. იგივე მოდელების საშუალებით შესაძლებელი იქნება იმის განსაზღვრაც, თუ რომელი მექანიზმების ჩართვა გამოიწვევს მედიკამენტის მოქმედების განეიტრალების არასასურველ შედეგს, და როგორი დამატებითი (ე.წ. „გვერდითი“) ეფექტები შეიძლება იყოს მოსალოდნელი. არაა გამორიცხული, რომ მომავალში დაავადებული უჯრედების განკურნების მიზნით მრავალი მედიკამენტის ერთობლივი გამოყენებაც იქნება შესაძლებელი, და მოდელირების საფუძველზე გაანგარიშებული სტრატეგიის გამო, მათი მოქმედება მაქსიმალურად იქნება კოორდინირებული.
დასკვნა:
თუმცა სისტემების ბიოლოგია თავისი არსებობის საწყის სტადიებშია, მას მეტად მნიშვნელოვანი პერსპექტივები გააჩნია როგორც ფუნდამენტური მეცნიერების პრობლემების, ასევე გამოყენებითი საკითხების გადაჭრის თვალსაზრისით. ექსპერიმენტული ბიოლოგია თანდათან გადადის მოლეკულური დონიდან სისტემების დონეზე, და ამ გადასვლას შეუძლია უმნიშვნელოვანესი ძვრები მოახდინოს რთული ბიოლოგიური მექანიზმების შესწავლაში, ისევე როგორც ამ შესწავლის შედეგად დაგროვილი ცოდნის პრაქტიკულ გამოყენებაში.
გელა თევზაძე.
24 საათი 17.04.11
|
|
8 მანათობელი ცილები: კვლევითი და დეკორატიული მნიშვნელობა |
▲ზევით დაბრუნება |
მოლეკულური ბიოლოგიის ერთ-ერთი მნიშვნელოვანი საკითხია გენების მიერ კოდირებული ცილების განლაგების (ლოკალიზაციის) შესწავლა. ასეთი კვლევისათვის მეტად სასარგებლოა ცილების დანიშვნა (მარკირება) ისეთი მოლეკულით, რომლის უჯრედსშიდა განლაგების განსაზღვრა შედარებით იოლია. ასეთი მოლეკულის კარგი მაგალითია „მანათობელი ცილები“, რომლებიც ხშირად და წარმატებულად გამოიყენება მოლეკულურ ბიოლოგიაში. მანათობელი („ფლუორესცირებადი“) ცილების თვისებები და გამოყენებითი მნიშვნელობა ამ სტატიის თემას წარმოადგენს.
მწვანე მანათობელი ცილა (Green Fluorescent Protein, GFP): 1962 წელს მედუზის ერთ-ერთი სახეობის (Aequorea victoria) ფლუორესცენციის (ნათების) მექანიზმის შესწავლისას აღმოჩნდა, რომ ნათებას იწვევდა ცილა, რომელიც მზის სინათლეზე მომწვანო ფერისა ჩანდა, ულტრაიისფერ სხივებში კი მკვეთრი მწვანე შუქით ანათებდა. ეს აღმოჩენა ეკუთვნის ოსამუ შიმომურას პრინსტონის უნივერსიტეტიდან (Osamu Shimomura, Princeton Univeristy), მისი დამსახურება 2008 წელს ნობელის პრემიით აღინიშნა. 1979 წელს, ისევ შიმომურას ექსპერიმენტების შედეგად, მოხდა ქრომოფორის (ცილის მოლეკულის მანათობელი ნაწილის) იდენტიფიკაცია (ნახ. 1): გაირკვა, რომ ქრომოფორი წარმოიქმნებოდა GFP-ს სამი მომიჯნავე ამინომჟავისაგან (ამინომჟავები ცილის შემადგენელი ერთეულებია), მათ შორის დამატებითი ქიმიური ბმების წარმოქმნის შედეგად.
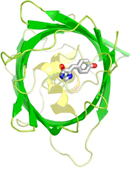
ნახ. 1: მწვანე მანათობელი ცილის სტრუქტურა. ცენტრში მოთავსებულია ქრომოფორი (ორი შეკავშირებული ექვსკუთხედი).
1992 წელს მოხდა GFP-ს გენის კლონირება. აღმოჩნდა, რომ GFP დიდი ზომისა არაა - ის მხოლოდ 238 ამინომჟავისაგან შედგება და, რაც მთავარია, მას ნებისმიერ ორგანიზმში შეუძლია ქრომოფორის წარმოქმნა, რის შედეგადაც ვიღებთ მანათობელი ეფექტი. ორიოდე წლის შემდეგ GFP-ს გენის ბაქტერიებში შეყვანის შედეგად მიიღეს „მწვანე მანათობელი ბაქტერიები“. 1995 წლის ნოემბრისათვის GFP-ს შეყვანა 30-ზე მეტ სხვადასხვა ორგანიზმში განხორციელდა, მომდევნო ათწლეულში კი წელიწადში საშუალოდ 500-ზე მეტ სტატიაში იყო გამოყენებული GFP-მეთოდოლოგია.
სხვა ორგანიზმების მანათობელი ცილები: Aequorea-ს მწვანე მანათობელი ცილის მზარდმა პოპულარობამ წარმოშვა კითხვა, არსებობს თუ არა მსგავსი მანათობელი ცილები სხვა ორგანიზმებში. ასევე, ბიოლოგიური კვლევაში GFP-ს გამოყენების შედეგად, წარმოიქმნა სხვა ფერის მანათობელი ცილების საჭიროებაც (მაგ., რომელიმე ორგანიზმის ორი სხვადასხვა ცილის შესასწავლად, თუ ერთი მათგანი GFP-თ იქნებიდა „დანიშნული“, მეორე კი - განსხვავებული ფერის ცილით, იხ. ქვემოთ). 1999 წელს მიხეილ მატცისა და სერგი ლუკიანოვის (რუსეთის მეცნიერებათა აკადემია; Михаил Матц, Сергей Лукьянов, РАН) მიერ გამოქვეყნდა ცნობა წითელი მანათობელი ცილის შესახებ, რომელიც მარჯნის პოლიპების ერთ-ერთი სახეობის (ზღვის ანემონების, ნახ. 2) საცეცებიდან მიიღეს. ამ წითელ ცილას ეწოდა DsRed, რადგან იგი მიიღეს ანემონიდან, რომლის ლათინური სახელწოდებაა Discosoma sp. DsRed-ის შემდგომმა შესწავლამ და სტრუქტურულმა ანალიზმა აჩვენა, რომ DsRed-სა და GFP-ს ბევრი საერთო თვისება აქვს და ქრომოფორების წარმოქმნის მექანიზმებიც მსგავსია. ამ კვლევების საფუძველზე გაირკვა, რომ მარჯნის კოლონიებში არსებული მრავალი ფერი განისაზღვრება სწორედ მანათობელი (ფლუორესცირებადი) ცილების არსებობით მარჯნის პოლიპების უჯრედებში. ეს ცილები GFP-ს მსგავსი მოლეკულების ჯგუფს (უფრო ზუსტად, ოჯახს) მიეკუთვნებიან. ამ ჯგუფის ცილებს მსგავსი სტრუქტურა ახასიათებთ, მაგრამ აღგზნებადობისა (გაღიზიანების საპასუხოდ ნათების წარმოქმნის სისწრაფით) და ფლუორესცენციის სიძლიერის თვალსაზრისით განსხვავდებიან.

ნახ. 2: ზღვის ანემონები, რომლებიც წითელ მანათობელ ცილას შეიცავენ.
მანათობელი ცილებისა და მათი მსგავსი მოლეკულების ანალიზი მედუზებში, კიბოსნაირებში და ხერხემლიანებში მიგვანიშნებს, რომ ამ ცილებს საერთო წინაპარი ჰყავდათ. სავარაუდოა, რომ ევოლუციის განმავლობაში პირველი მანათობელი ცილა 500-1000 მილიონი წლის წინ წარმოიქმნა, და რომ მას მწვანე ნათება შეეძლო. წითელი მანათობელი ცილები უფრო გვიანდელი წარმოშობისაა, და იგი ერთმანეთისაგან დამოუკიდებლად წარმოიქმნა რამდენიმე განსხვავებული სახეობის ორგანიზმში. ხერხემლიანებში (მათ შორის ადამიანში) მანათობელი ცილების მსგავსი სტრუქტურის მოლეკულები არსებობს, მაგრამ მათი ნათების უნარი დაკარგულია მუტაციების შედეგად იმ ამინომჟავებში, რომლებიც მნიშვნელოვანია ქრომოფორის წარმოქმნისათვის.
მანათობელი ცილების გაუმჯობესებული ვერსიები: GFP-ს აღმოჩენით გამოწვეული ენთუზიაზმის მიუხედავად, მალე გასაგები გახდა, რომ ეს ცილა იდეალურად არ შეესაბამებოდა ბიოლოგიური ექსპერიმენტების მოთხოვნილებებს. მაგალითად, GFP-ს ქრომოფორის წარმოქმნისათვის ოპტიმალურია დაბალი ტემპერატურა, რაც ადვილად აიხსნება იმ გარემოებით, რომ მედუზის ცხოვრებია გარემო ოკეანეა და ცილის ევოლუცია სწორედ ცივ გარემოსთან შეთავსების მიმართულებით წარიმართა. დაბალ ტემპერატურაზევე უფრო ადვილად ხდებოდა ცილის მესამეული სტრუქტურის წარმოქმნაც (ე.წ. „დაგორგვლა“), რაც ასევე აუცილებელია GFP-ს ფუნქციისათვის. ბიოლოგიური ექსპერიმენტების ჩატარებისას კი უფრო გამოსადეგი იქნებოდა ცილა, რომლისთვისაც როგორც „დაგორგვლის“, ასევე ქრომოფორის წარმოქმნის ოპტიმალური ტემპერატურა 37°C-ს ფარგლებში იქნებოდა. 1996 წელს GFP-ს სწორედ ასეთი ვარიანტი მიიღეს, და ეს შედეგი შესაძლოა განვიხილოთ, როგორც გენური ინჟინერიის გეგმაზომიერი გამოყენების ერთ-ერთი უბრწყინვალესი მაგალითი: „სითბოს მოყვარული“ ვარიანტის მიღება მოხდა GFP-ს გენში იმ ფრაგმენტების იდენტიფიკაციით, რომლებიც შესაბამის ცილაში „სიცივისადმი მიდრეკილებას“ განსაზღვრავდნენ, და ამ ფრაგმენტების შეცვლა მუტაციების შეყვანის გზით. ასევე, GFP-გენის მუტირებით შესაძლებელი გახდა ცილის კიდევ ერთი თვისების შეცვლა: უჯრედებში GFP-ს დიდი კონცენტრაციით დაგროვების შედეგად მისი დიმერიზაცია (ორმოლეკულიანი სტრუქტურის წარმოქმნა) ხდება, რაც ნათების შესუსტებას იწვევს და ექსპერიმენტული თვალსაზრისით არასასურველია. მუტანტური GFP კი მონომერის (ერთმოლეკულიანი სტრუქტურის) სახით არსებობს თავისი კონცენტრაციის მიუხედავად.
მკვლევართა აზრით, GFP-ს შეზღუდული შესაძლებლობები ასევე გამოიხატებოდა მის „ერთფეროვნებაში“: არ არსებობდა სხვადასხვა ფერის მანათობელი ცილები (DsRed წითელი ცილის გამონაკლისით). გენური ინჟინერიის მეთოდების გამოყენებით ამ სირთულის დაძლევაც მოხერხდა: 1990-იან წლებში შეიქმნა ლურჯი (Blue Fluorescent Protein, BFP), ლურჯ-მწვანე (Cyan, CFP) და ყვითელი (Yellow, YFP) ვარიანტები. საინტერესოა, რომ ლურჯი და მწვანე ვერსიები შეიქმნა ქრომოფორში (ამ ფრაგმენტის სამი ამინომჟავიდან ერთ-ერთის, იხ. ზემოთ) მუტაციების შეყვანით, ხოლო ყვითელ ცილაში არა ქრომოფორი, არამედ მის ფარგლებს გარეთ არსებული ამინომჟავაა შეცვლილი.
არანაკლებ მნიშვნელოვანი ცვლილებები განიცადა წითელმა მანათობელმა ცილამ. ამ შემთხვევაში, ბიოლოგიური ექსპერიმენტების თვალსაზრისით მთავარ პრობლემებს წარმოადგენდა DsRed-ის ტეტრამერიზაცია (ოთხმოლეკულიანი სტრუქტურა), რაც ასევე ამცირებდა ნათებას, ისევე როგორც ქრომოფორის წარმოქმნის მცირე ეფექტურობა და დაბალი სიჩქარე. მონომერული DsRed მიღებული იქნა გენური ინჟინერიისა და ე.წ. „მართული ევოლუციის“ მეთოდების გამოყენებით. მიღებული მონომერული ცილიდან რამდენიმე ვარიანტია მიღებული, რომელთა ნათების სპექტრი ინფრაწითელიდან ყვითელ ფერამდე ვრცელდება.
მანათობელი ცილები ბიოლოგიურ კვლევაში: მანათობელი ცილები ყველაზე ხშირად გამოიყენება ცილების უჯრედსშიდა განლაგების გამოსარკვევად. ამ მიზნით, მანათობელი ცილის გენს „გადააბამენ“ იმ გენს, რომელშიც კოდირებულია ინფორმაცია საკვლევი ცილის შესახებ. ასეთი ჰიბრიდული, ანუ „შერწყმული“ (fusion) გენის ექსპრესიის (მის მიერ კოდირებული ინფორმაციის „ამოკითხვის“) შედეგად წარმოიქმნება ცილა, რომელიც ინარჩუნებს საკვლევი ცილის თვისებებს, მაგრამ ასევე მანათობელი ცილის თვისებებსაც იძენს. ასეთი ცილის უჯრედსშიდა გადაადგილებისა თუ განლაგების განსაზღვრა შედარებით ადვილია, რადგან ამისათვის საკმარისია მიკროსკოპის ქვეშ მოთავსებულ უჯრედს დავანათოთ ულტრაიისფერი სხივები და ამის შედეგად უჯრედიდან გამომავალ ნათებას დავაკვირდეთ. ამ მეთოდით (რომელსაც ფლუორესცენტული მიკროსკოპია ეწოდება) შესაძლოა ნებისმიერი ცილის ან უჯრედის ფრაგმენტის მორფოლოგიის, დინამიკის, უჯრედის შიგთავსში გადანაცვლების და ა.შ. აღწერა. მრავალნაირი მანათობელი ცილის არსებობის გამო, ასევე შესაძლებელია რამდენიმე ცილის უჯრედსშიდა გადაადგილების ერთდროული დაკვირვება, ისევე როგორც მათი ურთიერთქმედების აღწერა. შექმნილია მეთოდები, რომლებიც ორი სხვადასხვა ცილის ფლუორესცენციაზე დაკვირვების შედეგად განსაზღვრავენ, დაშორებულია თუ არა ეს ორი ცილა 10 ნმ-ზე (10-9 მ) ნაკლებ მანძილზე.
რასაკვირველია, მსგავსი ექსპერიმენტებისათვის აუცილებელია დავრწმუნდეთ, რომ საკვლევი ცილის ფუნქცია არ იცვლება მანათობელი ცილის დამატების შედეგად. მხოლოდ მაშინ ექნება ლოკალიზაციის ექსპერიმენტების დასკვნები სანდო, რადგან ფუნქციის უცვლელობა უფლებას გვაძლევს დიდი ალბათობით ვივარაუდოთ, რომ საკვლევი ცილის უჯრედსშიდა განლაგებაც არ შეცვლილა. საბედნიეროდ, გენური ინჟინერიის მეთოდების გამოყენებით შესაძლებელია, „შერწყმული“ კონსტრუქციის სხვადასხვა ვარიანტები გადაისინჯოს, და თუ საკვლევი და მანათობელი ცილების „გადაბმის“ ერთი ვერსია გამოუსადეგარია, რომელიმე სხვა მისაღები იქნება (მაგ., მანათობელი ცილა საკვლევი ცილის დასაწყისში, შუაში ან ბოლოში). ასეთი ექსპერიმენტები ექსპერიმენტულ ბიოლოგიაში უკვე ჩვეულებრივ მოვლენად იქცა.
მანათობელი ცილების „დეკორატიული“ გამოყენება: ცხოველები, რომლების უჯრედებში მანათობელი ცილა გროვდება, საკმაოდ დიდ შთაბეჭდილებას ახდენენ და რასაკვირველია, დეკორატიულ მნიშვნელობასაც იძენენ. „მანათობელი ცხოველების“ მრავალი მაგალითი არსებობს: მწვანე და იისფერი თაგვები (ნახ. 3), მანათობელი თევზები, ყვითელი მანათობელი ღორი (ნახ. 4) და წითლად მანათობელი კატა. სავარაუდოა, რომ ტექნიკურად ყველაზე რთული და ამავე დროს ყველაზე შთამბეჭდავი მაგალითია 2007 წელს შექმნილი თაგვი, რომლის ტვინის თითოეული ცალკეული უჯრედის „შესაღებად“ 90 სხვადასხვა ფერის მანათობელი ცილიდან ერთ-ერთი იყო გამოყენებული (ნახ. 5). ასეთ ცხოველებს, დეკორატიული თუ „გასართობი“ დანიშნულების გარდა, უდაო პრაქტიკული მნიშვნელობაც აქვთ: მაგალითად, მანათობელი თაგვები წარმატებით გამოიყენება ცოცხალ ცხოველებში ავთვისებიანი სიმსივნის განვითარების შესასწავლად.

ნახ. 3: იისფრად და ცისფრად მანათობელი თაგვები.

ნახ. 4: ღორის ორგანიზმში ყვითელი მანათობელი ცილის (YFP) შეყვანის შედეგად, მისი დინგი და ჩლიქები ყვითელ ფერს იძენს (მარცხნივ). „YFP-ღორის“ გვერდით შედარებისათვის ჩვეულებრივი ღორია ნაჩვენები.
ნახ. 5: „Brainbow“ თაგვის ტვინის უჯრედები. სახელწოდება ორი სიტყვის გაერთიანებითაა მიღებული: Brain (ტვინი) და rainbow (ცისარტყელა - მრავალი ფერის ნათების აღსანიშნავად).
დასკვნა: GFP და სხვა მანათობელი ცილები ევოლუციისა და გენური ინჟინერიის „ერთობლივი მოქმედების“ შესანიშნავი მაგალითია: არსებულ მოლეკულებზე დაყრდნობით, შესაძლებელი გახდა მანათობელი ცილის ახალი ვარიანტების მიღება, რომლებიც ბიოლოგიური კვლევისა თუ დეკორატიული მიზნებისათვის უფრო შესაფერისი გამოდგა. ამ ცილების მრავალფეროვნებამ მნიშვნელოვნად გააადვილა მრავალი ფუნდამენტური ბიოლოგიური პრობლემის შესწავლა და, ზოგიერთ შემთხვევაში, პრობლემის გადაწყვეტაც. დეკორატიული „მანათობელი ცხოველების“ არსებობა (და მათი შეძენის სურვილი) კი იმასაც მიანიშნებს, რომ უნდობლობა გენმოდიფიცირებული ორგანიზმების მიმართ თანდათანობით მცირდება, რაც უეჭველად დადებითი მოვლენაა.
გელა თევზაძე.
24 საათი 01.05.11
|
|
9 პრიონები: კანიბალიზმი ადამიანებში, სიგიჟე ძროხებში და საოცარი გამონაკლისი მოლეკულურ ბიოლოგიაში. |
▲ზევით დაბრუნება |
ბიოლოგიაში არსებობს რამდენიმე კანონი, რომლისაგან გამონაკლისის არსებობა წარმოუდგენელია. მათ შორისაა დებულებები, რომ ყველა ცოცხალი ორგანიზმი უჯრედებისაგან შედგება, რომ ცილები ორგანიზმების სამშენებლო მასალაა, რომ წყალი აუცილებელია სიცოცხლისათვის. ასეთივე „დოგმად“ მიჩნეული იყო, რომ დეზოქსირიბონუკლეინის მჟავაში (დნმ-ში) კოდირებული გენეტიკური ინფორმაციის ამოსაკითხად აუცილებელი იყო ამ ინფორმაციის „გადაწერა“ რიბონუკლეინის მჟავაში (რნმ-ში), შემდეგ კი რნმ-იდან ამ ინფორმაციის „გადათარგმნა“ ცილაში. მოვლენათა ასეთი თანმიმდევრობის საფუძველზე, გენეტიკური ინფორმაციის მემკვიდრეობით გადაცემა მხოლოდ დნმ-ით შეიძლება, და ნებისმიერი ცვლილება ცილაში მემკვიდრეობით გადაიცემა მხოლოდ მაშინ, თუ ეს ცვლილება დნმ-შიც არის კოდირებული. სხვაგვარი ცვლილებები ცილის სტრუქტურაში (მაგ., დაზიანებული ცილა) მემკვიდრეობით არ გადაეცემა, რადგან ეს დაზიანება დნმ-ს არ შეხებია. ამ დებულების უნივერსალობა მხოლოდ მაშინ გახდა საეჭვო, როდესაც მეცნიერებმა ადამიანისა და შინაური ცხოველების რამდენიმე ავადმყოფობის შესწავლისას ძნელად ასახსნელი კანონზომიერებები აღმოაჩინეს.
კურუ და კანიბალიზმი. წინა ფრაზაში ნახსენები დაავადებები - კრუტცფელდტ-ჯაკობის ავადმყოფობა (variant Creutzfeldt-Jakob disease, vJCD) და კურუ (Kuru) - ნერვული სისტემის მძიმე დაზიანებას იწვევს და 6-12 თვის განმავლობაში ფიზიკური და გონებრივი მოქმედების სრული განადგურებითა და სიკვდილით მთავრდება. კურუს ეპიდემია XX საუკუნის 40-50-იან წლებში პაპუა-ახალი გვინეის აღმოსავლეთ კუნძულებზე დაიწყო და, პირვანდელი დაკვირვებით, რიტუალური კანიბალიზმის გამო გავრცელდა. დაავადების მსხვერპლთა 75%25 ზდრასრული ქალები იყვნენ, დარჩენილი 25%25-ს უმეტესობა კი - ბავშვები. მათგან ყველაზე პატარა 4.5 წლისა იყო.
მრავალი წლის განმავლობაში მიმდინარე კანიბალისტური რიტუალების აღმოფხვრის შედეგად კურუს დაავადებების სიხშირემ მნიშვნელოვნად იკლო 1960-იანი წლებიდან, და თითქმის გაქრა დაახლოებით 40 წელიწადში. დაავადების ცალკეული შემთხვევები ამ პერიოდში ისევ კანიბალიზმს უკავშირდებოდა. თუ დავუშვებთ, რომ კანიბალიზმი ამ რეგიონში აღარ გვხვდება, შესაძლოა, ეს შემთხვევები დაავადების გრძელი საინკუბაციო პერიოდით (>30 წლით) აიხსნას.
კურუს კვლევის დასაწყისში, დაავადების გავრცელების ტენდენციაზე დაკვირვებისას, რამდენიმე მცდარი დასკვნა გაკეთდა: რადგან დაავადება ახლო ნათესავებში იყო გავრცელებული და მათი წინაპრების უმეტესობაც კურუთი იყო დაავადებული, ამ დაკვირვებების საფუძველზე წარმოიშვა მოსაზრება, რომ კურუ ერთი თაობიდან მეორეს მემკვიდრეობით გადაეცემოდა რომელიღაც „ინფექციური აგენტის“ (ანუ დნმ-ში კოდირებული ინფორმაციის სახით). ამ დაავადების ხშირ გავრცელებას ქალებში ხსნიდნენ ე.წ. სქესთან შეჭიდულობის მექანიზმით, ანდა გენით, რომლის გამოვლენის ალბათობა ქალის ორგანიზმში გაცილებით მეტი იყო.
პრიონული დაავადებები: მოგვიანებით ცხადი გახდა, რომ კურუს ეპიდემიოლოგიის ზოგიერთი ნიშანი ვერ აიხსნება ამ დაავადების გენეტიკურად (ყოველ შემთხვევაში, „ჩვეულებრივად“ - დნმ-ით) გადაცემით. აღმოჩნდა, რომ დაავადების გავრცელებაში მთავარი როლი ეკუთვნის უჩვეულო ცილას, რომელსაც პრიონი ეწოდება. ამ ცილას გამრავლების უნარი აქვს, თუმცა იგი „ცოცხალი ორგანიზმის“ განსაზღვრებას ვერ აკმაყოფილებს. ადამიანის პრიონი (PrP) განისაზღვრება PRNP გენით, რომელშიც ამ ცილის აგებულების ინფორმაციაა კოდირებული.
პრიონები მრავალ ნერვულ დაავადებას იწვევენ. მათ შორისაა უკვე მოხსენიებული vJCD და კურუ. „პრიონული“ დაავადებების გავრცელება შესაძლოა მემკვიდრეობით (მუტაციები PRNP გენში), მაგრამ არსებობს ამ დაავადებების როგორც ინფექციური, ასევე სპორადული (რაიმე კანონზომიერების გარეშე წარმოქმნილი) შემთხვევებიც. ინფექციური თვისებების წარმოქმნა კი, როგორც აღმოჩნდა, დაკავშირებულია PrP ცილის კონფორმაციის ცვლილებასთან: შესაძლოა, ჯანმრთელი ცილის კონფორმაცია იმგვარად შეიცვალოს, რომ იგი ინფექციური გახდეს, თუმცა მისი შესაბამისი გენი ამ დროს უცვლელი რჩება.
რაც არანაკლებ საინტერესოა, ამ ცილას გააჩნია თავისი „ინფექციური“ კონფორმაციის გავრცელების უნარი: ინფექციური ცილის გავლენით, მის გარშემო მყოფი „ჯანმრთელი“ PrP ცილებიც ავადდებიან (დაწვრილებით იხ. ქვემოთ), და ახლა ისინი ავრცელებენ ამ დაავადებას. ინფექციური ცილა თანდათან გროვდება ორგანიზმში, და საბოლოოდ ნერვული სისტემის სერიოზულ დაზიანებას იწვევს. სწორედ ამ გარემოებით აიხსნება კურუს დაავადების სიხშირე ქალებსა და ბავშვებში: როგორც აღმოჩნდა, კანიბალისტურ რიტუალებში მონაწილეობას მხოლოდ ქალები იღებდნენ, რომლებსაც, ბუნებრივია, პატარა ბავშვებიც ახლდნენ. კურუთი დაავადებული ორგანიზმის ნაწილებთან ერთად მათ სხეულში შედიოდა - და გროვდებოდა - ცილის ინფექციური ფორმა, რომელიც დაავადების განვითარებას აჩქარებდა. თუ ბიჭები ადრეულ ასაკში ამ დავადებას გადაურჩებოდნენ, კურუს განვითარების საშიშროება მნიშვნელოვნად იკლებდა, რადგან მამაკაცები კანიბალისტურ რიტუალებში მონაწილეობას აღარ იღებდნენ და, მაშასადამე, წინაპრებისაგან ინფიცირებული ცილის მიღების საშიშროებასაც იცილებდნენ.
ამგვარად, კანიბალისტური რიტუალების აღმოფხვრის შედეგად დაავადების სიხშირის მნიშვნელოვანი შემცირებაც ადვილი ასახსნელია: კურუთი დაავადებული (და გარდაცვლილი) ადამიანი უკვე აღარ წარმოადგენდა ინფექციის წყაროს, და კურუს გავრცელება მხოლოდ „ჩვეულებრივი“ მემკვიდრეობით გადაცემის გზით (ანუ დნმ-ით) გახდა შესაძლებელი. შეიძლება დავასკვნათ, რომ ამ შემთხვევაში კანიბალიზმზე უარის თქმამ არა მარტო სოციალური პროგრესი, არამედ ჯანმრთელობის გაუმჯობესებაც მოიტანა.
პრიონები ცხოველებში: XX საუკუნის 90-იანი წლების დასაწყისში ევროპაში გაჩნდა საქონლის ნერვული სისტემის დაავადება, რომელიც ძროხებში უცნაურ მოძრაობას (და საბოლოოდ, ისევ სიკვდილს) იწვევდა და რომელიც „ძროხების სიგიჟის ავადმყოფობად“ (Mad Cow Disease) მონათლეს. როგორც აღმოჩნდა, ამ დაავადების გავრცელების მექანიზმი ისეთივე იყო, როგორც კურუს შემთხვევაში, ოღონდ აქ კანიბალიზმი „უნებლიედ“ ხდებოდა: ფერმერები საკვებში ურევდნენ დაკლული საქონლის დაფქულ ნარჩენებს (ტვინი, ძვლები და ა.შ.), რის შედეგადაც ხდებოდა ინფექციური ცილის შეტანა ორგანიზმში და შემდგომში მისი გავრცელება. ასეთივე მექანიზმით ხდებოდა მსგავსი ავადმყოფობის (Scrappie) გავრცელება ცხვრებში. ამ შემთხვევებშიც, როგორც კი ინფექციური პრიონების შემცველი ნარჩენები ცხოველთა საკვებიდან გამოირიცხა, დაავადებათა სიხშირემ საგრძნობლად დაიკლო.
პრიონების გამოწვეული დაავადების მოლეკულური მექანიზმები: ვარაუდი იმის შესახებ, რომ პრიონების ინფექციური თვისებები მხოლოდ ცილებით (და არა დნმ-ით) გადაიცემა, პირველად გამოთქვა სტენლი პრუსინერმა (Stanley Prusiner, University of California, San Francisco), რისთვისაც მას 1997 წლის ნობელის პრემია მიენიჭა. ეს დებულება „მხოლოდ ცილის“ (protein-only) ჰიპოთეზის სახელითაა ცნობილი. პრუსინერი თვლიდა, რომ PrPSc (არანორმალური კონფორმაციის მქონე ცილა) მოქმედებს მის გარშემო მყოფ ჯანმრთელ (ნორმალურად „დაკეცილ“, PrPC) ცილებზე, და აიძულებს მათ, რომ კონფორმაცია შეიცვალონ, ანუ მიიღონ ისეთივე სამგანზომილებიანი სტრუქტურა, როგორიც თვით PrPSc -ს აქვს. ადვილი წარმოსადგენია, რომ ამის შედეგად შეცვლილი კონფორმაციის მქონე ცილები მრავლდებიან - ისინი მოქმედებენ სხვა ჯანმრთელ ცილებზე და მათ კონფორმაციასაც ცვლიან. როგორც უკვე აღვნიშნეთ, განსხვავება ჯანმრთელსა და ინფექციურ ცილებს შორის მხოლოდ მათ კონფორმაციაში, ანუ სამგანზომილებიან სტრუქტურაშია: ამინომჟავების (ცილების სამშენებლო ერთეულების) შემადგენლობისა და მათი თანმიმდევრობის მიხედვით PrPC და PrPSc ცილები არაფრით განსხვავდებიან ერთმანეთისაგან.
ამ მოდელის შესამოწმებლად შეიქმნა ახალი მეთოდი, რომელსაც უწოდეს ცილის არასწორი კონფორმაციის ციკლური გამრავლება (protein misfolding cyclic amplification, PMCA). PMCA-ს გამოყენებით, შესაძლებელი გახდა PrPSc -ცილების სინჯარაში გამრავლება. მეთოდის არსი შემდეგშია: პრიონებით ინფიცირებული თაგვის ტვინი იმგვარად დამუშავდება, რომ ყველა უჯრედი დაიშალოს და ე.წ. „უუჯრედო“ ჰომოგენური სითხე წარმოიქმნას. როდესაც ამ სითხეს შეურევენ ჯანმრთელი თაგვის ტვინისაგან მიღებულ სითხეს, დაავადებულ ტვინში არსებული PrPSc-ცილები გარდაქმნიან ყველა ჯანმრთელ PrPC-ს PrPSc-დ. შემდეგ ამ ნარევს ხსნიდნენ ჯანმრთელი ტვინისაგან მიღებულ სითხეში, და პროცესს იმეორებდნენ. ასობით ციკლისა და 10-20-ჯერ გახსნის შედეგად ამ ნარევში ერთი მოლეკულაც კი არ უნდა დარჩენილიყო თავდაპირველად გამოყენებული დაავადებული ტვინისაგან. ამის მიუხედავად, თუ ნარევს ჯანმრთელ თაგვს შეუშხაპუნებდნენ, იგი კვდებოდა პრიონებით გამოწვეული ავადმყოფობისაგან. ამრიგად, ახალი PrPSc-ცილები შეიძლება გაჩნდეს „უუჯრედო“ გარემოში, და მათ წარმოსაქმნელად PrPSc-ს მხოლოდ რამდენიმე მოლეკულის არსებობაა საჭირო.
ნახ. 1: პრიონის არასწორი კონფორმაციის ციკლური გამრავლება.
ამავე ექსპერიმენტმა დაადასტურა, რომ მცდარია ალტერნატიული მოდელი, რომელიც ამტკიცებს, რომ პრიონულ დაავადებებს გარკვეული სახის ვირუსები იწვევენ: ვირუსების გამრავლებისათვის უჯრედების არსებობაა აუცილებელი, ამ ცდაში კი PrPSc-ს წარმოქმნა „უჯრედებისაგან თავისუფალ“ (cell-free) გარემოში ხდებოდა.
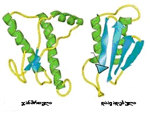
ნახ. 2: ჯანმრთელი და ავადმყოფობის გამომწვევი (PrPC და PrPSc) ცილების სტრუქტურა.
რამდენიმე კითხვა კვლავ უპასუხოდ რჩება. მაგალითად, რამდენად დარწმუნებული ვართ, რომ ზემოთაღწერილ ექსპერიმენტში „უუჯრედო“ გარემო არცერთ უჯრედს არ შეიცავდა? ეჭვების საბოლოოდ გასაფანტავად, საჭიროა ამ ექსპერიმენტში არა ავადმყოფი ცხოველისაგან მიღებული, არამედ ქიმიურად სინთეზირებული PrPSc-ს გამოყენება. ამგვარად მოდიფიცირებული ცდა საბოლოოდ დაასაბუთებდა, რომ პრიონულ დაავადებებში ერთადერთი ინფექციური აგენტი სწორედ PrPSc-ცილაა და არა დნმ (ან ჯერაც უცნობი ვირუსები).
ნახ. 3: PrPC-ს გარდაქმნა PrPSc-დ.
ასევე, გაურკვეველია, როგორ გაივლის კვების დროს მიღებული PrPSc მომნელებელი სისტემას და აღწევს ტვინის უჯრედებში. ამ შემთხვევაში ორი შესაძლო მექანიზმი არსებობს: 1) თავის ტვინში უშუალოდ ის მოლეკულები შედიან, რომლებმაც საკვებთან ერთად შეაღწიეს ორგანიზმში; 2) PrPSc, რომლებმაც ორგანიზმში შეაღწიეს, ჯერ თავის მეზობლებს აავადებენ პერიფერულ ნერვულ სისტემაში, და ახალდაავადებული მოლეკულები სხვებზე მოქმედებენ, რის შედეგადაც „საბოლოო მიზანს“ - თავის ტვინის ინფიცირება - ეტაპობრივად აღწევენ? ამ პრობლემების გადაჭრა შესაძლებელი უნდა გახდეს დამატებითი კვლევების შედეგად.

ნახ. 4: პრიონული „სიგიჟით“ დაავადებული ძროხა.
დასკვნა: თუმცა პრიონებით გამოწვეული დაავადებების წარმოშობისა და გავრცელების მოლეკულური მექანიზმები სრულყოფილად არაა შესწავლილი, უკვე საკმარისი ცოდნაა დაგროვილი, რომ ამ დაავადებების გავრცელებას ხელი შეეშალოს. მაგალითად, ფერმებში ცხოველთა ნარჩენების საკვებად გადამუშავების აკრძალვამ, რომელიც ინგლისში დაკანონდა, მნიშვნელოვნად შეუწყო ხელი „საქონლის სიგიჟის“ შემთხვევების შემცირებას. ასევე, ევროპის ზოგიერთ ქვეყანაში საქონლის ხორცის დამზადების წინ რეკომენდირებულია საქონლის ზურგის ტვინის ამოცლა, რადგან ზურგის ტვინში პრიონების მაქსიმალური კონცენტრაციაა მოსალოდნელი. და ბოლოს, რაც მეცნიერებისათვის არანაკლებ მნიშვნელოვანია, პრიონების შესწავლის შედეგად აღმოჩენილი იყო გენეტიკური ინფორმაციის გადაცემის უაღრესად საინტერესო მექანიზმი, რომლის ერთადერთ მაგალითად ჯერ-ჯერობით თვით პრიონები რჩებიან.
გელა თევზაძე.
24 საათი 15.05.11
|
|
10 უჯრედის ციკლი: მოძრაობისა და შეჩერების განსაზღვრული მონაცვლეობა. |
▲ზევით დაბრუნება |
ზუსტად ათი წლის წინ, 2001 წელს, ნობელის პრემიის ლაურეატი ფიზიოლოგიასა და მედიცინაში სამი მეცნიერი გახდა: ლილანდ ჰარტველი, ტიმოთი ჰანტი და პოლ ნერსი (Leland H. Hartwell, Fred Hutchinson Cancer Research Center, Seattle, USA; R. Timothy (Tim) Hunt, Imperial Cancer Research Fund, London; Paul M. Nurse, Imperial Cancer Research Fund, London). პრემიის მინიჭებით აღიარებული იქნა მათი წვლილი „უჯრედის ციკლის საკვანძო რეგულატორების აღმოჩენაში“. ეს სტატია მათ აღმოჩენებს ეძღვნება.

ნახ.1: 2001 წლის ნობელის პრემიის ლაურეატები: ლილანდ ჰარტველი, ტიმოთი ჰანტი და პოლ ნერსი (Leland Hartwell, Tim Hunt, Paul Nurse)
როგორც ცნობილია, ყველა ორგანიზმი უჯრედისაგან შედგება. ეს უჯრედები გაყოფის გზით მრავლდებიან. ზრდასრული ადამიანის ორგანიზმში 100 000 მილიარდი უჯრედია, და ყველა მათგანი ერთადერთი უჯრედიდან - განაყოფიერებული კვერცხუჯრედიდან - იღებს სათავეს. ზრდასრულ ორგანიზმშიც კი ძალიან ბევრი უჯრედია, რომელიც მუდმივად იყოფა და დაღუპული უჯრედების ადგილს იკავებს. სანამ უჯრედი გაყოფას დაიწყებს, ის უნდა გაიზარდოს (ანუ, სხვაგვარად, მიაღწიოს გარკვეულ ზომას), გააორმაგოს თავისი ქრომოსომები (გენეტიკური ინფორმაციის - დნ მჟავის შემცველი ერთეულები) და გაყოს გაორმაგებული ქრომოსომები იმგვარად, რომ ისინი თანაბრად გადანაწილდნენ ორ ახალწარმოქმნილ უჯრედში. უჯრედის ციკლის განმავლობაში სწორედ ამ მრავალი პროცესის რეგულაცია და კოორდინაცია ხდება.
2001 წლის ნობელის პრემიის ლაურეატებს ეკუთვნით უმნიშვნელოვანესი აღმოჩენები უჯრედის ციკლის გენეტიკური კონტროლის კვლევის დარგში. ამ აღმოჩენებიდან უპირველესია მოლეკულები, რომლებიც უჯრედის ციკლის კონტროლს უზრუნველყოფენ ყველა ეუკარიოტულ ორგანიზმში (ანუ ისეთ ორგანიზმში, რომლის უჯრედები ბირთვს შეიცავს), მათ შორის საფუარის სოკოებში, მცენარეებში, ცხოველებსა და ადამიანებში. კონტროლის ფუნქციას ეს მოლეკულები უჯრედის ციკლის ყველა ეტაპზე ასრულებენ. უჯრედის ციკლის დეფექტებმა შეიძლება გამოიწვიოს ავთვისებიანი სიმსივნის განვითარება. ეს კი, თავის მხრივ, კიბოს წარმოშობის მექანიზმების გამორკვევისა და მათი განკურნების შესაძლო სქემების გამომუშავების პერსპექტივას წარმოშობს.
ნახ. 2: უჯრედის ციკლის სქემა: G1 - ზრდის სტადია; S - დნმ-ის სინთეზი; G2 - დნმ-ის სინთეზის შემდგომი ზრდა; M - უჯრედის გაყოფა.
ნახ. 3: ადამიანის უჯრედის ციკლის სტადიების ფოტოგრაფია.
ლილანდ ჰარტველის დამსახურება უჯრედის ციკლის რეგულატორების აღმოჩენაა. ერთ-ერთი, რომელიც სახელით „START“აღინიშნება, უჯრედის ციკლის პირველი ეტაპის კონტროლში გადამწყვეტ ფუნქციას ასრულებს. ჰარტველმა ასევე განსაზღვრა „checkpoint“-ის ცნება, რაც მეტად მნიშვნელოვანი იყო უჯრედის ციკლის ამოქმედების მექანიზმის გაგებას.
პოლ ნერსმა გამოყო და დაახასიათა СDK (cyclin-dependent kinase) - უჯრედის ციკლის ერთ-ერთი უმნიშვნელოვანესი რეგულატორი. ნერსმა არგეთვე აჩვენა, რომ ეს ცილა ევილუციის განმავლობაში კონსერვატულია (ანუ ერთი და იგივე შემადგენლობა აქვს იმისდა მიუხედავად, თუ რომელი ორგანიზმიდანაა გამოყოფილი. СDK სხვადასხვა ცილებს ამატებს ფოსფატს (ფოსფორისა და ჟანგბადის ნაერთს), რაც უჯრედის ციკლის „წინ წაწევის“ სიგნალი ხდება.
ტიმოთი ჰანტმა აღმოაჩინა ციკლინები - ცილები, რომლებიც СDK-ს რეგულატორები არიან, ყოველი უჯრედის გაყოფის გარკვეულ სტადიაზე ციკლინების დეგრადაცია ხდება. როგორც აღმოჩნდა, ეს მექანიზმი უაღრესად მნიშვნელოვანია უჯრედის ციკლის რეგულაციისათვის.
ეუკარიოტული უჯრედები, რომლებშიც ქრომოსომები ბირთვშია მოთავსებული და ამრიგად გამოყოფილია უჯრედის დანარჩენი ნაწილებისაგან, დედამიწაზე დაახლოებით ორი მილიარდი წლის წინ გაჩნდა. ასეთი უჯრედების შემცველი ორგანიზმები შეიძლება იყვნენ როგორც ერთუჯრედიანები (მაგ., საფუარი, ამება), ასევე მრავალუჯრედიანები (მცენარეები, ცხოველები). ადამიანის სხეულში ძალიან ბევრი უჯრედია - საშუალოდ ერთი მოლიარდი ერთ გრამში.
ის ფაქტი, რომ უჯრედების გამრავლება მათი გაყოფის საშუალებით ხდება, ცნობილია უკვე საუკუნეზე მეტი ხნის განმავლობაში. უჯრედის გაყოფისა და, ზოგადად, უჯრედის ციკლის მოლეკულური მექანიზმების შესწავლა კი მხოლოდ ბოლო სამი ათწლეულის განმავლობაში ხდება. განვიხილოთ ეს მექანიზმები:
უჯრედის ციკლის კონტროლი: ყველა ცოცხალი ორგანიზმისათვის აუცილებელია, რომ მათი უჯრედების ციკლის სხვადასხვა ფაზების (სტადიების) კოორდინაცია ზედმიწევნითი სიზუსტით ხდება. ფაზების თანმომდევრობა უცვლელი უნდა იყოს, და მხოლოდ მაშინ, როდესაც რომელიმე ფაზა ბოლომდეა დასრულებული, შესაძლებელია მისი მომდევნო ფაზის დაწყება. ამ კოორდინაციის შენარჩუნება აუცილებელია, რომ უჯრედის ფუნქციები და, საბოლოოდ, მისი ერთიანობა არ დაირღვეს. მაგალითად, ქრომოსომების რაოდენობის არათანაბარმა განაწილებამ ორ უჯრედს შორის, რაც ფაზებს შორის კოორდინაციის დაკარგვის ერთ-ერთი შესაძლო შედეგია, შესაძლოა ავთვისებიანი სიმსივნის გამვითარება გამოიწვიოს.
როგორც ბიოლოგიის, ასევე მედიცინის განვითარებისათვის ძალიან მნიშვნელოვანია უჯრედის ციკლის კონტროლისა და კოორდინაციის შესწავლა. 2001 წლის ნობელი პრემიის ლაურეატებმა აღმოაჩინეს რამდენიმე მნიშვნელოვანი მოლეკულური მექანიზმი, რომელთა საშუალებით ხდება უჯრედის გადასვლა ციკლის ერთი ფაზიდან მეორეში.
უჯრედის ციკლის გენები საფუარის უჯრედებში: გასული საუკუნის 60-იან წლებში ლილანდ ჰარტველმა დაიწყო უჯრედის ციკლის შესწავლა გენეტიკური მეთოდების გამოყენებით. თავისი ექსპერიმენტებისათვის მან აირჩია საფუარი (Saccharomyces cerevisiae) - ერთუჯრედიანი სოკო, რომლის მრავალი უპირატესობა უკვე ცნობილი იყო: საფუარი ეუკარიოტია (ამიტომ მისი უჯრედების აგებულება ძალიან ჰგავს ადამიანისას), მაგრამ ამავე დროს ერთუჯრედიანი ორგანიზმია, რაც მისი უჯრედების ზრდის სისწრაფესა და, მაშასადამე, ექსპერიმენტების შედარებით მოკლე ვადას განსაზღვრავს. საფუარის უჯრედების კვლევისათვის ჯერ კიდევ მაშინ არსებობდა მრავალი გენეტიკური და მიკრობიოლოგიური მეთოდი, რამაც ასევე განსაზღვრა ამ ორგანიზმის, როგორც უჯრედის ციკლის შესწავლისათვის თითქმის იდეალური მოდელური სისტემის, არჩევანი.
1970-1971 წლებში ჩატარებული ექსპერიმენტების შედეგად ჰარტველმა მიიღო საფუარის მუტანტური უჯრედები, რომლებშიც უჯრედის ციკლის ჩვეულებრივი მსვლელობა დარღვეული იყო. თანაც, როგორც აღმოჩნდა, სხვადასხვა უჯრედებში ეს დარღვევა სხვადასხვაგვარად გამოიხატებოდა: როგორც შემდგომში აღმოჩნდა, ყოველ ცალკეულ შემთხვევაში მუტაცია განსხვავებულ გენში იყო „მოთავსებული“. ამრიგად, გამოირკვა, რომ უჯრედის ციკლის კონტროლი ხდება გენების ერთობლიობით, და ამ გენებიდან თითოეული მნიშვნელოვანია პროცესის ნორმალური წარმართვისათვის. ყველა ამ გენის სახელწოდება შეიცავს სიტყვას CDC, რაც „უჯრედის გაყოფის კონტროლს“ (Cell Division Control) ნიშნავს. ერთ-ერთი ასეთი გენი, რომელსაც ჰარტველმა CDC28 უწოდა, უჯრედის ციკლის პირველ სტადიას მართავს და ამის გამო ასევე მოიხსენიება, როგორც „START“.
ჰარტველი ასევე იკვლევდა გამოსხივების გავლენას საფუარის უჯრედებზე. ამ ექსპერიმენტების საფუძველზე მან ჩამოაყალიბა „checkpoint“-ის (საყარაულო პუნქტის) კონცეფცია. უჯრედის checkpoint აჩერებს ციკლის განვითარებას მაშინ, როდესაც დნ მჟავა დაზიანებულია. ასეთი შეჩერების არსი იმაში მდგომარეობს, რომ უჯრედს საშუალება ეძლევა, დაზიანებული დნმ შეაკეთოს და შემდეგ გადავიდეს ციკლის მორიგ ფაზაზე. ჰარტველმა checkpoint-ის კონცეფცია განავრცო კონტროლის იმ მექანიზმებზეც, რომლებიც ზოგადად უზრუნველყოფენ ციკლის სტადიების სწორ თანმიმდევრობას.
პოლ ნერსი თავის ექსპერიმენტებში უჯრედის ციკლის პროცესებს გენეტიკური მეთოდების გამოყენებით შეისწავლიდა. კვლევის ობიექტი აქაც საფუარი იყო, მაგრამ სხვა სახეობისა - Schizosaccaromyces pombe. ამ საფუარის უჯრედები ე.წ. „თანაბარი გაყოფით“ მრავლდება, როდესაც ერთი უჯრედი ორი თანაბარი ზომის შვილეულ უჯრედს წარმოქმნის. ასეთი გაყოფა მას განასხვავებს ჰარტველის მიერ გამოყენებული Saccharomyces cerevisiae-საგან, რომელიც „გაკვირტვით“ მრავლდება: უჯრედს გამოეყოფა უფრო მცირე ზომის უჯრედი („კვირტი“), რომელიც გენეტიკური ინფორმაციის იგივე რაოდენობას შეიცავს, რაც „დედა უჯრედშია“. ვარაუდობენ, რომ ევოლუციის პერიოდში ამ ორ საფუარს ბოლო საერთო წინაპარი დაახლოებით ერთი მილიარდი წლის წინ ჰყავდა, მას შემდეგ კი ისინი ერთმანეთისაგან დამოუკიდებლად ვითარდებიან და, მაშასადამე, მხოლოდ „შორეული ნათესავები“ არიან.
1970-იან წლებში ნერსმა აღმოაჩინა Schizosaccaromyces pombe-ს გენი, რომელსაც cdc2 უწოდა. როგორც გაირკვა, ეს გენი აკონტროლებს უჯრედის ციკლის ორ სხვადასხვა პროცესს: ზრდის სტადიიდან დნმ-ის სინთეზზე გადასვლას (რითაც ის ჰარტველის აღმოჩენილი CDC28-ს მსგავსია), და დნმ-ის სინთეზისშემდგომი ზრდის სტადიის შემდეგ უშუალოდ უჯრედის გაყოფის დაწყებას. 1987 წელს კი ნერსმა გამოყო გენი, რომელსაც იგივე ფუნქცია აქვს ადამიანის ორგანიზმში და რომელიც მოგვიანებით CDK1 (cyclin-dependent kinase, ციკლინზე დამოკიდებული კინაზა) ეწოდა. ამ გენში კოდირებული ინფორმაციით განისაზღვრება ცილა (კინაზა), რომლის ამოქმედება (აქტივაცია) დამოკიდებულია ამ ცილის მოლეკულასთან ფოსფატის (ფოსფორისა და ჟანგბადის ატომების ერთობლიობის) მიერთებაზე. ამ პროცესით, ანუ ფოსფატის მიერთებითა თუ მოცილებით. მრავალი ცილის მოქმედების კონტროლი ხდება. კინაზის მოქმედება, ერთი მხრივ, ფოსფატის მიერთებაზეა დამოკიდებული, მეორე მხრივ კი ამ ცილას თვითონ შეუძლია სხვა ცილების აქტივაცია მათთვის ფოსფატის მიერთების გზით. „ციკლინზე დამოკიდებულება“ მიანიშნებს, რომ CDK-ს მოქმედება უჯრედის ციკლის სტადიაზეა დამოკიდებული. ადამიანის ორგანიზმში რამდენიმე CDK-ცილაა, მაშინ როდესაც საფუარში ამ ფუნქციას ერთადერთი ცილა ასრულებს (CDC28 და cdc2 შესაბამისად, Saccharomyces cerevisiae-ში და Schizosaccaromyces pombe-ში).
პირველი ციკლინის აღმოჩენა: ტიმოთი ჰანტმა პირველი ციკლინი 1980-იან წლებში აღმოაჩინა. ციკლინები ცილებია, რომლებიც წარმოიქმნებიან და იშლებიან უჯრედის ციკლის გარკვეული სტადიების განმავლობაში.თვით სახელწოდება „ციკლინები“ მიანიშნებს სწორედ მათ დამოკიდებულებას უჯრედის ციკლზე. ციკლინები უერთდებიან CDK-ცილებს, რის შედეგადაც ხდება CDK-ს მოქმედების რეგულაცია და იმ „სამიზნე“ მოლეკულების არჩევა, რომლებსაც ფოსფატი უნდა მიუერთდეს. ჰარტველის და ნერსის კვლევებისაგან განსხვავებით, ჰანტი თავისი ექსპერიმენტების მოდელურ ორგანიზმად ზღვის ზღარბს იყენებდა. შემდგომ, ციკლინები ბევრ სხვა ორგანიზმში აღმოაჩინეს. ადამიანის უჯრედებში ათამდე სხვადასხვა ციკლინია.
ზოგადად, ცილის პერიოდული დაგროვება უჯრედებში და შემდეგ მისი დეგრადაცია უჯდერის ციკლის რეგულაციის მნიშვნელოვანი მექანიზმია: CDK-ს რაოდენობა უჯრედში უცვლელია, მაგრამ მათი მოქმედება განისაზღვრება ციკლინების მიერთებით. ციკლინების რაოდენობა კი უჯრედის ციკლის სტადიაზეა დამოკიდებული. ეს ერთიერთქმედება შეიძლება ავტომანქანის ძრავასა და სიჩქარის გადამრთველ კოლოფს შევადაროთ: ძრავა (CDK) შეიძლება მუდმივად მუშაობდეს, მაგრამ სიჩქარის გადამრთველი (ციკლინები) განსაზღვრავს მანქანის მოძრაობასა თუ შეჩერებას.
დასკვნა: უჯრედის ციკლის კვლევას მრავალი პრაქტიკული გამოყენება აქვს, უპირველეს ყოვლისა სამედიცინო თვალსაზრისით. ქრომოსომების არასწორი გადანაწილება უჯრედებს შორის, მათი რაოდენობის სტაბილურობის დარღვევა, და სხვა გენეტიკური ცვლილებები უჯრედის ციკლის დარღვევებთანაა დაკავშირებული. ასევე, აღმოჩნდა, რომ მუტაციები CDK და ციკლინების გენებში, ისევე როგორც შესაბამისი ცილების გადაჭარბებული რაოდენობა, შეიძლება ავთვისებიანი სიმსივნის განვითარების წინაპირობად იქცეს. ამ მექანიზმების შესწავლის საფუძველზე შესაძლოა კიბოს საწინააღმდეგო მედიკამენტების შექმნის განსხვავებული სტრატეგიის შემუშავებაც.
გელა თევზაძე.
24 საათი 12.06.11
|
|
11 იმუნური სისტემა: ცვლადი და მუდმივი კომპონენტების ერთობლიობა. |
▲ზევით დაბრუნება |
ადამიანის ორგანიზმი დაბადებიდანვე (თუ ჩასახვიდანვე არა) განიცდის გარემო ფაქტორების გავლენას, და ამ ფაქტორების დიდი ნაწილი საზიანოა. ზიანის მომტანი ფაქტორებიდან ზოგიერთი იწვევს სხვადასხვა დაავადებას, რომლის წინააღმდეგ ბრძოლა შეუძლებელი იქნებოდა, რომ არ არსებობდეს დაცვითი მექანიზმი, რომელიც ავადმყოფობის გამომწვევ ორგანიზმებს (ბაქტერიები, ვირუსები) აუვნებლებს ან მათ მოქმედებას ანელებს. ეს მექანიზმი იმუნური სისტემის სახელითაა ცნობილი, მისი უმთავრესი ნაწილი კი ანტისხეულებია, რომელსაც ორგანიზმი უცხო სხეულის ამოცნობისას გამოიმუშავებს.
რა აგებულებისაა და როგორ მოქმედებს ანტისხეული?

ნახ.2: ანტისხეულების მიერთება ვირუსის ნაწილაკთან
სახელწოდება „ანტისხეული“ (გერმ. Antikörper) პირველად გამოიყენა პაულ ერლიხმა (Paul Ehrlich) 1891 წელს თავის სტატიაში, სადაც აღნიშნა: „თუ ორი ნაერთი ორი სხვადასხვა ანტისხეულის წარმოქმნას იწვევს, მაშინ ეს ნაერთებიც არაა ერთნაირი“. ამ წინადადებაში მოქცეულია იმუნოლოგიის არსი: ნებისმიერ უცხო სხეულზე, რომელი ორგანიზმში შეიჭრება, ორგანიზმი რეაგირებას ახდენს მხოლოდ ამ სხეულისათვის სპეციფიური ანტისხეულების გამომუშავების გზით. იმის გასააზრებლად, თუ რამდენად მოქნილია ეს სისტემა, საკმარისია გავაცნობიეროთ, რომ იმუნურ სისტემას ძალუძს, ნებისმიერი („სრულიად უცხო/აქამდე უნახავი“) ანტიგენის მიმართ გამოიმუშავოს ანტისხეულები.
ნახ. 1: ანტისხეულის სტრუქტურა: ანტიგენის მიერთების ადგილი („Y-ის განშტოებები“); ცვლადი (ღია ფერის) და მუდმივი (მუქი ფერის) ნაწილები; მსუბუქი (ვარდისფერი) და მძიმე (ლურჯი) ჯაჭვები.
ანტისხეულები, რომლებიც ასევე იმუბოგლობულინების სახელით მოიხსენიება, დიდი ზომის ცილებია. ანტისხეულების საშუალებით ადამიანის იმუნური სისტემა ცნობს და აუვნებლებს ორგანიზმში შეჭრილ უცხო ორგანიზმებს (მაგ., ბაქტერიები, ვირუსები). ანტისხეული ცნობს უცხო ორგანიზმის რომელიმე ნაწილს, რომელიც განისაზღვრება, როგორც ანტიგენი.
ანტისხეულები ასო Y-ის ფორმის ცილებია. „Y“-ის განშტოებული ბოლოები ცნობენ ანტიგენს; „Y“-ის ბოლოებისა და ანტიგენის ურთიერთქმედება შეიძლება ბოქლომსა და გასაღებს შევადაროთ: თითოეული ანტისხეული მხოლოდ ერთი სახის ანტიგენს ცნობს, და ამგვარი სპეციფიურობა მიერთების დიდ სიზუსტეს უზრუნველყოფს. ანტისხეულის მიერთების შედეგად ანტიგენი (მაგ., ბაქტერია) „მოინიშნება“ და ხილული ხდება იმუნური სისტემის სხვა ნაწილების (მაგ., ლიმფოციტების) მიერ. რომლებიც ამ სამიზნეს ანადგურებენ. ანტისხეულებს აგრეთვე ძალუძთ ანტიგენის უშუალო გაუვნებლება - თუნდაც ბაქტერიის იმ ნაწილის ბლოკირებით, რომელიც აუცილებელია მასპინძელ ორგანიზმში შეჭრისათვის და იმ გამრავლებისათვის.
ანტისხეულები წარმოიქმნება სისხლის თეთრი უჯრედების ერთ-ერთი ნაირსახეობის მიერ, რომელსაც პლაზმური უჯრედები ეწოდება. ანტისხეულები ორი სხვადასხვა ფორმით არსებობს: ერთი მათგანი წყალში ხსნადია და უჯრედების მიერ გამოიყოფა „თავისუფლად მოძრავი“ ფორმის სახით; მეორე მიმაგრებულია ლიმფოციტების ერთ-ერთი სახეობის (B-უჯრედების) ზედაპირზე და ასევე მოიხსენიება, როგორც B-უჯრედების რეცეპტორი (B-cell receptor, BCR).
ხსნადი ანტისხეულები სისხლში მოძრაობენ, მათი ფუნქცია სხეულში შეჭრილი მიკროორგანიზმების აღმოჩენაა. BCR-რეცეპტორები ხელს უწყობენ B-უჯრედების აქტივირებას და მათ გარდაქმნას პლასმურ უჯრედებად ან ე.წ. „მეხსიერების B-უჯრედებად“ (memory B-cells). „მეხსიერების უჯრედები“ ორგანიზმში დიდი ხნის განმავლობაში რჩებიან, რის შედეგადაც იგივე ანტიგენთან განმეორებითი შეხვედრისას ორგანიზმი უფრო სწრაფ რეაქციას ახერხებს. სწორედ ამ მექანიზმზეა დაფუძნებული იმუნიზაციის („აცრის“) პრინციპი.
ანტისხეულები ორი ძირითადი ერთეულისგან შედგება: ორი დიდი ზომის მძიმე ჯაჭვისაგან და ორი მცირე მსუბუქი ჯაჭვისაგან. მძიმე ჯაჭვის რამდენიმე განსხვავებული ნაირსახეობა არსებობს, და ანტისხეულების კლასიფიკაცია სწორედ ამის საფუძველზე ხდება. ძუძუმწოვრებში 5 განსხვავებული სახის ანტისხეული (ე.წ. იზოტიპი) არსებობს, რის გამოც ნებისმიერი ანტიგენის წინააღმდეგ ყველაზე ეფექტური იზოტიპის მიმართვაა შესაძლებელი.
თუმცა ანტისხეულების ზოგადი აგებულება მეტ-ნაკლებად ერთგვაროვანია, მისი ბოლოები („Y“-ის განშტოებები) უაღრესად მრავალფეროვანია. ამ ბოლოების შემადგენელი ცილები სტრუქტურის ცვლილებამ შესაძლოა რამდენიმე მილიონი განსხვავებული ანტისხეული წარმოქმნას. ეს, თავის მხრივ, უზრუნველყოფს ანტიგენების დიდი რაოდენობის გამოცმობას და მათ წინააღმდეგ მოქმედებას. ანტისხეულებს აგრეთვე შეუძლიათ მძიმე ჯაჭვების ერთმანეთში მიმოცვლა, რაც, ბუნებრივია, მათი იზოტიპის ცვლილებას იწვევს. რადგან ცვლადი ნაწილი ამ დროს შენარჩუნებულია, ერთი და იგივე სპეციფიურობის ანტისხეული იმუნური სისტემის სხვადასხვა ნაწილში გამოიყენება.
ანტისხეულების ფორმები: ზედაპირული იმუნოგლობულინები (Ig) მიმაგრებულია B-უჯრედების გარსზე ტრანსმემბრანული ფრაგმენტის მეშვეობით. ანტისხეულები, რომლებიც სისხლში მოძრაობენ, Ig-ს იდენტურია, მაგრამ ტრანსმემბრანული ფრაგმენტი არ გააჩნიათ. სწორედ ამ განსხვავებით განისაზღვრება ანტისხეულების ორი ფორმა: გარსზე (მემბრანაზე) მიმაგრებული და ხსნადი.
მემბრანაზე მიმაგრებულ ანტისხეულს ზედაპირული იმუნოგლობულინი (surface immunoglobulin, sIg) ან მემბრანული იმუნოგლობულინი (membrane immunoglobulin, mIg) ეწოდება. ეს ანტისხეულები B-უჯრედების რეცეპტორის ნაწილია. მათი საშუალებით B-უჯრედებს სხეულში არსებული ანტიგენის აღმოჩენა შეუძლიათ. B-უჯრედების ზედაპირზე ჩვეულებრივ 50 ათასიდან 100 ათასამდე ანტისხეულია მიმაგრებული.
იზოტიპები: ძუძუმწოვრების ანტისხეულები, როგორც აღვნიშნეთ, 5 სხვადასხვა სახეობად (იზოტიპად) იყოფა: IgA, IgD, IgE, IgG და IgM. „Ig“ ყველა სახელწოდებაში იმუნოგლობულინის აღმნიშვნელია; იზოტიპები განსხვავდება ბიოლოგიური თვისებების, ორგანიზმში მდებარეობის, და სხვადასხვა ანტიგენთან ურთიერთქმედების უნარის მიხედვით. B-უჯრედებთან მიერთებული ანტისხეულების იზოტიპი უჯრედის განვითარებისა და სპეციალიზაციის განმავლობაში იცვლება. მოუმწიფებელი B-უჯრედები, რომლებიც ანტიგენს ერთხელაც არ „შეხვედრიან“, გამოყოფენ მხოლოდ IgM იზოტიპის მემბრანასთან მიმაგრებულ ფორმას. სიმწიფის მიღწევისას, B-უჯრედები როგორც IgM, ასევე IgD ანტისხეულებს, და ამ დროს უკვე ანტიგენთან ურთიერთქმედებისათვის მზად არიან. შემდგომ, როდესაც ანტისხეული და ანტიგენი ერთმანეთს უკავშირდება, ხდება B-უჯრედების აქტივაცია, და ამ უჯრედიდან პლაზმური უჯრედი წარმოიქმნება, რომელსაც უკვე თავისუფლად მოძრავი ანტისხეულების გამოყოფის უნარი აქვს. აქტივირებული B-უჯრედების ერთი ნაწილი იზოტიპს იცვლის, რის შედეგადაც , IgM და IgD იზოტიპების ანტისხეულები გარდაიქმნება IgA, IgE, ან IgG ანტისხეულებად. ამ ანტისხეულებს განსხვავებული ფუნქციები აქვთ, მაგალითად: IgA ხელს უშლის სხვადასხვა მიკროორგანიზმების გამრავლებას; IgE ებმის სხვადასხვა სახის ალერგენს და ასევე იცავს ორგანიზმს სხვადასხვა პარაზიტული ჭიებისაგან; IgG უზრუნველყოფს პათოგენების წინააღმდეგ ორგანიზმის რეაგირებას, ისევე როგორც ჩანასახის იმუნურ დაცვას ფეხმძიმობის დროს.
ანტისხეულების სტრუქტურა: ანტისხეულები დიდი ზომის ცილებია, რომლების ზოგიერთ ფრაგმენტზე შაქრის მოლეკულებია მიოერთებული ასეთი ცილები განისაზღვრება, როგორც გლიკოპროტეინები. ანტისხეულის ძირითადი შემადგენელი ერთეული იმუნოგლოგულინის მონომერია (ანუ ის Ig-ს მხოლოდ ერთ ერთეულს შეიცავს). ზოგიერთ შემთხვევაში, თავისუფლად მოძრავი ანტისხეულები შეიძლება რამდენიმე (ორ, ოთხ ან ხუთ) ერთეულს შეიცავდნენ. ანტისხეულის ცვლად ნაწილს („Y-ის განშტოებებს“) V-ფრაგმენტი (variable) ეწოდება, ხოლოდ მუდმივ ნაწილს - C-ფრაგმენტი (constant).
იმუნოგლობულინების ფუნქციები: ჩანასახის განვითარების დროს, ისევე როგორც დაბადების პირველ დღეებში, ანტისხეულების მიწოდება ხდება ე.წ. პასიური იმუნიზაციის გზით დედის ორგანიზმიდან. ანტისხეულების მიერ იმუნიტეტის უზრუნველყოფა ხდება ავადმყოფობის გამომწვევ ორგანიზმებთან (ბაქტერიები, ვირუსები) მიერთების გზით, რის შედეგადაც: 1) მნიშვნელოვნად ფერხდება პათოგენის შეჭრა უჯრედებში და მათი დაზიანება; 2) ხდება სპეციალიზირებული უჯრედების აქტივაცია, რომლებიც პათოგენებს ამოიცნობენ და ანადგურებენ; 3) ასევე აქტიურდება იმუნური რეაქციის მექანიზმები, რომელთა შორის ერთ-ერთი უმნიშვნვნელოვანესია კომპლემენტის ბმის რეაქცია.
კომპლემენტის ბმის რეაქცია: კომპლემენტის სისტემა დამხმარე, ანუ „კომპლემენტარულია“ ორგანიზმის პათოგენებისაგან განთავისუფლების პროცესში, რომელსაც ძირითადად ანტისხეულები და სპეციალიზირებული „შთანმთქმელი უჯრედები“ (ფაგოციტები) ასრულებენ. კომპლემენტის სისტემა შედგება მცირე ზომის ცილებისაგან, რომლებსაც ღვიძლი გამოიმუშავებს. ეს ცილები სისხლში ყოველთვის იმყოფებიან, მაგრამ მათი მოქმედების აქტივაცია მხოლოდ ანტისხეულებისაგან მიღებული სიგნალის შედეგად ხდება. ამ სიგნალის მიღების შედეგად, წარმოიქმნება უცხო უჯრედებისათვის მომაკვდინებელი ნაერთები (ციტოკინები).
ნახ. 3: უჯრედის სხვადასხვა ნაწილის ურთიერთგანლაგების გამორკვევა „მანათობელი ანტისხეულების“ დახმარებით: უჯრედის მატრიქსი („ჩონჩხი“, მწვანე).ნუკლეინის მჟავები (დნმ, ლურჯი), „ბოჭკოები“, რომლებიც უჯრედის გაყოფის დროს გენეტიკური მასალის (დნმ-ის) ზუსტად გადანაწილებას ახდენენ (წითელი).
ზოგადად, კომპლემენტის სისტემა: 1) აძლიერებს ანტიგენების შთანთქმას ფაგოციტების მიერ; 2) იზიდავს „შთანმთქმელ უჯრედებს“ (მაკროფაგებს და ნეიტროფილებს) პათოგენებისაკენ; 3) შლის პათოგენური უჯრედების გარსს; 4) ახდენს ანტიგენების შემცველი ნაერთების ერთად დაგროვებას (რაც მათ განადგურებას აადვილებს); 5) ურთიერთქმედებს ვირუსებთან და მათ გამრავლებას აფერხებს. და ბოლოს, კომპლემენტის სისტემის ზოგიერთი ნაწილი შედის ე.წ. „მემბრანების დაშლის კომპლექსში“, რომლის შედეგადაც ანტისხეულებს ძალუძთ მათთან მიერთებული ბაქტერიების სწრაფი განადგურება მათი დაშლის გზით.
იმუნოგლობულინების მრავალფეროვნება: თითქმის ყველა მიკროორგანიზმის შეჭრა ადამიანის სხეულში იწვევს იმუნურ რეაქციას. მრავალი სახის მავნე ორგანიზმის გამოცნობა და გაუვნებლება ანტისხეულების დიდ მრავალფეროვნებას მოითხოვს, რაც, თავის მხრივ, მათი შემადგენელი ცილების აგებულების დიდი მრავალფეროვნებითაა გაპირობებული. სავარაუდოა, რომ ადამიანის ორგანიზმი 10 მილიარდ სხვადასხვა ანტისხეულს წარმოქმნის, და მათგან თითოეულს სპეციფიური ანტიგენის ამოცნობა და მიერთება შეუძლია (ამ სტატიის დასაწყისში განხილული „გასაღებისა და ბოქლომის“ პრინციპით).
V(D)J რეკომბინაცია. იმუნოგლობულინების მიერ ფრაგმენტების მიმოცვლა, რისი შედეგიც ახალი სახის „ცვლადი“ ფრაგმენტები წარმოიქმნება, V(D)J რეკომბინაციის სახელითაა ცნობილი. ანტისხეულების როგორც მძიმე, ასევე მსუბუქი ჯაჭვები გენების რამდენიმე ფრაგმენტითაა კოდირებული. ესენია ცვლადი (Variable, V), მრავალფეროვანი (Diversity, D), და შემაერთებელი (Joining, J) ფრაგმენტები. მძიმე ჯაჭვები სამივე ფრაგმენტს შეიცავენ, ხოლო მსუბუქი ჯაჭვები - მხოლოდ V და J ფრაგმენტებს. ადამიანის გენებში ამ ფრაგმენტების მრავალი ასლი არსებობს, და სწორედ ამ ასლების შემთხვევითი მიერთება განაპირობებს ანტისხეულების მრავალფეროვნებას: მაგალითად, მძიმე ჯაჭვის „აწყობისას“ გამოიყენება V, D, და J ფრაგმენტების თითო ასლი, და რადგან არჩევანი საკმაოდ დიდია, წარმოქმნილი ანტისხეულების მრავალფეროვნებაც მნიშვნელოვანია.
ანტისხეულების გამოყენება დიაგნოსტიკისა და მკურნალობისათვის. დიაგნოსტური მიზნებისათვის ის გარემოება გამოიყენება, რომ ორგანიზმი უცხო სხეულის აღმოჩენის პასუხად ანტისხეულებს გამოიმუშავებს. რამდენიმე ვირუსული დაავადების დიაგნოსტიკის დროს ამ ვირუსების წინააღმდეგ გამომუშავებული ანტისხეულების რაოდენობა განისაზღვრება: თუ ანტისხეულები მცირე რაოდენობითაა ან საერთოდ არ არის, დაავადების არსებობა გამოირიცხება. გარკვეული სახის იმუნოგლობულინების რაოდენობის ზრდა ზოგიერთ შემთხვევაში ღვიძლის დაზიანების უტყუარი მაჩვენებელია, მაშინაც კი, როდესაც ამ დაზიანების მიზეზი ჯერ კიდევ არაა დადგენილი: მაგალითად, IgA-ს დიდი რაოდენობა ალკოჰოლიზმით გამოწვეული ციროზის მიმანიშნებელია, IgM-ის - ვირუსული ჰეპატიტისა, ხოლო IgG - აუტოიმუნური ჰეპატიტის დროს. აუტოიმუნური დაავადებები ასევე ამოიცნობა იმ ანტისხეულების დაგროვებით, რომლებიც საკუთარი ორგანიზმის სასარგებლო უჯრედებს (მაგ., სისხლის წითელ უჯრედებს) უერთდებიან, როგორც ეს ჰემოლიზური ანემიის დროს ხდება. ზოგიერთი იმუნური დაავადება, მაგალითად X-ქრომოსომასთან შეჭიდული აგამაგლობულინემია და ჰიპოგამაგლობულინემია, ანტისხეულების არარსებობით ან მათი რაოდენობის უკიდურესი სიმცირითაა გამოწვეული.
ანტისხეულების გამოყენება ბიოლოგიური ექსპერიმენტებისათვის. ამ მიზნებისათვის, ანტისხეულები გამოიყენება სხვადასხვა უჯრედსშიდა კომპონენტის (ცილების, ნუკლეინის მჟავების, ცხიმების) განლაგების გამოსარკვევად. ანტისხეულებს ამ კომპონენტების მიმართ რომელიმე ცხოველის ორგანიზმში გამოიმუშავებენ (თაგვი, ვირთხა, თხა, ცხვარი, ცხენი), და ცხოველის სისხლიდან ანტისხეულებს გამოყოფენ. ასეთ ანტისხეულებზე ქიმიური რეაქციის მეშვეობით ამაგრებენ „მანათობელ“ მოლეკულებს. როდესაც უჯრედში არსებულ ცილისა თუ ცხიმის მოლეკულას ასეთი ანტისხეული მიუერთდება, მისი მდებარეობის გარკვევა საკმაოდ ადვილია მიკროსკოპის დახმარებით - საკმარისია „მანათობელი წერტილების“ აღმოჩენა უჯრედის შიგთავსში.
დასკვნა: ანტისხეულების დაცვით სისტემას უდიდესი მნიშვნელობა აქვს ორგანიზმისათვის. მართლაც გასაოცარია ის ფაქტი, რომ ამ სისტემას ძალუძს ნებისმიერი „მომხდურის“ ამოცნობა და, თუ მისი სრული გაუვნებლება არა (როგორც უმეტეს შემთხვევაში ხდება), მნიშვნელოვნად შეფერხება მაინც. მოლეკულური ბიოლოგიის და გენური ინჟინერიის განვითარებასთან ერთად, ანტისხეულებმა ახალი მნიშვნელობა შეიძინეს როგორც გამოყენებითი და დიაგნოსტური მიზნებისათვის, ასევე ფუნდამენტური კვლევებისათვის.
გელა თევზაძე.
24 საათი 26.06.11
|
|
12 სქესის განსაზღვრის მექანიზმები მცენარეებსა და ცხოველებში. |
▲ზევით დაბრუნება |
ბავშვის დაბადების წინ, უმთავრესი საკითხის - მისი ჯანმრთელობის მდგომარეობის - გამორკვევის გარდა, მშობლებისა და ნათესავებისათვის ზოგიერთ შემთხვევაში არანაკლები მნიშვნელობა ენიჭება ამ ბავშვის სქესის გაგებასაც. ბავშვის სქესის გაგება ჩანასახის განვითარების მხოლოდ გარკვეულ სტადიაზე შეიძლება, თუმცა სქესის განსაზღვრა გაცილებით ადრე ხდება - განაყოფიერებული კვერცხუჯრედის წარმოქმნისთანავე. ეს ერთი უჯრედი შეიცავს სრულ გენეტიკურ ინფორმაციას, რომელიც ორგანიზმის ზრდისა და განვითარებისთვისაა აუცილებელი. ამ ინფორმაციის ერთ-ერთი მნიშვნელოვანი ნაწილია ორგანიზმის სქესი, რომლის განსაზღვრის მექანიზმები ადამიანსა და სხვა მრავალუჯრედიან ორგანიზმებში ამ სტატიის თემაა.
აუტოსომები და სქესობრივი ქრომოსომები. როგორც ცნობილია, ყველა ორგანიზმის გენეტიკური ინფორმაცია დეზოქსირიბონუკლეინის მჟავაში (დნმ-ში, დნ მჟავაში) არის კოდირებული. ეს ინფორმაცია და, მაშასადამე, დნმ მოთავსებულია ერთ ქრომოსომაში (ბაქტერიების შემთხვევაში) ან რამდენიმე ქრომოსომაშია გადანაწილებული (სოკოებში, მცენარეებსა და ცხოველებში). წინამდებარე სტატია ისეთი ორგანიზმების მაგალითებს განიხილავს, რომლებსაც სქესობრივი გამრავლების უნარი აქვთ, ანუ ზემოთჩამოთვლილ ორგანიზმებიდან ყველას, ბაქტერიების გარდა. როგორც უკვე აღვნიშნეთ, ამ ორგანიზმებს რამდენიმე ქრომოსომა აქვთ (დაინტერესებულთათვის - მრავალი ორგანიზმის ქრომოსომების რაოდენობა ამ ბმულზეა დასახელებული:
http://en.wikipedia.org/wiki/List_of_organisms_by_chromosome_count). ორგანიზმების განვითარება ევოლუციური თვალსაზრისით და მათი სირთულე ქრომოსომების რაოდენობასთან არავითარ კავშირში არაა: მაგალითად, ადამიანს 46 ქრომოსომა აქვს, ვირთხას - 42, ბამბას (რომელიც უმაღლეს მცენარეებს მიეკუთვნება) - 52, ერთ-ერთ გვიმრას (უმდაბლესი მცენარეების წარმომადგენელს) - ~1200, ძაღლს - 78, საფუარ სოკოს - 32, ხილის ბუზს (დროზოფილას) კი - 8.
რადგან ეს ორგანიზმები სქესობრივი გზით მრავლდებიან, ყველა მათგანი ქრომოსომების თანაბარ რაოდენობას იღებს ორივე მშობლისაგან. ყოველი ქრომოსომა ორი ასლითაა წარმოდგენილი, რომლებისგანაც ერთი დედისგანაა მიღებული, მეორე კი - მამისაგან. ამ ქრომოსომებიდან უმეტესობა ერთნაირია როგორც მდედრობითი, ასევე მამრობითი სქესის წარმომადგენლებში, და ასეთ ქრომოსომებს აუტოსომები ეწოდებათ. ის ქრომოსომები კი, რომელთა არსებობა ან მათი რაოდენობა განსხვავდება მდედრობითსა და მამრობით ორგანიზმებში, სქესობრივი ქრომოსომების სახელწოდებითაა ცნობილი. მაგალითად, დროზოფილას 8 ქრომოსომიდან 6 აუტოსომაა, 2 კი - სქესობრივი ქრომოსომა. სხვაგვარად რომ ვთქვათ, დროზოფილას 3 წყვილი აუტოსომა აქვს და 1 წყვილი სქესობრივი ქრომოსომა. თითოეულ წყვილში ერთი ქრომოსომა დედისგანაა მიღებული, მეორე კი - მამისაგან.
სქესის განსაზღვრა მცენარეებში. სქესის მიხედვით მცენარეები იყოფა ერთსახლიან და ორსახლიან მცენარეებად. ერთსახლიან მცენარეებში ცალკეულ ინდივიდები როგორც მდედრობითი, ასევე მამრობითი სქესის ყვავილების შეიცავენ, მაშინ როდესაც ცალკეულ ორსახლიან მცენარეზე ან მხოლოდ მდედრობითი, ან მხოლოდ მამრობითი ყვავილები იზრდება.
სიმინდი ერთსახლიანი მცენარის შესანიშნავი მაგალითია. მდედრობითი სქესის ყვავილებიდან სიმინდის ტარო წარმოიქმნება, მამრობითი სქესის ყვავილები კი მცენარის ღეროს წვერზე არიან მოქცეული და ე.წ. საგველას ქმნიან. მცენარის განვითარების დროს ყვავილებში ორივე სახის სასქესო ორგანოები წარმოიქმნება, მაგრამ შემდგომ ერთ-ერთი სქესის ორგანოები აღარ ვითარდება. ასეთი განვითარება 6 სხვადასხვა გენითაა განსაზღვრული, რომლებსაც Tassellseed („საგველას თესლის“, შემოკლებით Ts) ეწოდებათ. მუტაციები ამ გენებში იწვევენ სიმინდის მდედრობითი ან მამრობითი ყვავილების არასწორ განვითარებას, მაგალითად: ts1 და ts2 მუტაციების შედეგად საგველაში მდედრობითი სქესის ყვავილების განვითარების შეწყვეტა აღარ ხდება; ts5 მუტაცია ასევე იწვევს მდედრობითი სქესის ყვავილების წარმოქმნას, მაგრამ მხოლოდ საგველას ქვედა ნაწილში, მაშინ როდესაც ზედა ნაწილში მამრობითი სქესის ყვავილები ნორმალურად ვითარდებიან. ts4-ის და ts6-ის შედეგად საგველაში მდედრობითი სქესის, ტაროში კი მამრობითი სქესის ყვავილები ვითარდება.
ორსახლიანი მცენარეების მაგალითზე კი უკვე შესაძლოა სქესის განსაზღვრის სხვადასხვა მექანიზმების აღწერა: ერთ შემთხვევაში, მცენარეების სქესი განისაზღვრება ერთ-ერთი სქესობრივი ქრომოსომის არსებობითა თუ არარსებობით, მეორეში კი - სქესობრივი ქრომოსომებისა და აუტოსომების რაოდენობის შეფარდებით. დაწვრილებით განვიხილოთ ეს მექანიზმები:
სქესის განსაზღვრის აქტიური Y(იგრეკ) ქრომოსომის სისტემა: ზოგიერთ ორსახლიან მცენარეში (მაგალითად, კანაფში, აგრეთვე მიხაკისებრთა ოჯახის ერთ-ერთ წარმომადგენელში) მდედრობითი და მამრობითი მცენარეები ერთმანეთისაგან განსხვავდება სქესობრივი ქრომოსომებით: მამრობითი სქესის მცენარეებში ერთი X (იქს) და ერთი Y(იგრეკ) ქრომოსომაა, მდედრობითში კი ორი X. Y ქრომოსომა შეიცავს გენებს, რომლებიც მამრობით სქესს განსაზღვრავენ, ისევე როგორც გენებს, რომლებიც მდედრობითი სქესის განვითარებას თრგუნავენ. X ქრომოსომა არანაკლებ მნიშვნელოვანია როგორც მდედრობითი, ასევე მამრობითი სქესის მცენარეების არსებობისათვის: ქრომოსომების „ამოცლის“ მეთოდით შეიძლება ისეთი უჯრედების მიღება, რომლებიც ერთ X-ს შეიცავენ, მაგრამ უჯრედი, რომელიც თუნდაც ერთ X-ს არ შეიცავს, არასიცოცხლისუნარიანი აღმოჩნდა. ამ სისტემაში Y ქრომოსომა სქესის განსაზღვრის მთავარი ფაქტორია: ერთი Y ქრომოსომისა და სამი X ქრომოსომის შემცველი უჯრედიდან მაინც მამრობითი სქესის მცენარე ვითარდება. ეს ფაქტი, ისევე როგორც ის გარემოება, რომ ჰორმონების დამატება არ იწვევს სქესის ცვლილებას, მიგვანიშნებს, რომ ამ მცენარეებში Y ქრომოსომას გადამწყვეტი მნიშვნელობა აქვს სქესის განსაზღვრაში.
სატაცურის (ლათ. Asparagus, რუს. спаржа) შემთხვევაში, სადაც XY ქრომოსომებს ასევე მამრობითი სქესის მცენარეები შეიცავენ, იშვიათად ხდება ისეთი მამრობითი მცენარეების წარმოქმნა, რომლის ყვავილები ორივე სქესის ორგანოებს შეიცავენ. ასეთი ყვავილების თვითდამტვერვისას, შესაძლებელია YY ქრომოსომების შემცველი მამრობითი სქესის მცენარეების მიღება. YY მამრობითი და XX მდედრობითი მცენარეების შეჯვარებით იღებენ XY გენოტიპის (გენოტიპი გენების ერთობლიობას ნიშნავს) შთამომავლობას, სადაც ყველა მცენარე მამრობითი სქესისაა. რადგან მამრობითი სქესის სატაცური უფრო დიდი ზომისაა, ასეთ შეჯვარებას კომერციული მნიშვნელობაც აქვს.
სქესის განსაზღვრის ბალანსის სისტემა: X-ისა და აუტოსომების რაოდენობის შეფარდება.
მჟაუნას (ლათ. Rumex, რუს. щавель) გვარის მცენარეებში სქესის განსაზღვრის სხვა სისტემა მოქმედებს: აქ სქესი დამოკიდებულია X ქრომოსომებისა და აუტოსომების რაოდენობის შეფარდებაზე. უნდა დავაზუსტოთ, რომ აუტოსომები განიხილება, როგორც ერთობლიობა: დიპლოიდურ მცენარეში, რომელიც ყველა ქრომოსომის ორ ასლს შეიცავს (მათგან ერთი დედისაგანაა მიღებული, მეორე კი - მამისაგან, იხ. ზემოთ), აუტოსომების რაოდენობა პირობითად 2-თ განისაზღვრება. თუ მცენარეში ორი X ქრომოსომაა, მაშინ X ქრომოსომებისა და აუტოსომების რაოდენობის შეფარდება (ე.წ. ბალანსი) 1.0-ია და ასეთი მცენარე მდედრობითი სქესისაა. მამრობითი სქესის მცენარეებში ერთი X (და ერთი Y) ქრომოსომაა, და ამიტომ X-ქრომოსომებისა და აუტოსომების რაოდენობის შეფარდება 0.5(ანუ 1/2)-ია. თუ ორგანიზმი პოლიპლოიდურია (ანუ აუტოსომების ერთობლიობა არა 2, როგორც დიპლოიდში, არამედ 3 ან მეტია), ასეთი ორგანიზმიც მამრობითი სქესისა იქნება. ამრიგად, თუ X ქრომოსომებისა და აუტოსომების რაოდენობის შეფარდება 1.0 ან 1.0-ზე მეტია, ეს მდედრობით სქესს განსაზღვრავს, 0.5 ან 0.5 ნაკლები - მამრობითს. ის მცენარეები, რომლებშიც შეფარდება 0.5-სა და 1.0-ს შორისაა (მაგალითად, 0.66 - ორი X ქრომოსომა და აუტოსომების სამი ერთობლიობა), ჰერმაფროდიტიზმს ან შერეულ სქესს ავლენენ.
მჟაუნას შემთხვევა იმითაცაა საინტერესო, რომ მდედრობითი სქესის მცენარეებში ორი X ქრომოსომაა, მაშინ როდესაც მამრობითი სქესი ერთ X და ორ Y ქრომოსომას შეიცავს. ამის გამო, რადგან აუტოსომების რიცხვი ორივე სქესში თანაბარია, მდედრობითი სქესის მცენარეებში 14 ქრომოსომაა, მამრობითისაში კი 15. მჟაუნას Y ქრომოსომები აუცილებელია განაყოფიერებისათვის, მაგრამ არა მამრობითი სქესის ყვავილების წარმოქმნასა და განვითარებისათვის, ისევე როგორც მდედრობითი სქესის ყვავილების განვითარების შესაჩერებლად. ეს გარემოება კიდევ ერთხელ ასაბუთებს, რომ მჟაუნას Y ქრომოსომა მნიშვნელოვანია გამრავლებისათვის, მაგრამ არა სქესის განსაზღვრისათვის.
სქესის განსაზღვრა დროზოფილაში: განხილული მაგალითების საფუძველზე უკვე ჩანს, რომ სქესის განსაზღვრის რამდენიმე მექანიზმი არსებობს. როგორც აღვნიშნეთ, ეს მექანიზმები არა მარტო მცენარეებში, არამედ სხვა ორგანიზმებშიცაა გავრცელებული. ხილის ბუზი დროზოფილა, რომელიც 8 ქრომოსომას შეიცავს (6 აუტოსომა და 2 სქესობრივი ქრომოსომა), სქესის განსაზღვრის გენეტიკური მექანიზმის არანაკლებ კარგი მაგალითია:
ნახ. 1: მდედრობითი (მარცხნივ) და მამრობითი (მარჯვნივ) სქესის დროზოფილა.
მდედრობითი და მამრობითი სქესის დროზოფილები გარეგნულად ერთმანეთისაგან მნიშვნელოვნად განსხვავდებიან (ეს ფაქტი ე.წ. სქესობრივ დიმორფიზმს გამოხატავს). ჩვეულებრივ, მდედრობითი სქესის დროზოფილებს ორი X ქრომოსომა აქვთ, მამრობითი სქესისა კი - ერთი X და ერთი Y. ორივე სქესის წარმომადგენლებს აუტოსომების ორი ერთობლიობა აქვთ (3 წყვილი, ყოველ წყვილში თითოეული ქრომოსომა სხვადასხვა მშობლისგანაა მიღებული). ისევე როგორც ერთ-ერთ განხილულ შემთხვევაში, სქესი დროზოფილაში განისაზღვრება X ქრომოსომებისა და აუტოსომების ერთობლიობის რაოდენობის შეფარდებით (აუტოსომების რაოდენობა A-თი აღინიშნება). X:A შეფარდება, როგორც ჯერ კიდევ მეოცე საუკუნის დასაწყისში დადგინდა, მდედრობითი სქესის შემთხვევაში 1.0 ან 1.0-ზე მეტია, მამრობით სქესში კი 0.5 ან 0.5-ზე ნაკლები. Y ქრომოსომა მამრობითი სქესის ორგანიზმებში სპერმის ნორმალური განვითარებისთვისაა აუცილებელი, მაგრამ მათი სქესობრივი ნიშან-თვისებებისათვის წარმოქმნისათვის საჭირო არაა.
როგორ ხდება X:A შეფარდების ინფორმაციის გაგება უჯრედების მიერ და რა მექანიზმით ხდება ამ ინფორმაციის გამოხატვა დროზოფილის ორგანიზმში? X:A შეფარდებით განისაზღვრება უმთავრესი რეგულატორული გენის, Sex-lethal (Sxl)-ის ფუნქციონალური მდგომარეობა. მდედრობითი სქესის დროზოფილებში Sxl გააქტივებულია, და მისი აქტივობის შედეგად ხდება კიდევ ოთხი გენის აქტივირება. სხვადასხვა მუტაციების ანალიზის საფუძველზე განისაზღვრა, რომ Sxl-ზე კიდევ ბევრი გენის ფუნქციაა დამოკიდებული, რომლებიც ე.წ. „მდედრობითი სიგნალის“ გადაცემას უზრუნველყოფენ, რაც ორგანიზმის განვითარების განმსაზღვრელი ცილების სინთეზში გამოიხატება. მამრობითი სქესის დროზოფილაში Sxl გენი აქტიური არაა, და შესაბამისად არც Sxl-ის მიერ რეგულირებადი გენების ფუნქციაა გამოხატული. ამის შედეგად კი, თავის მხრივ, იმ გენების ექსპრესია (ანუ მათში კოდირებული ინფორმაციის ამოკითხვა) ხდება, რომლებიც მამრობითი სქესის ორგანიზმის განვითარებისთვისაა აუცილებელი.
საინტერესოა Sxl-ისა და მასზე დამოკიდებული გენების მუტაციების შესწავლაც, რადგან ასეთი ანალიზით შესაძლოა გამოაშკარავდეს არა მარტო გენების ზოგადი ფუნქცია, არამედ მათი (და, შესაბამისად, მათ მიერ კოდირებული ცილების) ადგილი სქესის განმსაზღვრელი გენების სისტემაში. მაგალითად, მუტაცია თვით Sxl გენში იწვევს მამრობითი სქესის ფენოტიპის (გარეგნული შესახედაობის) დროზოფილების განვითარებას, თუნდაც მათი უჯრედები ორ X ქრომოსომას შეიცავდეს. ამავე გენის სხვაგვარი მუტაციის შედეგად XY ორგანიზმებში უკვე მდედრობითი სქესის ფენოტიპი წარმოიქმნება. თუმცა ასეთი ორგანიზმები ორივე შემთხვევაში ზრდასრულ ასაკამდე იღუპებიან. Sxl-ის მიერ აქტივირებული გენების მუტაციების შედეგი კი „შუალედური სქესის“ წარმოქმნაა როგორც XX, ასევე XY ორგანიზმებში.
გენეტიკური კვლევების შედეგად დადგინდა, Sxl-ის აქტივაციისათვის კიდევ ორი გენია აუცილებელი - da და dk. მუტაციები da (ან dk) გენში მომაკვდინებელია ორგანიზმისათვის, მაგრამ თუ იგივე ინდივიდი Sxl მუტაციასაც შეიცავს, მაშინ ის დაღუპვას გადაურჩება. ანუ, სხვაგვარად რომ ვთქვათ, ცალკ-ცალკე მუტაციები Sxl-სა და da (dk)-ში დამღუპველია, მაგრამ ორივე ერთად ერთმანეთის ეფექტს ანეიტრალებს და სიცოცხლის შენარჩუნების შესაძლებლობას იძლევა. ამ ეფექტის ერთი შესაძლო ახსნა შემდეგშია: ცნობილია, რომ მუტაციები da/dk გენებში მომაკვდინებელია მხოლოდ მდედრობითი სქესისათვის, რომელიც, როგორც აღვნიშნეთ, ორ X ქრომოსომას შეიცავს. da გენი ამ ორი ქრომოსომის გადაჭარბებულ აქტივაციას (რაც ორგანიზმისათვის საზიანოა) აბრკოლებს, ამ გენში მუტაციის შეყვანის შედეგად კი ეს დაბრკოლება აღარ ხდება. სამაგიეროდ, Sxl-ის მუტაციის დამატება (რომლის შედეგადაც X-ქრომოსომაზე მოთავსებული გენები „გამოირთვება“), ამ პრობლემას მეტ-ნაკლებად აბათილებს.
ამრიგად, სქესის განსაზღვრა დროზოფილაში შესაძლოა შემდეგი დებულებებით შეჯამდეს:
მდედრობითი სქესი: 1) როცა X:A=1, რამდენიმე სხვა ფაქტორთან ერთად ეს იწვევს Sxl გენის „ჩართვას“; 2) Sxl-ის „ჩართვის“ შედეგად, აქტიურდება როგორც თვით Sxl გენი (ე.წ. აუტორეგულაცია), ასევე Sxl-ზე დამოკიდებული რამდენიმე სხვადასხვა გენიც; 3) ამ გენების თანმიმდევრული მოქმედებით ხდება იმ გენების „გამორთვა“, რომლებიც მამრობითი სქესის ორგანიზმის განვითარებას აპირობებენ.
მამრობითი სქესი: 1) როცა X:A=0.5, Sxl გენი „გამორთულია“; 2) შესაბამისად, ინაქტივირებულია ანუ ასევე „გამორთული „Sxl-ზე დამოკიდებული გენები; 3) ერთ-ერთი გენი, რომელიც მდედრობით ორგანიზმში Sxl-ის გავლენით „გამორთულია“,Sxl-ის „არყოფნის“ პირობებში აქტიურდება და მდედრობითი სქესის ორგანიზმის განმსაზღვრელი გენების „გამორთვას“ იწვევს, რის შედეგადაც მამრობითი სქესის დროზოფილა ვითარდება.
როგორც ზემოთქმულიდან ჩანს, დროზოფილაში სქესის განსაზღვრა X:A ბალანსის საფუძველზე ხდება, მაგრამ ეს ბალანსი მხოლოდ საწყისი რგოლია გენების იმ ჯაჭვში, რომლებიც „ჩართვისა“ თუ „გამორთვის“ საფუძველზე მიიღება ამათუიმ სქესის ორგანიზმი.
და ბოლოს, როგორ ხდება სქესის განსაზღვრა ძუძუმწოვრებში და, კერძოდ, ადამიანში? ამ შემთხვევაში, ერთი შეხედვით, სისტემა საკმაოდ მარტივია: Y ქრომოსომის არსებობა განაყოფიერებულ კვერცხუჯრედში საკმარისია იმისათვის, რომ მამრობითი სქესის ორგანიზმი განვითარდეს. როგორც თაგვებში, ასევე ადამიანებში ერთი X ქრომოსომის არსებობა მდედრობითი სქესის ორგანიზმის განვითარებას იწვევს (გავიხსენოთ, რომ დროზოფილას შემთხვევაში ერთი X ქრომოსომა მამრობითი სქესის განმსაზღვრელია: X:A=1:2=0.5). და პირიქით, თუ ორგანიზმში თუნდაც ერთი Y ქრომოსომაა, იგი აუცილებლად მამრობითი სქესის იქნება, თუნდაც მისი უჯრედები ორ ან მეტ X ქრომოსომას შეიცავდეს.
მაგრამ რითია გაპირობებული Y ქრომოსომის ასეთი კრიტიკული მნიშვნელობა? როგორც ადვილი წარმოსადგენია, ეს განისაზღვრება ამ ქრომოსომაზე მოთავსებული გენებით და მათ მიერ კოდირებული ფუნქციებით. ერთ-ერთი გენის სახელწოდებაა TDF (testis-determining factor, სათესლეების განმსაზღვრელი ფაქტორი), რომელსაც მოგვიანებით SRY (sex-determining region, Y chromosome) ეწოდა. თაგვებზე ჩატარებული ცდების შედეგად გაირკვა, რომ ამ გენის „ჩართვა“ ხდება სასქესო ორგანოების განვითარების ადრეულ სტადიაზე, როდესაც სათესლეების განვითარება იწყება. კიდევ ერთი, არანაკლებ საინტერესო ფაქტი: როდესაც SRY გენი თაგვის ახალგანაყოფიერებული მდედრობითი სქესის უჯრედებში მოათავსეს, ამ უჯრედებიდან მამრობითი სქესის ცხოველები განვითარდნენ.
შემდგომმა კვლევებმა აჩვენა, რომ SRY გენი ასევე აუცილებელია სათესლეების განვითარებისათვის ადამიანის ორგანიზმში. SRY-ს მუტაციები 25%25-ის სიხშირით გვხვდება ადამიანებში, რომლებიც X და Y ქრომოსომებს შეიცავენ, მაგრამ ფენოტიპურად ქალები არიან. და პირიქით, ადამიანები, რომლებსაც მამაკაცივით გამოიყურებიან, მაგრამ ორ X ქრომოსომას (მდედრობითი სქესის გენოტიპს) შეიცავენ, ხშირ შემთხვევაში Y ქრომოსომის მცირე ზომის ფრაგმენტებსაც ატარებენ.
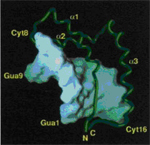
ნახ. 2: დნმ (მწვანე) ფორმას იცვლის - „იღუნება“ SRY ცილასთან (ცისფერი) ურთიერთქმედების შედეგად.
SRY გენის და მისი პროდუქტის (SRY ცილის) მოქმედების მექანიზმი ბოლომდე დადგენილი არაა, თუმცა რამდენიმე საინტერესო კანონზომიერება უკვე აღმოჩენილია: SRY ცილა შეიცავს დნმ-ის ბმის უნარის მქონე ფრაგმენტს, რაც იმის მიმანიშნებელია, რომ ეს ცილა სხვა გენების რეგულატორულ ფრაგმენტებს ებმის. ეს კი, თავის მხრივ, ნიშნავს, რომ ასეთი ბმა გავლენას ახდენს ამ გენებში კოდირებული ინფორმაციის ამოკითხვაზე. რამდენიმე წლის წინ აღმოჩნდა, რომ SRY ცილასთან ბმის შედეგად დნმ-ის კონფიგურაცია მნიშვნელოვნად იცვლება - იგი მკვეთრად იხრება და 70-80-იან კუთხეს ქმნის. ასევე დადგენილია SRY ცილისა და დნმ-ის ურთიერთქმედების შედეგად წარმოქმნილი კომპლექსის სამგანზომილებიანი სტრუქტურა.
რაც განსაკუთრებით საინტერესოა, SRY გენის მუტაციების შესწავლის საფუძველზე გამოირკვა, რომ ის მუტაციები, რომლებიც სქესის ცვლილებას იწვევს, სწორედ იმ მონაკვეთში იყო განლაგებული, რომელშიც დნმ-ის ბმის უნარის მქონე ფრაგმენტია კოდირებული: ეს მუტაციები ან SRY ცილის დნმ-თან ბმის უნარს ამცირებენ, ან ამ ბმის შედეგად დნმ-ის „მოხრის კუთხეს“ ცვლიან. ამ აღმოჩენის უდაო მნიშვნელობის მიუხედავად, ჯერ გაურკვეველია, რომელი გენების რეგულატორულ ფრაგმენტებთან ურთიერთქმედებს SRY ცილა.
ამ გენების იდენტიფიკაციისას ერთ-ერთი შესაძლო კანდიდატია AMH, რომლის მიერ კოდირებული ცილა სათესლეებში გროვდება მათი ფორმირების შემდეგ (გავიხსენოთ, რომ SRY ცილა სათესლეების განვითარების ადრეულ სტადიაზე წარმოიქმნება). რადგან SRY უფრო ადრე წარმოიქმნება, ვიდრე AMH, შესაძლოა, რომ ამ უკანასკნელის რეგულაცია SRY ცილით ხდება. მეორე მხრივ, AMH-ის კიდევ ერთი შესაძლო რეგულატორია SF-1 გენი, რომელიც არა მარტო სასქესო ორგანოების განვითარებას, არამედ თირკმელების ფორმირებასა და სტეროიდების სინთეზსაც აკონტროლებს. ამრიგად, SF-1 ჭეშმარიტად გლობალური რეგულატორის შესანიშნავი მაგალითია, და ორგანიზმის მრავალი ფუნქციის ურთიერთკავშირის კარგი მაჩვენებელია. რამდენიმე წლის წინ აღმოაჩინეს კიდევ ერთი გენი (Sox9), რომელიც, SRY-ს დარად, დნმ-ის ბმის უნარიოს მქონე ფრაგმენტს შეიცავს.
ნახ. 3: სქესის განსაზღვრა ადამიანში: ნაჩვენებია თითოეული მშობლისაგან მიღებული სქესობრივი ქრომოსომები.
დასკვნა: სავარაუდოა, რომ ადამიანის სქესის განმსაზღვრელი გენების სია ჯერ მეტად არასრულია. როგორც აღვნიშნეთ, მუტაციები SRY-ში სქესის ცვლილების მხოლოდ 25%25-ს განსაზღვრავს. არსებობს რამდენიმე გენი, რომელიც X ქრომოსომაზეა განლაგებული და სქესის განსაზღვრაში საკმაოდ მნიშვნელოვანი როლი აქვს. ზოგადად კი, ადამიანის სქესის განსაზღვრა მხოლოდ გენეტიკური ფაქტორების (ქრომოსომების და მათზე განლაგებული გენების) გავლენით ხდება: დედისაგან, რომელსაც ორი X ქრომოსომა აქვს, ბავშვი ყოველთვის X სქესობრივ ქრომოსომას იღებს (22 აუტოსომასთან ერთად); ამიტომ ბავშვის სქესი მხოლოდ იმით განისაზღვრება, თუ რომელ სქესობრივ ქრომოსომას მიიღებს მამისაგან (რომელსაც როგორც X, ასევე Y ქრომოსომები აქვს): X ქრომოსომის მიღება მდედრობით სქესს განაპირობებს, Y-ისა კი - მამრობითს.
გელა თევზაძე.
24 საათი 19.07.11
|
|
13 გრეგორ მენდელი: თანამედროვე გენეტიკის დამფუძნებელი და ერთ-ერთი უდიდესი მეცნიერი კაცობრიობის ისტორიაში |
▲ზევით დაბრუნება |
20 ივლისი გრეგორ მენდელის დაბადების დღეა.

ნახ. 1: გრეგორ იოჰან მენდელი (1822-1884).
მენდელის დამსახურება კაცობრიობის წინაშე - და მის მიერ აღმოჩენილი კანონების გავლენა თანამედროვეობაზე - არანაკლებია, ვიდრე ნიუტონისა და დარვინის თეორიებისა. ამ გარემოების მიუხედავად, ნიუტონისა და დარვინისაგან განსხვავებით, მენდელის გარდაცვალების დღეს (1884 წლის 6 იანვარი) მისი წვლილი მეცნიერებაში ჯერ მეტად შორს იყო საყოველთაო აღიარებისაგან. ამ სტატიის მიზანია მკითხველს მოუთხროს როგორც მენდელის სამეცნიერო ღვაწლის (რაც მეტ-ნაკლებად ცნობილია), ასევე მისი ცხოვრების შესახებ. სავარაუდოა, რომ მენდელის ცხოვრების დეტალების გაცნობა არც თუ ისე უინტერესო იქნება, რადგან ჭეშმარიტად იშვიათია ადამიანი, რომლის ცხოვრება, კარიერა და სამეცნიერო მოღვაწეობა ერთმანეთთან ასეთ სრულყოფილ ჰარმონიაში ყოფილიყოს. ამასთანავე, მენდელის პიროვნული თვისებების საფუძველზე ადვილი ასახსნელია მისი აღმოჩენებიცა და ზოგადად, მისი ბედიც. თანამედროვენი აღნიშნავენ, რომ ამ თვისებათა შორის გამოირჩეოდა მიზანსწრაფულობა, თანმიმდევრულობა, ნათელი აზროვნება, სიმართლისა და სამართლიანობის პატივისცემა, თავის კეთილისმყოფელთა ერთგულება, და რეალობის პრაქტიკული შეფასების უნარი.
იოჰან მენდელი (ეს იყო მისი სახელი ერისკაცობაში) დაიბადა 1822 წლის 20 ივლისს, სილეზიაში (მაშინ - ავსტრიის სამფლობელო, ამჟამად ჩეხეთი). მენდელის დაბადების ადგილთან დაკავშირებით საბჭოთა პროპაგანდაში საინტერესო მასალა დაგროვდა: გენეტიკის მოძულების დროს, ამ მეცნიერების (იმდროინდელი ტერმინოლოგიით - ცრუმეცნიერების) დამფუძნებლად „ავსტრიელი ბერი მენდელი“ მოიხსენიებოდა; გენეტიკის „რეაბილიტაციის“ შემდეგ, ამავე მეცნიერების მამამთავრად „ჩეხი მეცნიერი მენდელი“ აღიარეს.
მენდელის მამა, ანტონ მენდელი, მოიჯარადრე იყო, მაგრამ საკუთარი ნიჭის წყალობით ფერმის მფლობელი გახდა. მას ყველაზე მეტად ხეხილის ბაღში მუშაობა უყვარდა, რაც თავის შვილს იოჰანსაც შეასწავლა. ასე რომ მენდელის ინტერესს ბოტანიკისა და, კერძოდ, მებაღეობის მიმართ ჯერ კიდევ ადრეულ ბავშვობაში ჩაეყარა საფუძველი. მეცნიერული ნიჭი კი, შესაძლოა, მენდელმა დედის (როზინა შვირტლიხის) ოჯახიდან მიიღო: ცნობილია, რომ ბიძამისი, ანტონ შვირტლიხი, კერძო სკოლის დამფუძნებელი იყო იმავე სოფელში, სადაც იოჰან მენდელი დაიბადა. ამ სკოლაში ის ბავშვები სწავლობდნენ, რომლებიც ჯერ საკმარისად არ წამოზრდილიყვნენ იმისათვის, რომ მეზობელ სოფელში არსებულ სამრევლო სკოლაში ევლოთ. როგორც ამბობენ, თვით მენდელი ხშირად აღნიშნავდა, რომ მის წინაპრებს შორის მასწავლებლებიც იყვნენ.
ჯერ კიდევ საწყისი განათლების მიღებისას, მენდელის ერთ-ერთმა მასწავლებელმა (ავგუსტინელთა ორდენის წევრმა) შეამჩნია მისი უდაო მეცნიერული ნიჭი და ურჩია, რომ სამეცნიერო კვლევებისათვის ერთ-ერთი საუკეთესო გარემო წყნარი სამონასტრო ცხოვრება გახლდათ. 1841-1843 წლებში მენდელი სწავლობდა გიმნაზიაში, რაც ბერების განათლებისათვის აუცილებელ ეტაპად ითვლებოდა. ერთი იმდროინდელი შემთხვევა მის მადლიერების გრძნობასა და პასუხისმგებლობას კარგად წარმოაჩენს: აღმოჩნდა, რომ ოჯახის მიერ გამოყოფილი ფინანსები საკმარისი არ იყო სწავლის საფასურის გადასახდელად, და მენდელის ერთ-ერთმა დამ დარჩენილი გადასახადი თავისი მზითვიდან გაიღო. ამ დახმარების მიღება მენდელის მიზანსწრაფულობაზეც მეტყველებს - მას სურდა, აუცილებლად დაესრულებინა სწავლა. მადლიერების ნიშნად კი მენდელმა მოგვიანებით საკუთარი ხარჯებით დააფინანსა სამივე დისშვილის როგორც საწყისი, ასევე საუნივერსიტეტო განათლება.
1843 წელს მენდელი მიიღეს ქალაქ ბრუნის (ამჟამად ბრნო, ჩეხეთი) ავგუსტინელთა ორდენის მონასტერში, სადაც ის მორჩილი გახდა. საინტერესოა, რომ ერთადერთ დაბრკოლებად განიხილებოდა ჩეხური ენის უცოდინრობა (გავიხსენოთ, რომ მენდელი გერმანულენოვან ოჯახში დაიბადა), მაგრამ იქვე გამოთქვეს იმედი, რომ ამ ენის შესწავლას ნიჭიერი ახალგაზრდა დიდ დროს არ მოანდომებდა.
მომდევნო სამი წელი მენდელმა თეოლოგიის შესწავლას დაუთმო. 1846 წლის 26 დეკემბერს იგი ბერად აღიკვეცა და სახელად მიიღო „გრეგორი“. 1847 წელს მენდელი მღვდლად აკურთხეს, 1848 წლის 26 ივლისს კი ბრუნის მრევლის პასტორი გახდა. მომდევნო წლებში (1849-1850) იგი ადგილობრივ გიმნაზიასა და ტექნიკურ სკოლაში ასწავლიდა ბერძნულ ენას, ფიზიკასა და ბუნებისმეტყველებას. 1851-ში კი, როდესაც მენდელი ოცდამეათე წელში გადადგა, ბოლოს და ბოლოს აღსრულდა მისი ოცნება, მიეღო მეცნიერული ექსპერიმენტული მუშაობისათვის აუცილებელი წვრთნა: ბრუნის მონასტერმა საკუთარი ხარჯით გაგზავნა მენდელი ვენის უნივერსიტეტში, სადაც იგი ესწრებოდა ლექციებს ბოტანიკაში, ზოოლოგიაში და ფიზიკაში. მან ასევე შეისწავლა ექსპერიმენტული ფიზიკის კურსიც. ამასთანავე, იგი ცდილობდა შეევსო თავისი ცოდნა საბუნებისმეტყველო მუზეუმებისა და ლაბორატორიების დათვალიერებით, ისევე როგორც მეცნიერებთან საუბრებით. ობიექტურობა მოითხოვს იმ გარემოების მოხსენიებასაც, რომ ვენაში ყოფნისას მან ორჯერ სცადა მეცნიერული ხარისხის მოპოვება - ორჯერვე წარუმატებლად.
თუმცა მენდელმა ვენის უნივერსიტეტი ხარისხის გარეშე დატოვა, იქ მიღებული ცოდნა ექსპერიმენტულ მუშაობაში მას, როგორც ცნობილია, შემდგომში მეტად გამოადგა: მენდელის ექსპერიმენტებმა დაამტკიცა, რომ მას ბრწყინვალედ ჰქონდა ათვისებული როგორც მეცნიერული მეთოდოლოგია, ასევე რაციონალური ანალიზის პრინციპები.
ვენიდან დაბრუნებისას, მენდელმა მუშაობა დაიწყო ბუნებისმეტყველების მასწავლებლად „ბრუნის თანამედროვე სკოლაში“, სადაც მან 14 წელი გაატარა. სწორედ ამ დროის განმავლობაში ჩაატარა მენდელმა მისი - აწ სახელგანთქმული - ექსპერიმენტები ბარდის ნიშან-თვისებების მემკვიდრეობით გადაცემის შესწავლის მიზნით. ეს ექსპერიმენტები ჩატარებული იყო ბრუნის მონასტრის ბაღში.
ნახ. 2: ბარდის 6 ნიშან-თვისება (შვიდიდან, რომლებსაც მენდელი შეისწავლიდა. არაა ნაჩვენები მხოლოდ ყვავილები ღეროზე განლაგების კანონზომიერება).
იმდროინდელ ევროპელ მეცნიერებს განსაკუთრებით იტაცებდა ცოცხალი ორგანიზმების მრავალფეროვნების საკითხი, და ამ მრავალფეროვნების საფუძვლების ახსნა უმნიშვნელოვანეს და ურთულეს ინტელექტუალურ გამოწვევად იყო მიჩნეული. ზოგიერთნი ცდილობდნენ, ევოლუციის ახსნა იმ „შუალედურ ფორმებში“ მოენახათ, რომლებიც მონათესავე სახეობების შეჯვარების შედეგად წარმოიშობოდა. თუმცა ისიც ნათელი იყო, რომ ამგვარი შუალედური ფორმების შთამომავლობა ხშირად საწყისი ორგანიზმების (შუალედური ორგანიზმების მშობლების) მსგავსი ხდებოდა, ამიტომ ახლონათესაური სახეობების შეჯვარება ევოლუციის პროცესში წარმოქმნილი მრავალფეროვნების ასახსნელად არ გამოდგებოდა. ჩარლზ დარვინი კი შეეცადა, ბუნებრივი პროცესებისათვის მიესადაგებინა ხელოვნური სელექციის დარგში დაგროვილი შედეგები, და ბუნებრივი გადარჩევის თეორია შექმნა. როგორც ცნობილია, ამ თეორიამ დროის გამოცდას გაუძლო და ბუნებრივი გადარჩევა თანამედროვე მეცნიერების თვალსაზრისითაც ევოლუციის ერთ-ერთ უმთავრეს წარმმართველ ძალად ითვლება.
მენდელმაც სცადა გაერკვია, შესაძლებელია თუ არა ახალი ფორმების (და, მაშასადამე, ცვალებადობის) მიღება ორი განსხვავებული ახლონათესაური ორგანიზმის შეჯვარების შედეგად. ორწლიანი ექსპერიმენტების შემდეგ მან დაასკვნა: „ცხადია, რომ ამ გზით ბუნება ვერ მიაღწევდა პროგრესს ახალი სახეობების წარმოქმნის თვალსაზრისით. სავარაუდოა, რომ სხვა [უფრო დახვეწილი] მექანიზმები არსებობს“.
ამ პრობლემის მკაფიო ანალიზი მენდელის მიერ შესანიშნავადაა წარმოჩენილი მისი კლასიკური ნაშრომის „ექსპერიმენტები მცენარეთა ჰიბრიდებზე“ („Versuche über Pflanzen-Hybriden“, 1866 წ.) წინასიტყვაობაში: „...დღემდე, არცერთი ექსპერიმენტი არ ჩატარებულა იმგვარად, რომ შესაძლებელი ყოფილიყო ჰიბრიდების შთამომავლობის სხვადასხვა ფორმების რაოდენობის განსაზღვრა, ანდა ამ ფორმების რაოდენობების ურთიერთშეფარდების რიცხობრივი გაანგარიშება. ასეთი ექსპერიმენტის ჩატარება უაღრესად რთულია და დიდ [მეცნიერულ] თავდადებას მოითხოვს, მაგრამ ეს ერთადერთი შესაძლო გზაა, რომ გაეცეს პასუხი არცთუ მცირე მნიშვნელობის კითხვას ორგანული ფორმების ევოლუციური ისტორიის შესახებ. წინამდებარე ნაშრომი სწორედ ასეთი დეტალური ექსპერიმენტის აღწერას გთავაზობთ. იგი შეგნებულადაა დაგეგმილი, როგორც მცენარეთა მცირე ჯგუფის ანალიზი და, რვა წლის შრომის შედეგად, თითქმის დასრულებულია“. ამ ციტატაში, პრობლემის გამოკვეთის შესაშური სიცხადის გარდა, კიდევ ორი გარემოებაა შესანიშნავი: „მცენარეთა მცირე ჯგუფი“ 21 ათას (თუ წინასწარ ექსპერიმენტებს ჩავთვლით, 29 ათას) ინდივიდუალურ მცენარეს შეიცავდა. ამასთანავე, თითოეული მცენარის ანალიზი 7 სხვადასხვა ნიშან-თვისების მემკვიდრეობით გადაცემის შესწავლას ითვალისწინებდა.
არა მარტო ექსპერიმენტის გულმოდგინე დაგეგმვა მენდელის მიერ, არამედ საცდელ ობიექტად ბარდის (ლათ. Pisum sativum, ინგლ. pea, რუს. горох) არჩევანიც მის მეცნიერულ შორსმჭვრეტელობასა და დეტალებისადმი უზადო ყურადღებას წარმოაჩენს. მენდელი მოქმედებდა არა უეცარი შთაგონების ზეგავლენით, არამედ ყოველმხრივ დახვეწილი გეგმის მიხედვით. მისივე განსაზღვრებით, ბარდა არჩეული იქნა შემდეგი ფაქტების გათვალისწინებით: 1) ბარდის სხვადასხვა ჯიშები ავლენდნენ თვისებებს, რომლებიც მათთვის დამახასიათებელი იყო და ადვილად განირჩეოდა სხვა ჯიშების თვისებებისაგან; 2) ხელოვნური დამტვერვის მეთოდი ამ მცენარის შემთხვევაში შედარებით მარტივი იყო და სანდო, ადვილად განმეორებად შედეგებს იძლეოდა; 3) ჰიბრიდებისა და მათი შთამომავლობის ნაყოფიერება არ მცირდებოდა როგორც ჰიბრიდიზაციის (ანუ ჯვარედინი დამტვერვის), ასევე თვითდამტვერვის შედეგად.
მენდელის ექსპერიმენტების თავისებურება - და უპირატესობა - სხვა მკვლევარების მიერ ჩატარებულ ცდებთან შედარებით, უმთავრესად შემდეგი გარემოებით განისაზღვრება: მენდელი შეჯვარებისათვის იყენებდა ახლონათესაურ ფორმებს (ერთი სახეობის სხვადასხვა ჯიშებს), და ამ შეჯვარების შედეგად მიღებული ჰიბრიდები გამოირჩეოდნენ ისეთივე ნაყოფიერებით, როგორც მათი მშობლები. არანაკლებ მნიშვნელოვანია, რომ შვიდი სხვადასხვა ნიშან-თვისებიდან მენდელი თითოეული ნიშნის მემკვიდრეობით გადაცემას სხვა ნიშნებისაგან დამოუკიდებლად იკვლევდა.
ამ დებულებების საილუსტრაციოდ, მართებული იქნება იმ ნიშან-თვისებების ჩამოთვლა, რომლებსაც მენდელი აკვირდებოდა ბარდის მცენარეებზე. როგორც აღვნიშნეთ, სულ 7 თვისების მემკვიდრეობითი გადაცემა შეისწავლებოდა: ღეროს სიგრძე (გრძელი ან მოკლე), ყვავილების განლაგება ღეროზე (მის გასწვრივ ან კენწეროში), პარკის ფორმა (მსხვილი ან დანაოჭებული) და ფერი (ყვითელი ან მწვანე), მარცვლის ფორმა (გლუვი ან დანაოჭებული) და ფერი (ყვითელი ან მწვანე), ყვავილის ფერი (თეთრი ან იასამნისფერი). როგორც ეს ჩამონათვალი ცხადყოფს, ყველა ნიშანი ადვილი დასადგენია და მკვეთრად გამოირჩევა თავისი „მეწყვილისაგან“: თითქმის შეუძლებელია ყვითელი და მწვანე მარცვლების, თეთრი და იისფერი ყვავილების, ან თუნდაც გრძელი და მოკლე ღეროს ერთმანეთში არევა, რაც უაღრესად მნიშვნელოვანია დიდი მასშტაბის ანალიზისათვის.
თითოეული ნიშნის დამოუკიდებელი ანალიზი კი ნიშნავს, რომ თუ ყვითელი გლუვი მარცვლების მქონე მცენარეს მწვანე დანაოჭებულ მარცვლებიან მცენარეს აჯვარებდნენ, მენდელი ცალკე აღნუსხავდა მარცვლის ფერის მემკვიდრეობით გადაცემის სტატისტიკას, და ცალკე - მარცვლის ფორმის მემკვიდრეობის ამსახველ შედეგებს. ესეც მეტად მნიშვნელოვანი მეთოდი იყო, რადგან მან მენდელის ანალიზს დამატებითი დამაჯერებლობა და სიცხადე შესძინა.
თვით მენდელის კანონებს (რომლებსაც იგი, მისთვის დამახასიათებელი თავმდაბლობით, „პრინციპებად“ მოიხსენიებდა, ანუ უარყოფდა მათ აყვანას „კანონის“ რანგში და უფრო ზოგადი საფუძვლების სტატუსს ანიჭებდა) აქ მხოლოდ მოკლედ შევეხებით:
სეგრეგაციის (დათიშვის) კანონის თანახმად, ნებისმიერ ნიშან-თვისებას განსაზღვრავენ „ფაქტორები“, რომლებიც უცვლელად გადაეცემიან შთამომავლობას. ეს ფაქტორები (თანამედროვე ტერმინოლოგიით მათ „გენებს“ ვუწოდებთ) შეიძლება არ გამოვლინდნენ ორგანიზმში, მაგრამ ხდება მათი შენარჩუნება და მემკვიდრეობით გადაცემა, სადაც კვლავ შესაძლოა მათი გამოვლინება. ყოველ ორგანიზმს რომელიმე ნიშან-თვისების განმსაზღვრელი გენის ორი ასლი აქვს, რომლისგანაც ერთს დედისგან იღებს, მეორეს კი - მამისაგან. მაგალითად, ყვითელი და მწვანე მარცვლებიანი მცენარეების შეჯვარების შედეგად მიღებულ ყველა მცენარეს ყვითელი მარცვლები აქვს: თუმცა ისინი შეიცავენ როგორც ყვითელი, ასევე მწვანე ფერის განმსაზღვრელ გენებს, მათგან მხოლოდ ყვითელი გამოვლინდება. ასეთი მცენარეების თვითდამტვერვის შედეგად მიღებული შთამომავლობიდან ზოგიერთს მწვანე ფერის მარცვლები უვითარდება. ეს იმის გამო ხდება, რომ, როგორც აღვნიშნეთ, პირველ თაობაში ყველა მცენარეს აქვს როგორც ყვითელი, ასევე მწვანე ფერის განმსაზღვრელი გენი („ფაქტორი“, მენდელის ტერმინოლოგიით), და ამ თაობის თვითდამტვერვისას ეს ფაქტორები ერთმანეთისაგან დაითიშება და შთამომავლობას თითოეული მშობლისაგან ან მხოლოდ ყვითელი ფერის განმსაზღვრელი გენი გადაეცემა, ან მხოლოდ მწვანე ფერისა. ამის შედეგად შესაძლოა ისეთი მცენარეების წარმოქმნა, რომლებშიც ორივე გენი მწვანე ფერის განმსაზღვრელია, და სწორედ მათში ვითარდება მწვანე მარცვლები.
ნიშან-თვისებათა დამოუკიდებლად გადაცემის კანონი. ამ კანონის თანახმად, ყოველი ფაქტორის (გენის) მემკვიდრეობით გადაცემა სხვა ფაქტორებისაგან დამოუკიდებლად ხდება. მენდელის ექსპერიმენტის შემთხვევაში ეს მართლაც ასე იყო: მაგ., მარცვლის ფერი და ფორმა ერთმანეთისაგან დამოუკიდებლად გადაეცემოდა შემდეგ თაობას. გავიხსენოთ, რომ მენდელი 7 ნიშან-თვისების მემკვიდრეობითობას იკვლევდა; მან ყველა შემთხვევაში ისეთი სტატისტიკური შედეგები მიიღო, რომლებიც ერთმანეთისაგან დამოუკიდებელი მემკვიდრეობითობით აიხსნებოდა. აქ აუცილებელია აღინიშნოს ერთი გარემოება: ბარდას 7 წყვილი ქრომოსომა აქვს, და როგორც შემდგომში აღმოჩნდა, მენდელის არჩეულ ყველა ნიშანზე თითო ქრომოსომა „მოდიოდა“ (ანუ ამ ნიშნების განმსაზღვრელი გენებიდან ყველა ცალკ-ცალკე ქრომოსომაზე იყო განლაგებული). მენდელს რომ თავისი ანალიზისათვის ნებისმიერი მერვე ნიშანი დაემატებინა, მაშინ სულ ცოტა ორი გენი ერთსა და იმავე ქრომოსომაზე აღმოჩნდებოდა და არა ერთმანეთისაგან დამოუკიდებლად, არამედ ერთად (ე.წ. შეჭიდულობის პრინციპის თანახმად) გადაეცემოდა შემდეგ თაობებს.
ამ ანალიზის შედეგები, რომლებიც მენდელმა ბრუნის საბუნებისმეტყველო საზოგადოებას 1864 წლის 8 თებერვალს წარუდგინა, საფუძვლად დაედო ყველა მომდევნო შრომას მემკვიდრეობითობის შესახებ. თუმცა მენდელის შრომებს მყისიერი აღიარება არ ერგოთ, და მეტად დასანანია, რომ ეს აღმოჩენები დარვინის თვალთახედვის მიღმა დარჩა: დარვინისა და მენდელის იდეების შერწყმა (როგორც ეს თანამედროვე მეცნიერებაში ხდება) ევოლუციის თეორიას განავითარებდა და სრულყოფას მიუახლოვებდა.
შემდგომში მენდელი შეეცადა, თავისი ექსპერიმენტების შედეგები სხვა ცოცხალ ორგანიზმებზე (როგორც მცენარეებზე, ასევე ცხოველებზე) შეემოწმებინა. ამ კვლევების შედეგები გაცილებით ნაკლებად წარმატებული იყო, ალბათ იმის გამოც, რომ კვლევის ობიექტებს არ გააჩნდათ ის თვისებები, რომლებიც მენდელმა ბარდის ანალიზის დროს დიდი გულმოდგინებით შეარჩია.
1868 წელს მენდელი მონასტრის აბატად აირჩიეს, და ადმინისტრაციული სამუშაოს მოზღვავებამ იგი აიძულა, ექსპერიმენტებისათვის თავი დაენებებინა. მენდელი თავს დავალებულად თვლიდა მონასტრისაგან და ამ ვალდებულების გამო ის თავს უფლებას ვერ მისცემდა, ახალი თანამდებობის მოვალეობები თუნდაც ნაწილობრივ უგულებელყო. აბატად არჩევა იმის ნიშანიც იყო, თუ რა დიდი ავტორიტეტი ჰქონდა მენდელს ბრუნის მოქალაქეთა შორის. მენდელის აბატად ყოფნის ერთ-ერთი ღირსშესანიშნავი ეპიზოდი გახლავთ მის მიერ იმ გადასახადზე უარის თქმა, რომელიც ბისმარკის მთავრობამ კათოლიკურ მონასტრებს 1873 წელს დაუნიშნა. ისტორიკოსთა აზრით, მენდელს მშვენივრად ესმოდა ასეთი უარის უნაყოფობა (საბოლოოდ მთავრობამ სამონასტრო ქონებას ყადაღა დაადო), მაგრამ მისთვის ეს პრინციპული პოზიციის გამოხატულება იყო, რადგან ამ გადასახადებს უსამართლოდ თვლიდა. მაშინაც კი, როდესაც მთავრობის წარმომადგენლებმა მას კომპრომისის სანაცვლოდ სენატორობა შესთავაზეს, მენდელმა უარი განაცხადა და უსამართლო გადასახადის წინააღმდეგ ბრძოლისათვის თავი არ დაუნებებია. საბოლოოდ ამ ხანგრძლივმა კონფლიქტმა მის ჯანმრთელობასაც დაასვა დაღი, და 1884 წლის 6 იანვარს გრეგორ იოჰან მენდელი ქრონიკული ნეფრიტის შედეგად გარდაიცვალა.

ნახ. 3: მენდელის ძეგლი ბრნოს მონასტერში.
მენდელის ცხოვრების თითქმის ყველა ეტაპს გმირობის კვალი ატყვია: ჭაბუკობის წლებში, სიღარიბის პირობებში, სწრაფვა განათლების შესაძენად; მოწიფულობისას - უაღრესად რთული ექსპერიმენტების ჩატარება და დასრულება, დაბრკოლებებისა და იმედგაცრუების მიუხედავად; სიცოცხლის ბოლო ათწლეულის განმავლობაში - ბრძოლა უსამართლო დაბეგვრის წინააღმდეგ სამონასტრო ქონების დასაცავად. გრეგორ მენდელის გარდაცვალებისას მისი, როგორც მეცნიერის, სახელი უცნობი იყო, და მხოლოდ გარდაცვალებიდან 35 წლის შემდეგ მას მიეგო დამსახურებული პატივი: მენდელის კანონები ხელახლა იქნა აღმოჩენილი. გენეტიკური კვლევების შედეგად მეცნიერებმა მრავალი ახალი კანონზომიერება დაადგინეს და არაერთი მოლეკულური მექანიზმი განსაზღვრეს, მაგრამ ბიოლოგიის ამ დარგის დამფუძნებლის წოდება ურყევადაა დაკავშირებული გრეგორ მენდელის სახელთან. თვითონ მენდელი კი, თანამედროვეთა მოგონებების მიხედვით, თავისი შრომების მნიშვნელობაში სრულიად დარწმუნებული იყო და მეგობრებთან საუბრისას ხშირად ამბობდა: „ჩემი დროც მოვა“. მალე მისი აღმოჩენიდან 150 წელი შესრულდება, და მენდელის წვლილი გენეტიკის როგორც კლასიკურ, ასევე თანამედროვე ასპექტებში (გენური ინჟინერია, გენმოდიფიცირებული ორგანიზმები) ჭეშმარიტად განუზომელია.
გელა თევზაძე.
24 საათი 30.07.11
|
|
14 ევოლუცია: დარვინის თეორია და მისი განვითარება. |
▲ზევით დაბრუნება |
I. რა უნდა იცოდეს ყველა ადამიანმა ევოლუციის შესახებ - და რატომ?
წინამდებარე სტატია (და ალბათ, რამდენიმე მისი მომდევნოც) ევოლუციის საკითხებს (და ზოგადად, ევოლუციის პროცესის სხვადასხვა ინტერპრეტაციაც) ეძღვნება. შევეცდები შემოგთავაზოთ როგორც ორიგინალური სტატიების თარგმანი, ასევე ნაშრომების ერთობლიობის მომოხილვაც და ამ ნაშრომების შემაჯამებელი კომენტარებიც. დღევანდელი სტატია კარგ ამოსავალ წერტილად მიმაჩნია ამ თემაზე მსჯელობისათვის.

ნახ. 1: ჩარლზ დარვინი (1809-1882).

ნახ. 2: „სახეობათა წარმოშობის“ პირველი გამოცემის (1859) თავფურცელი.
1860 წელს, როდესაც დარვინის „სახეობათა წარმოშობის“ („The Origin of Species“, 1859) გამოცემიდან ერთი წელი იყო გასული, ინგლისის ერთ-ერთი ეპისკოპოსის მეუღლეს უთქვამს: „ჩემო ძვირფასებო, მაიმუნის შთამომავლებო! იმედი ვიქონიოთ, რომ ეს მართალი არაა, მაგრამ თუ ეს სიმართლეა, ვილოცოთ, რომ ეს ამბავი საყოველთაოდ ცნობილი არ გახდეს“. თანამედროვე დიდ ბრიტანეთით ეს მანდილოსანი ალბათ უკმაყოფილო დარჩებოდა, მაგრამ ამერიკის შეერთებულ შტატებში კი დღეს კარგად იგრძნობდა თავს: 2004 წლის გამოკითხვის შედეგების მიხედვით (Gallup poll, November 2004), რესპონდენტების 45%25 თავს კრეაციონისტად თვლიდა. კიდევ 38%25 თეისტური ევოლუციის მოძღვრებას იზიარებდა, ანუ მიაჩნდა, რომ ადამიანის ევოლუცია მილიონობით წლის განმავლობაში გრძელდებოდა, მაგრამ ეს პროცესი ღვთის ნებით წარიმართებოდა. და მხოლოდ 13%25-სათვის იყო მისაღები ევოლუციის ე.წ. „სრულიად ნატურალისტური“ მოდელი, რომლის თანახმადაც ევოლუცია ზეგარდმო ძალებისაგან დამოუკიდებლად მიმდინარეობს.
უკვე დროა, რომ ბუნებისმეტყველების მასწავლებლებმა მეტი დაჟინებით მოითხოვონ, რომ ევოლუციის თეორია საუნივერსიტეტო განათლების უცვლელი ნაწილი გახდეს, მათ შორის ჰუმანიტარული ფაკულტეტებისთვისაც. ამ დებულების საფუძველი მხოლოდ ფაქტების შესწავლას არ ემყარება. ცნობილია, რომ მრავალი ჰუმანიტარული ფაკულტეტის პროგრამაში შეტანილია ბიოლოგიის კურსი. ასეთი კურსის გავლის შედეგად, სტუდენტებს ექმნებათ ზოგადი წარმოდგენა დარვინის თეორიაზე, მის ზოგიერთ - შედარებით ადვილად გასაგებ - მხარდამჭერ ფაქტზე და, შესაძლოა, ამ თეორიის პრაქტიკული გამოყენების რამდენიმე მაგალითზე. მიუხედავად ამისა, ბიოლოგიის მასწავლებლების უმეტესობა ევოლუციის თემაზე საუბრისას საკმარისად ნათლად არ გამოყოფს რამდენიმე უმნიშვნელოვანეს საკითხს. ამ სტატიაში აღწერილია 4 შესაძლო პასუხი კითხვაზე: „რა უნდა იცოდეს ყველა სტუდენტმა (და ალბათ, ყველა ადამიანმა) ევოლუციის შესახებ?“ გასათვალისწინებელია ის გარემოებაც, რომ თვით ევოლუციის იდეა ხშირად სკეპტიციზმის ან, უფრო მეტიც, აგრესიული დამოკიდებულების გამომწვევია.
ჰუმანიტარული ფაკულტეტების ერთ-ერთი მიზანია, სტუდენტებმა გაითავისონ ის დებულება, რომ „სწავლისა თუ ცოდნის შეძენის მრავალი გზა არსებობს“, ისევე როგორც გაეცნონ იმ იდეებს, რომლების გავლენით შევძელით, უკეთ შეგვეცნო როგორც ადამიანი, ისევე თვით ბუნება. ამის გამო, პირველი მნიშვნელოვანი საკითხი, რომლის გათვითცნობიერებაც აუცილებელია, შემდეგში მდგომარეობს: დარვინის თეორიამ ევოლუციის შესახებ შეცვალა ადამიანების აზრი საკუთარ ადგილზე სამყაროში. საუკუნეების განმავლობაში, ფილოსოფოსები განიხილავდნენ საკითხს ადამიანის ადგილის შესახებ სამყაროში და ადამიანის სხვასდასხვა შესაძლო განსაზღვრებების შესახებ ბჭობდნენ. მატერიალისტური ფილოსოფიის შეხედულება იმის შესახებ, რომ ადამიანის წარმოშობას მხოლოდ ბუნებრივ (და არა ზებუნებრივ) ძალებს უნდა ვუმადლოდეთ, საყოველთაოდ მიღებული არაა. მაგრამ იდეის მიღება აუცილებელი არაა, რომ მისი გავლენა ვაღიაროთ. თუნდაც ფილოსოფიური თვალსაზრისით, უნივერსიტეტის ყველა კურსდამთავრებულმა უნდა იცოდეს დარვინისეული ევოლუციის შესახებ.
ცოდნა ევოლუციის შესახებ ასევე გვეხმარება გავაცნობიეროთ მეცნიერება, როგორც ცოდნის შეძენის გზა. მაგალითად, ყველასათვის ნათელია, რომ მეცნიერის უდიდეს მიღწევად ითვლება საყოველთაოდ აღიარებული თეორიის ჩამოყალიბება და განვითარება? ცნობილია, რომ თეორიები ერთმანეთისაგან განსხვავდება იმით, თუ რამდენად დამაჯერებლად ხსნიან ამათუიმ მოვლენას, ისევე როგორც მათი მხარდამჭერი ფაქტების რაოდენობით. მაგრამ მხოლოდ რამდენიმე თეორიას უდევს საფუძვლად ურყევი ფაქტები და მხოლოდ რამდენიმე თეორია პოვებს გამოძახილს მეცნიერების რომელიმე მთავარი დარგის ყველა ნაწილში. ამგვარი უმნიშვნელოვანესი თეორიების მაგალითებია ევოლუცია, თერმოდინამიკა, მოლეკულური ორბიტალები, მიზიდულობის ძალა, ფარდობითობა, ქვანტური ელექტროდინამიკა, და გეოლოგიური ფილების ტექტონიკა. თითქმის შეუძლებელია ბიოლოგიაში ისეთი დაკვირვების ან ჰიპოთეზის დასახელება, რომელსაც ევოლუციასთან არავითარი კავშირი არ ექნება.
ყველა მნიშვნელოვანი მეცნიერული თეორია მნიშვნელოვანწილად (თუ გამონაკლისის გარეშე არა) დასკვნებს ემყარება. ესაა მეორე დებულება, რომელიც როგორც ევოლუციის თეორიას, ასევე ზოგადად მეცნიერებას მიესადაგება. ექსპერიმენტების შედეგებზე უშუალო დაკვირვებაა შესაძლებელი, მაგრამ კვლევის ობიექტებზე ასეთი დაკვირვება უმეტესწილად შესაძლებელი არაა. ამ დებულების ახსნისას, სტუდენტებისათვის ასეთი მაგალითის მოყვანა იქნება მიზანშეწონილი: მოლეკულური ორბიტალების თეორია ხსნის, თუ როგორ მოქმედებენ ელექტრონები და როგორც წარმოიქმნება ქიმიური ბმები ატომებს შორის, რის შედეგადაც ატომები მოლეკულებად ერთიანდებიან. ამ მოვლენების უშუალო დაკვირვება შეუძლებელია, მაგრამ ელექტრონებისა და ქიმიური ბმების არსებობა საყოველთაოდაა აღიარებული მეცნიერთა მიერ, რადგან ეს თეორია დამაჯერებლად ხსნის მრავალ მოვლენას და, რაც მთავარია, ამ თეორიის საფუძველზე ჩამოყალიბებული ჰიპოთეზები თუ ვარაუდები გამონაკლისის გარეშე მართლდება. ქიმიკოსებს რომ ნატრიუმის ტუტესა და მარილმჟავას შერევის მეტი არაფერი ეკეთებინათ (აუცილებელია აღინიშნოს, რომ ამ ნაერთების შერევა ლაბორატორიის ფარგლებს გარეთ რეკომენდირებული არ არის), მათთვის უაღრესად ძნელი იქნებოდა, თბილი წყლის წარმოქმნა ამ შერევის შედეგად დაეკავშირებინათ ატომებს შორის ქიმიური ბმების ცვლასთან. საბედნიეროდ, ქიმიკოსებს მრავალი სხვა ექსპერიმენტიც აქვთ ჩატარებული და მათი უმეტესობა აღწერილზე გაცილებით უფრო ღრმააზროვანი და რთულია, რამაც ატომური ორბიტალების თეორიის წარმოქმნას და მის დახვეწას შეუწყო ხელი.
სტუდენტებს ასევე უნდა შევახსენოთ, რომ საშუალო თუ უმაღლეს სკოლაში მათ არ შეუსწავლიათ ის მნიშვნელოვანი ფაქტები, რომლებიც მოლეკულური ორბიტალების თეორიის მხარდამჭერად არიან მიჩნეული. მეტიც, მათ ალბათ არც უსწავლიათ საშუალო სკოლაში მოლეკულური ორბიტალების შესახებ. მოსწავლეები ალბათ ვერც გაიგებდნენ იმ ექსპერიმენტების არსს, რომლების შედეგადაც ეს თეორია ჩამოყალიბდა. რაც მთავარია, არცერთი ამ ექსპერიმენტთაგან არ გვაძლევს ელექტრონებისა თუ ქიმიური ბმების დანახვის შესაძლებლობას. აღსანიშნავია, რომ მოსწავლეებსა და სტუდენტებს ევოლუციის თეორიის მხარდამჭერი ფაქტების გაცილებით მეტი რაოდენობა მიეწოდებათ, ვიდრე ნებისმიერი სხვა თეორიის საფუძვლების წარმომქმნელი ფაქტებისა. სტუდენტებმა უნდა გააცნობიერონ, რომ თეორიის დასაბუთება დასკვნებზე დაყრდნობით უჩვეულო არაა.
მესამე ყურადსაღები ფაქტი ევოლუციის თეორიის შესახებ ისაა, რომ ყველა მნიშვნელოვანი თეორიის მსგავსად, მას მუდმივი წინააღმდეგობა ხვდებოდა ახალი ფაქტების სახით, რომლებიც, ერთი შეხედვით, ამ თეორიის გაბათილების საფუძველს იძლეოდნენ. რასაკვირველია, აქ ისეთი იდეები და შედეგები გვაქვს მხედველობაში, რომლებიც ლოგიკას არ ეწინააღმდეგება, და რომლებიც აიძულებენ მეცნიერებს, არსებული შეხედულებები გადასინჯონ. ბიოლოგებმა მზადყოფნით აღიარეს საერთო მემკვიდრეობის (common descent) ფაქტი, როგორც ევოლუციის თეორიის საფუძველი. მაგრამ სტუდენტებს ისიც უნდა მოვუთხროთ, რომ „შეგუება ბუნებრივი გადარჩევის შედეგად“ (ერთ-ერთი უმნიშვნელოვანესი მეცნიერული იდეა, რომელიც ოდესმე წარმოქმნილა) სულაც არ იყო უპირობოდ აღიარებული, და არა მარტო რელიგიური მოსაზრებების საფუძველზე. დარვინის გარდაცვალებიდან ათწლეულები იყო გასული, როცა მენდელისეული მემკვიდრეობის კანონები ხელმეორედ იქნა აღმოჩენილი, ეს კანონები და დარვინის თეორია ბუნებრივი გადარჩევის შესახებ შეუთავსებლად იქნა მიჩნეული. მუტაციები, რომლებიც მენდელის კანონების თანახმად გადაეცემოდნენ შთამომავლობას, მხოლოდ მნიშვნელოვან განსხვავებებს აპირობებდნენ, დარვინი კი ამტკიცებდა, რომ ევოლუციურად მნიშვნელოვანი ბუნებრივი მრავალფეროვნება (natural variation) ჩვეულებრივ საკმაოდ ძნელი შესამჩნევია. ბიოლოგებმა მხოლოდ 1920-30-იან წლებში შესძლეს შეეთავსებინათ ბუნებრივი გადარჩევა და კლასიკური გენეტიკა იმგვარად, რომ ეს ორი შემადგენელი ნაწილი ევოლუციის თეორიაში გაერთიანებულიყო (ეს წინააღმდეგობა გენეტიკასა და ევოლუციის თეორიის შორის, ისევე როგორც მისი გადაჭრის გზა, ერთ-ერთი მომავალი სტატიის თემა იქნება - GT).

ნახ. 3: მოტოო კიმურა (1924-1994).
კიდევ რამდენიმე ათწლეულის შემდეგ, ბუნებრივი გადარჩევის თეორიის ჭეშმარიტებაში დაეჭვდა მოტოო კიმურა, რომელის ამტკიცებდა, რომ დნმ-ის (გენეტიკური მასალის) ხილული ცვლილებების უმეტესობა სელექტიურად (და, მაშასადამე, ევოლუციის თვალსაზრისით) ნეიტრალურია. კიმურას იდეებიც გამართლდა, მაგრამ ამას ევოლუციის თეორიის გაბათილება სულაც არ მოჰყოლია: პირიქით, კიმურას დასკვნებმა ევოლუციის მოლეკულური მექანიზმების უკეთესი ახსნა უზრუნველყო, და ექსპერიმენტების ახალი მიმართულებებიც დასახა. ისევე, როგორც აინშტაინმა დახვეწა ნიუტონისეული მექანიკა, კიმურამ დახვეწა დარვინისეული ევოლუცია (კიმურას წვლილს და მის მეცნიერულ მემკვიდრეობასაც ერთ-ერთი სტატია მიეძღვნება - GT).
ასევე უნდა ვიცოდეთ, რომ მეცნიერები უარყოფენ „გონივრული დაგეგმვისა“ (intelligent design) და კრეაციონიზმის იდეებს არა იმიტომ, რომ მათ ძალიან ბევრი აქვთ „დაბანდებული“ ბუნებრივი გადარჩევის თეორიაში. „გონივრული დაგეგმვის“ იდეა უარყოფილია, რადგან იგი ვერ მოგვცემს ბუნების შეცნობის შესაძლებლობას: თუ ჩვენ დავუშვებთ, რომ რომელიმე ორგანიზმი „გონივრული დაგეგმვის“ საფუძველზეა შექმნილი, ამით ყოველგვარი კვლევა სრულდება. მაგრამ თუ ევოლუციის საბუთი გვაქვს, ეს მომავალი ექსპერიმენტების ამოსავალ წერტილს წარმოადგენს. და მართლაც, ევოლუციურ ბიოლოგიაში მუდმივად ხდება ახალი ფაქტების ძიება - როგორც არსებული დასკვნების მხარდამჭერის, ასევე მათი საწინააღმდეგო ფაქტებისა. ზოგადად, მეცნიერებისათვის ყველაზე მეტად ის იდეებია მომხიბვლელი, რომელთა ჭეშმარიტების შემოწმება მუდმივად გრძელდება.
მეოთხე დებულება ევოლუციის შესახებ იმაში მდგომარეობს, რომ ევოლუციას უდაოდ დიდი მნიშვნელობა აქვს ადამიანთა ყოველდღიური ცხოვრების თვალსაზრისითაც. პათოგენური მიკროორგანიზმები, რომლებიც ევოლუციის შედეგად ანტიბიოტიკებისადმი მდგრადობას იძენენ, მრავალი ადამიანის ავადმყოფობისა თუ დაღუპვის მიზეზია. მწერებიც ევოლუციის შედეგად ივითარებენ მდგრადობას თითქმის ყველა საწამლავის წინააღმდეგ, რომლებსაც ადამიანები მათ წინააღმდეგ იყენებენ. ამ საწამლავების დოზის გაზრდა, საბოლოოდ, მხოლოდ იმას იწვევს, რომ ხილი თუ ბოსტნეული სულ უფრო და უფრო მეტ ქიმიკატებს შეიცავს და საკვებად გამოყენებისათვის სიფრთხილეა საჭირო.
ევოლუციის მნიშვნელობა სახეობათაშიდა ცვალებადობით (variation within species) არ შემოიფარგლება. ბიოლოგიური კვლევის მოდელური ორგანიზმები - თაგვი, საფუარი სოკო, დროზოფილა (ხილის ბუზი) - მხოლოდ ექსპერიმენტებისათვის მოსახერხებელი ობიექტები არ არიან და არა მარტო ამ მოსაზრებით იყვნენ არჩეული. იმის გამო, რომ ამ ორგანიზმებსა და ადამიანებს საერთო წინაპრები ჰყავთ, „მათ და ჩვენ“ მრავალი საერთო თვისება გვახასიათებს. რასაკვირველია, ადამიანების მსგავსება თაგვებთან (ოთხსაკნიანი გული, ელენთა) მეტია, ვიდრე დროზოფილასთან, და სწორედ ამიტომ თაგვები შესანიშნავ მოდელს წარმოადგენენ ადამიანის გულის დაავადებისა თუ დიაბეტის შესასწავლად. მაგრამ თუ კვლევის მიზანი თვალის განვითარების შესწავლაა, დროზოფილაც მეტად საჭირო ინფორმაციას მოგვაწვდის. საფუარ სოკოსაც კი ადამიანებთან ბევრი საერთო თვისება აქვს - გაცილებით მეტი, ვიდრე ბაქტერიებს ან თუნდაც მცენარეებს. ამიტომ, საფუარის შესწავლის შედეგად მიღებული დასკვნები (უჯრედის ზრდისა და გაყოფის, ცილების სინთეზისა და დაშლის, გარემოს ცვლილებაზე რეაგირების მექანიზმების შესახებ) ადამიანის ორგანიზმსაც მიესადაგება.
რატომაა უფრო ადვილი ტუბერკულოზის ან ჯილეხის განკურნება, ვიდრე მალარიისა? ტუბერკულოზსა და ჯილეხს ბაქტერიები იწვევს, ხოლო მალარიის გამომწვევი ერთუჯრედიანი ცხოველია. იმის გამო, რომ ერთუჯრედიანი ცხოველები (პროტისტები) ადამიანის უფრო „ახლო ნათესავებია“, გაცილებით რთულია ისეთი წამლის პოვნა, რომელიც პროტისტებს დააზიანებს, ადამიანისათვის კი უვნებელი იქნება. ტრიქინების (ჭიების, მრავალუჯრედიანი ცხოველების) წინააღმდეგ ამგვარი წამლის პოვნა კიდევ უფრო რთულია, რადგან ეს ჭიები „ევოლუციურ კიბეზე“ ადამიანთან პროტისტებზე კიდევ უფრო ახლოს იმყოფებიან.
თუ დავუშვებთ, რომ განათლების მიზანია იმ იდეების დანერგვა, რომლებიც სამყაროს შეცნობას უწყობს ხელს, ყველა განათლებულმა ადამიანმა უნდა გააცნობიეროს ეს დებულებები ევოლუციის შესახებ. თუ ბიოლოგიის მასწავლებლები დანერგავენ ცოდნისა და სწავლისადმი სწრაფვას, შესაძლოა, მოსწავლეთაგან ზოგიერთნი მოინდომებენ როგორც „სახეობათა წარმოშობის“ წაკითხვას, ასევე გაეცნობიან თანამედროვე ნაშრომებს ევოლუციის შესახებ. ამის შედეგად კი სავსებით დასაშვებია, რომ კრეაციონისტების მიერ მოშველიებული რიტორიკული არგუმენტები მათთვის დამაბნეველი აღარ იქნება.
გელა თევზაძე.
24 საათი 26.08.11
|
|
15 ევოლუცია: დარვინის თეორია და მისი განვითარება. II. დარვინის იდეების გავლენა ევოლუციის კვლევაზე (ნაწილი I) |
▲ზევით დაბრუნება |
.
დარვინის „სახეობათა წარმოშობამ“ დაარწმუნა მსოფლიო ევოლუციის არსებობაში. დარვინისეული თეორია ბუნებრივი გადარჩევის შესახებ კი გაცილებით მეტ წინააღმდეგობას შეხვდა: მემკვიდრეობითობის მექანიზმის უცოდინრობა, ბუნებაში არსებული ცვალებადობის მექანიზმების არასრულყოფილი გაგება და, რაც მთავარია, მტკიცებულებების უქონლობა ამ თეორიის მოწინააღმდეგეთა რიცხვის გამრავლებას უწყობდნენ ხელს. ევოლუციის როგორც უწყვეტი, ასევე წყვეტილი განვითარების მხარდამჭერი არგუმენტები მრავლადაა ევოლუციის თეორიის ისტორიაში. ამ სტატიის მიზანია ასეთი არგუმენტების, მათი წარმოშობისა და განვითარების მიმოხილვა.

ნახ. 1: ერაზმუს დარვინი (1732-1802).
შესაძლებლობა და აუცილებლობა. ჩარლზ დარვინი არ გახლდათ პირველი ადამიანი, რომელმაც გამოთქვა იდეა, რომ ბუნება ევოლუციური გზით ვითარდება. ბუფონი (1707-1788), ლამარკი (1744-1829), და თვით დარვინის ბაბუა ერაზმუს დარვინი (1732-1802) ევოლუციის მომხრენი იყვნენ და მათი იდეები „სახეობათა წარმოშობა“-მდე (1859) ორი ან მეტი თაობით ადრე გამოითქვა. დარვინის თანამედროვემ რობერტ ჩემბერსმა (Robert Chambers, 1802-1871) გამოაქვეყნა „მტკიცებულებები ქმნილებათა ბუნებრივი ისტორიის შესახებ“ („Vestiges of the Natural History of Creation“, 1844), რომელშიც ეოლუციის მხარდამჭერი არგუმენტები იყო მოყვანილი. ეს წიგნი 10-ჯერ გამოიცა და პირველი გამოცემიდან 10 წლის განმავლობაში 15 ათას ცალზე მეტი გაიყიდა. შედარებისათვის, დარვინის „სახეობათა წარმოშობა“ გამოქვეყნებიდან პირველ ათწლეულში 5-ჯერ გამოიცა და ამ წიგნის 9250 ცალი გაიყიდა.
დარვინამდელ ევოლუციონისტებს ორი ძირითადი თვისება აერთიანებდათ: 1) ვერცერთმა მათგანმა ვერ მოახერხა დაერწმუნებინა მეცნიერები, რომ მცენარეებისა და ცხოველების განსაკუთრებული ნიშან-თვისებების ევოლუცია მოხდა სიცოცხლის განსხვავებული ფორმების საფუძველზე. 2) მეორე გარემოება, რომელიც მეტად მნიშვნელოვანია დარვინის წვლილის გათვითცნობიერებისათვის, იყო დარვინამდელი მეცნიერების აზრი, რომ ევოლუციას წარმართავდა რაღაც გარეშე ძალა, იქნებოდა ეს ზებუნებრივი ნება თუ გაუცნობიერებელი ბიოლოგიური სწრაფვა მეტი სირთულისაკენ. ასეთი ძალის არსებობის აღიარებით დარვინამდელი მეცნიერები ხარკს უხდიდნენ დასავლური ცივილიზაციის ერთ-ერთ უძველეს ტრადიციას. ამ ტრადიციის მიმდევარნი ამტკიცებდნენ, რომ ბუნება, ისევე როგორც ადამიანის ეთიკა ვერ წარმოიქმნებოდა საგნებისა თუ სხეულების მექანიკური („არაზებუნებრივი“) ურთიერთქმედებისა თუ გარდაქმნის შედეგად. ამავე შეხედულებით, ბუნებაში არსებობს განვითარების მიზანი და გეგმა, რაც განუყოფელია ადამიანის ეთიკისაგან. ქრისტიანული თეოლოგიის თანახმად, ღმერთმა შექმნა ცა და ხმელეთი, მცენარეები და ცხოველები, და ასევე დაადგინა მორალური კანონები ადამიანებისათვის.
დასავლურ კულტურაში არსებობდა ღრმა რწმენა, რომ ალბათობა და აუცილებლობა საკმარისი არაა ცოცხალი ორგანიზმების არსებული მრავალფეროვნების (და ამ მრავალფეროვნების კანონზომიერებების) წარმოსაქმნელად. ამ აზრს იზიარებდნენ ბუნებისმეტყველები, თეოლოგები, სოციოლოგები და ფილოსოფოსები. მაგალითად, მონტესკიე (1689-1755) ამტკიცებდა, რომ ნიადაგი, კლიმატი, და სოციალური პირობები ნაწილობრივ განსაზღვრავდნენ საზოგადოების ეთიკურ ნორმებს: „ისინი, რომელნიც ამტკიცებენ, რომ ბრმა შემთხვევითობას ძალუძს [სიცოცხლის] მრავალფეროვნების წარმოქმნა, აბსურდულად მსჯელობენ; მართლაც, რა უნდა იყოს უფრო უაზრო, ვიდრე იმის მტკიცება, რომ ბრმა შემთხვევითობამ შესაძლოა წარმოქმნას გონიერი არსებები?“
დარვინის ახალგაზრდობისას არსებული „ბუნების თეოლოგია“ ნახსენები ტრადიციის ყველაზე მარტივი ვერსია გახლდათ. ეპისკოპოსი უილიამ პეილი (William Paley, 1743-1805) თავის ნაშრომში („Natural Theology“, 1802) აცხადებდა, რომ მხოლოდ ის ფაქტი, რომ ცოცხალი ორგანიზმების ადაპტაცია არსებობს, დამაჯერებლად ამტკიცებს ღვთის არსებობასა და მის კეთილგანწყობას. საინტერესოა, რომ დარვინის მოგონებების საფუძველზე, პეილის „ბუნების თეოლოგია“ სტუდენტობისას წაკითხულ წიგნთაგან ყველაზე შთამბეჭდავი გახლდათ.
დარვინის „სახეობათა წარმოშობა“ მნიშვნელოვნად განსხვავდებოდა მისი წინამორბედი ევოლუციონისტების ნაშრომებისაგან. „სახეობათა წარმოშობამ“ მრავალი მეცნიერი დაარწმუნა, რომ ევოლუცია „ორგანული გზით“ წარიმართა. დარვინმმა შესძლო, შეექმნა ევოლუციის თეორია, რომელიც ყველა არსებულ შეხედულებაზე უფრო დამაჯერებელი იყო, და ამ დანაჯერებლობას მრავალი მხარდამჭერი არგუმენტის მოშველიების გზით მიაღწია (მაგ., ჰიბრიდების არსებობა, სტერილურობის სხვადასხვანაირი გამოვლენა ჰიბრიდებში, გეოლოგიური მონაცემები, გეოგრაფიული გავრცელება, ემბრიოლოგიური ფაქტები). „მეცნიერული ანტიევოლუციონისტები“ საკმაოდ წამგებიან მდგომარეობაში აღმოჩნდნენ „სახეობათა წარმოშობის“ გამოცემიდან პირველივე ათწლეულის განმავლობაში.
ევოლუცია არსებობს, მაგრამ არსებობს თუ არა ბუნებრივი გადარჩევა? დარვინმა წარმატებულად დაარწმუნა თავისი მკითხველები ევოლუციის არსებობაში, მაგრამ ევოლუციის მექანიზმის მისეულ ახსნას (ბუნებრივ გადარჩევას) თავიდანვე გაცილებით ნაკლები აღიარება ხვდა წილად. თავისი თეორიის ჩამოყალიბებიდან (1837) ერთ წელზე ნაკლები იყო გასული, როცა დარვინი აღნიშნავდა, რომ ბუნებრივი გადარჩევა სწორედ ის მექანიზმია, რომელიც მხოლოდ ალბათობისა და აუცილებლობის საფუძველზე მოქმედებს და რომლის არსებობას დარვინამდელი მოაზროვნენი მტკიცედ უარყოფდნენ. დარვინის აზრით, ბუნებრივი გადარჩევა იყო ძირითადი მექანიზმი, რომლის მოქმედებაც უზრუნველყოფს ორგანიზმების „გონივრულ დაგეგმვას“. ეს „არაზებუნებრივი“, არამართვადი, და შემთხვევითობაზე დაფუძნებული მექანიზმი წარმოქმნიდა იმ მრავალფეროვნებას, რომელსაც „ბუნების თეოლოგები“ ღვთიურ ძალას მიაწერდნენ, და რომელიც მრავალთათვის წარმოადგენდა მიზნისა თუ დაგეგმვის გამოსახულებას. თუნდაც ამ მიზეზით აიხსნება სწრაფი და ძლიერი უარყოფითი რეაქცია ბუნებრივი გადარჩევის იდეის მიმართ. დარვინმა შესძლო, მხოლოდ რამდენიმე (თუმცა საკმაოდ გავლენიანი) მეცნიერი დაერწმუნებინა, რომ ბუნებრივი გადარჩევა ევოლუციის ძირითადი მექანიზმია.

ნახ. 2: უილიამ პეილის „ბუნებრივი თეოლოგიის“ თავფურცელი.
პირდაპირი და არაპირდაპირი მტკიცებულებები. ბუნებრივი გადარჩევის იდეის მოწინააღმდეგეებს მრავალი არგუმენტი გააჩნდათ თუნდაც იმ მიზეზით, რომ დარვინმა ვერ შესძლო, ბუნებრივი გადარჩევის დამამტკიცებელი პირდაპირი საბუთები წარმოედგინა (ყოველ შემთხვევაში, ისეთივე დამაჯერებელი საბუთები, რომლების საშუალებითაც იგი ევოლუციური გარდაქმნების არსებობას ამტკიცებდა). ბინებრივი გადარჩევის მექანიზმისათვის მას მხოლოდ არაპირდაპირი მტკიცებულებები ჰქონდა, რომლებიც დარვინმა კარგად გამოიყენა. „სახეობათა წარმოშობის“ დასაწყისში აღწერილია ხელოვნური გადარჩევის ეფექტურობა, მოყვანილია მრავალი მაგალითი იმისა, თუ როგორ შეიქმნა მტრედების ერთმანეთისაგან მნიშვნელოვნად განსხვავებული ჯიშები ადამიანის მიერ გადარჩევის შედეგად, და რომ ყველა ეს ჯიში ველური მტრედისაგან იყო გამოყვანილი. მემკვიდრეობითი ცვალებადობა (თუმცა ნაკლები სიხშირით) არსებობს ბუნებრივ პოპულაციებშიც. ყველა ორგანიზმს აქვს გეომეტრიული პროგრესიით გამრავლების უნარი, რაც იწვევს საკვებისათვის კონკურენციას, ანუ ბრძოლას არსებობისათვის. (აქვე გავიხსენოთ დარვინისეული აფორიზმი, რომლითაც მან არსებობისათვის ბრძოლა აღწერა: „ძლიერნი, ჯანმრთელნი და ბედნიერნი გადარჩებიან და გამრავლდებიან“ - „The vigorous, the healthy , and the happy survive and multiply“). თავის მხრივ, ბრძოლა არსებობისათვის ხანგრძლივი დროის დროის განმავლობაში ბუნებრივი გადარჩევის პროცესად წარმოგვიდგება. ასეთი ლოგიკა სავსებით დამაჯერებელი იყო. დარვინმა ბუნებრივი გადარჩევის ორი „ცხოვრებისეული“ მაგალითი მოიყვანა: 1) მგლების სისწრაფე და ზედმეტი წონის უქონლობა; 2) მცენარეების ევოლუცია, რომლის შედეგადაც ისინი ტკბილ ნექტარს წარმოქმნიან, და ფუტკრის იმ ორგანოების განვითარება, რომლებიც ნექტარის მოსაპოვებლადაა საჭირო. აქვე უნდა აღვნიშნოთ, რომ ამ ორის გარდა, „სახეობათა წარმოშობაში“ ბუნებრივი გადარჩევის სხვა მაგალითებს ვერ მოვნახავთ, და თვით დარვინის აღიარებით, ეს ორად-ორი მაგალითიც უფრო „წარმოსახვითი ილუსტრაციების“ (imaginary illustrations) კატეგორიას განეკუთვნება.

ნახ. 3: შინაური მტრედების ჯიშების მრავალფეროვნების ილუსტრაცია დარვინის „სახეობათა წარმოშობიდან“. ცენტრში - ველური მტრედი (wild rock pigeon), მის გარშემო - შინაური მტრედების ჯიშები.
ადამ სეჯვიკი (Adam Sedgwick, 1785-1873), რომელიც დარვინის სტუდენტობისას მისი ერთ-ერთი პროფესორი იყო (გეოლოგიაში), მრავალი მეცნიერის აზრს გამოხატავდა ბუნებრივი გადარჩევის თეორიის მხარდამჭერი მტკიცებულებების შეფასებაში: „ბუნებრივი გადარჩევის თეორია დიდი პირამიდაა, რომელიც საკუთარ წვერზე დგას, და ეს წვერი მათემატიკურ წერტილს (ანუ უაღრესად მცირე სიდიდეს - GT) წარმოადგენს“... დარვინის აზრით, ბუნებრივი გადარჩევის მოქმედების გამოსავლენად სულ ცოტა 50 წელია საჭირო. ასე რომ, ცხადია, დარვინს არა მარტო არ ჰქონდა ბუნებრივი გადარჩევის მოქმედების რაიმე საბუთი, იმის იმედიც კი არ გააჩნდა, რომ ამ მოქმედების გამოვლენას თავისი სიცოცხლის განმავლობაში ერთხელ მაინც იხილავდა.
მემკვიდრეობითობა ბუნებრივი გადარჩევის დარვინისეული თეორიის კიდევ ერთ შესაძლო წინააღმდეგობას წარმოადგენდა. ბუნებრივი გადარჩევის თეორია ემყარებოდა შეხედულებას, რომ ევოლუციის მასალაა ბუნებრივ პოპულაციებში არსებული მრავალი შემთხვევითი მემკვიდრეობითი ცვალებადობა (ტერმინი „შემთხვევითობა“ ამ ფრაზაში ევოლუციის მიმართულებისაგან დამოუკუდებლობის, საწყისი ცვალებადობის ევოლუციური ნეიტრალურობის თვალსაზრისით გამოიყენება). დარვინის აზრით, ბუნებრივი გადარჩევის ობიექტი სწორედ ეს ცვალებადობა იყო, რაც, თავის მხრივ, ევოლუციური განვითარების უმთავრესი განმსაზღვრელი ფაქტორი გახლდათ. მაგრამ დარვინი ასევე მიიჩნევდა, რომ ზოგიერთ შემთხვევაში მემკვიდრეობითი ცვალებადობა შემთხვევითი არ იყო, და ეს ცვალებადობა „გრძელვადიანი ჩვევების“ (long-continued habits) შედეგად მიღებული ფიზიკური ნიშან-თვისებების მემკვიდრეობით გადაცემის საფუძველზე ხდებოდა. ასევე, შეძენილი თვისებების მემკვიდრეობით გადაცემას დარვინისათვის გაცილებით ნაკლები მნიშვნელობა ჰქონდა, ვიდრე ბუნებრივ გადარჩევას, როგორც ევოლუციის წამმართველ ძალას. დარვინის ოპონენტები აცხადებდნენ, რომ მემკვიდრეობითობის შეძენილი თუ „მიზანმიმართული“ თვისებებისაგან დამოუკიდებლად, მემკვიდრეობით გადაცემული ცვალებადობა ადაპტაციის (ანუ შეგუების) მიმართულებით იქმნებოდა. ასე რომ, მებისმიერი თეორია, რომელიც ცვალებადობის „მიზანმიმართულებას“ აღიარებდა, ამავე დროს ამცირებდა ბუნებრივი გადარჩევის მნიშვნელობას ევოლუციურ პროცესში. მემკვიდრეობითობის ამგვარი თეორიები მრავლად აესებობდა 1860-1900 წლებში.
1868 წელს დარვინმა ჩამოაყალიბა „პანგენეზის წინასწარგანმჭვრეტი ჰიპოთეზა“ (provisional hypothesis of pangenesis, პანგენეზი - ერთიანი წარმოშობა). ეს თეორია ამტკიცებდა, რომ ორგანიზმის ყველა ნაწილი წარმოქმნიდა უმცირესი ზომის თავისუფლად მოძრავ ნაწილაკებს (გემულებს - gemmules), რომლებიც მრავლდებოდნენ და გროვდებოდნენ რეპროდუქციულ სისტემაში, საიდანაც მომავალ თაობებს გადაეცემოდნენ. აქვე გავიხსენოთ, რომ გრეგორ მენდელის „ექსპერიმენტები მცენარეთა ჰიბრიდებზე“ („Versuche über Pflanzen-Hybriden“, 1866 წ.), რომელიც თანამედროვე გენეტიკის საფუძვლადაა მიჩნეული და რომელშიც მემკვიდრეობითობის კანონების აღწერილი, დარვინისათვის, ისევე როგორც იმდროინდელ მეცნიერთა უმრავლესობისათვის, უცნობი დარჩა.
პანგენეზის ჰიპოთეზა გავლენიან, მაგრამ არადამაჯერებელ მოძღვრებად დარჩა. თითქმის ყველა თეორია, რომელიც მეცხრამეტე საუკუნის მიწურულს ჩამოყალიბდა და რომელიც მემკვიდრეობითობის მექანიზმების ახსნას ცდილობდა, პანგენეზის თეორიის განხილვითა და მისი უარყოფით იწყებოდა. თითქმის ყველასათვის ცხადი იყო, რომ ბუნებრივი ევოლუციის უმთავრესი მექანიზმი მემკვიდრეობითობის მექანიზმებს ემყარებოდა. დარვინის შემოთავაზებული მემკვისრეობითობის თეორია კი არასაკმარისად მყარი იყო და ბუნებრივი გადარჩევის ოპონენტების მხრიდან კრიტიკის იოლ სამიზნედ იქცა.
უწყვეტი თუ წყვეტილი? დარვინის შეხედულება ევოლუციის შესახებ კიდევ ერთი საკამათო საკითხი გახდა. დარვინი აღიარებდა, რომ „ბუნებრივი გადარჩევა ყოველთვის უაღრესად ნელა იმოქმედებს... ხშირად მხოლოდ დროის ვრცელი ინტერვალების განმავლობაში, და ჩვეულებრივ გარკვეული დროის მონაკვეთში რომელიმე რეგიონის მხოლოდ რამდენიმე მცხოვრებზე“. დარვინიო ასევე ამტკიცებდა, რომ „ეს უაღრესად ნელი, მხოლოდ დროდადრო მოქმედი (intermittant) მექანიზმი“ კარგად შეესაბამება გეოლოგიურ ფაქტებს იმის შესახებ, თუ როგორ იცვლებოდნენ ამ სამყაროს მაცხოვრებლები. დარვინი დაჟინებით ამტკიცებდა, რომ ევოლუცია ნელი და თანდათანობითია, და მისი მოქმედება არა მიდმივად, არამედ მხოლოდ დროის ცალკეულ ინტერვალებში გამოვლინდება. დარვინის შეხედულებით, ცვალებადობის მატარებელი ორგანიზმები ახალი სახეობის წარმოშობის წინაპირობას წარმოადგენდნენ: ბუნებრივი გადარჩევა ორგანიზმებს ეკოლოგიურ ნიშებში „ათავსებდა“, სადაც ახალი ცვალებადობა იქმნებოდა. თანდათანობით, ზოგიერთ (თუმცა არა ყველა) შემთხვევაში ამ ცვალებადობის მატარებელი ორგანიზმები ერთმანეთისაგან მორფოლოგიურად განსხვავებულნი ხდებოდნენ და ერთმანეთთად შეჯვარების შემთხვევაში ნაყოფიერი შთამომავლობის წარმოქმნა აღარ შეეძლოთ, რაც ახალი სახეობის წარმოქმნის მიმანიშნებელია. ამ თეორიის მოწინააღმდეგეები მართებულად მიუთითებდნენ, რომ დარვინს ახალი სახეობის წარმოქმნის ერთი მაგალითიც კი არ მოუყვანია: ხელოვნური და ბუნებრივი გადარჩევის დეტალური შედარება მათთვის არასაკმარისად დამაჯერებელი გახლდათ. დარვინის მომხრეთაგან ზოგიერთიც კი არ იზიარებდა მის იდეას ცვალებადობისა და ევოლუციის უწყვეტობის შესახებ. მეტიც, დარვინის მომხრეთაგან ბევრს მიაჩნდა, რომ ბუნებრივი გადარჩევა ხდება „წყვეტილი“ ნახტომების შედეგად, რომლებიც მნიშვნელოვან (ასევე „წყვეტილ“ ცვალებადობაზეა დამოკიდებული). თვით დარვინმა, რასაკვირველია იცოდა მუტაციების არსებობის შესახებ („sports“, დარვინისდროინდელი ტერმინოლოგიით), მაგრამ მიაჩნდა, რომ ასეთი წარმონაქმნები მეტისმეტად იშვიათი იყო და ხშირად შთამომავლობის წარმოქმნაც არ შეეძლოთ, და ამიტომაც ცვალებადობის საფუძველს ვერ შექმნიდნენ.
ეს წინააღმდეგობა ორი შეხედულების („წყვეტილისა თუ უწყვეტი“ განვითარების) შორის ევოლუციის შესწავლის ისტორიაში ერთ-ერთი უმნიშვნელოვანესი თემაა და მეცნიერული დებატების მეტად მწვავე საკითხს წარმოადგენს, რომელსაც მომავალ წერილებში დავუბრუნდებით.
გელა თევზაძე.
24 საათი 31.08.11
|
|
16 ევოლუცია: დარვინის თეორია და მისი განვითარება. II. დარვინის იდეების გავლენა ევოლუციის კვლევაზე (ნაწილი II) |
▲ზევით დაბრუნება |
როგორც წინა სტატიაში აღვნიშნეთ, დარვინის „სახეობათა წარმოშობამ“ დაარწმუნა მსოფლიო ევოლუციის არსებობაში, მაგრამ დარვინისეული თეორია ბუნებრივი გადარჩევის შესახებ გაცილებით ნაკლები ერთსულოვნებით იყო მიღებული. ამ სტატიაში განვიხილავთ ევოლუციის შესწავლის შედეგად წარმოქმნილ სხვა იდეებს დარვინის თეორიასთან მიმართებაში.
ბუნებრივი გადარჩევის თეორიაზე დაყრდნობით, საკმაოდ ძნელი ასახსნელია ალტრუისტული სოციალური ქცევის ევოლუცია. „სახეობათა წარმოშობაში“ დარვინმა აღიარა ეს სიძნელე და ივარაუდა, რომ ამგვარი სოციალური ქცევის ევოლუციის შემთხვევაში ბუნებრივი გადარჩევა მოქმედებს მთლიან ოჯახებზე და არა სახეობის ცალკეულ წარმომადგენლებზე (ინდივიდუუმებზე). ბუნებრივი გადარჩევის თეორიის მოწინააღმდეგეთათვის ურთიერთშეთანხმებული ან ალტრუისტული სოციალური ქცევა დიდი ხნის განმავლობაში რჩებოდა იდეალურ არგუმენტად ინდივიდუალური გადარჩევისა და, მაშასადამე, ევოლუციის მთელი თეორიის გასაბათილებლად. მხოლოდ 1960-იან წლებში მოხერხდა ამ მოვლენის ახსნა ინდივიდუალური გადარჩევის მექანიზმების გამოყენებით: ალტრუისტული მოქმედების განმარტებისათვის უილიამ დონალდ ჰამილტონმა (William Donald Hamilton, 1936-2000) 1964 წელს განსაზღვრა მექანიზმი, რომელსაც „ნათესავთა გადარჩევა“ (kin selection) ეწოდა. თვით ტერმინი შემოღებულია 1964 წელსვე ჯონ მეინარდ სმიტის (John Maynard Smith, 1920-2004) მიერ, და აღნიშნავს იმგვარ ევოლუციურ სტრატეგიას, როდესაც ინდივიდუუმის ნათესავთა გადარჩევა (მათი გამრავლების უზრუნველყოფა) ხდება, თუნდაც თვით ინდივიდუუმის ხარჯზე. ასეთი გადარჩევის კლასიკურ მაგალითად მიჩნეულია მწერების (ფუტკრების ან ჭიანჭველების) კოლონია, რომელშიც უნაყოფო მდედრები („მუშები“) საკვებით მომარაგების გზით ხელს უწყობენ თავიანთ დედას („დედოფალს“) შთამომავლობის წარმოქმნაში. ამ გზით „მუშები“ გარკვეულწილად საკუთარი გენების შენარჩუნებასა და გამრავლებასაც უწყობენ ხელს, რადგან მათ და „დედოფალს“ ერთნაირი გენები მრავლად აქვთ.
ბუნებრივი გადარჩევისა და შეძენილი თვისებების მემკვიდრეობით გადაცემის გარდა, დარვინმა ბუნებრივი ევოლუციის მესამე მექანიზმის არსებობაც ივარაუდა. მას არ გასძნელებია, აეხსნა სქესობრივი გამრავლების როგორც წარმოშობა, ისევე შენარჩუნება, მაგრამ ამასთანავე დარვინმა ვერ შესძლო, ბუნებრივი გადარჩევის მოქმედების საფუძველზე აეხსნა სქესობრივი დიმორფიზმი (განსხვავება მდედრობითი და მამრობითი სქესის წარმომადგენლებს შორის) ან თუნდაც სხვაობა ადამიანთა რასებს შორის. დარვინის მიერ შემოთავაზებული „სქესობრივი გადარჩევა“ (sexual selection) წინააღმდეგობრივი იყო: ერთი მხრივ, ამ თეორიის მიხედვით, მდედრობითი სქესის ჩიტები უპირატესობას აძლევდნენ (ირჩევდნენ) მკვეთრად შეფერილ მამრებს, და ასეთი მამრების შთამომავლობა ასევე მკვეთრი შეფერილობით ხასიათდებოდა. მაგრამ ასეთი „გადარჩევა“ არ შეესაბამება ბუნებრივი გადარჩევის საფუძვლებს (მეტიც, მას ეწინააღმდეგება), რადგან მკვეთრად შეფერილი ჩიტები უფრო იოლად ხდებიან მტაცებლების მსხვერპლი.
დარვინის თეორიის კრიტიკოსები მეცხრამეტე საუკენეში. 1859-1872 წლებში გამოქვეყნდა დარვინის არაერთი მნიშვნელოვანი ნაშრომი, მათ შორის „სახეობათა წარმოშობის შესახებ“ („On the Origin of Species“, 1859), „ორქიდეათა განაყოფიერება“ („Fertilization of Orchids“, 1862), „ცხოველთა და მცენარეთა ცვალებადობა მოშინაურების შედეგად“ („Variations of Plants and Animals under Domestication“, 1868), „ადამიანის წარმოშობა“ („Descent of Man“, 1871), „ემოციების გამოხატვა ადამიანსა და ცხოველებში“ („Expression of Emotions in Man and Animals“, 1872). ამ შრომებმა კი, თავის მხრივ, გამოიწვია დიდძალი ლიტერატურის დაგროვება ევოლუციის თემაზე. 1872 წელს გამოქვეყნდა დარვინიზმის ბიბლიოგრაფიის მეორე გამოცემა, რომელიც 36 გვერდს მოიცავდა და 317 ავტორს ასახელებდა, რომელთაგან მრავალი რამდენიმე ნაშრომის ავტორი იყო. ბიბლიოგრაფია გერმანულად იყო გამოცემული და, ბუნებრივია, გერმანულენოვან ლიტერატურას ანიჭებდა უპირატესობას. ამის გამო, ბიბლიოგრაფია არასრული იყო - მაგალითად, ის ერთ რუსულენოვან ნაშრომსაც კი არ მოიხსენიებდა. ამ ბიბლიოგრაფიის გამოცემიდან სამი წლის შემდეგ (1875) დაბეჭდილი ანალოგიური გამოცემა უკვე ორჯერ მეტ დასახელებას შეიცავდა. 1900 წლისთვის კი ყველა იმ ნაშრომის აღნუსხვა, სადაც დარვინი ან მისი შრომები იყო ნახსენები, უკვე შეუძლებელი გახდა. ამ საკითხებზე დაგროვილი ლიტერატურა, რასაკვირველია, იმაზე მეტყველებს, რომ დარვინმა შესძლო დაერწმუნებინა არა მარტო მეცნიერები, არამედ ფილოსოფოსებიცა და თეოლოგებიც ევოლუციის არსებობაში. მაგრამ იგივე ლიტერატურა იმასაც მოწმობს, რომ ბუნებრივი და სქესობრივი გადარჩევის დარვინისეული თეორიების ყოველი დეტალი მწვავე კრიტიკის ქარცეცხლში გაატარეს.
არსებობს მოსაზრება, რომ 1882 წლისათვის (როდესაც დარვინი გარდაიცვალა) ცნობილ მეცნიერთაგან მხოლოდ ერთი მესამედი აღიარებდა ბუნებრივ გადარჩევას, როგორც ევოლუციის უმთავრეს ძალას, მაშინ როცა დარჩენილი ორი მესამედი მიიჩნევდა, რომ ევოლუციას სხვა მექანიზმები წარმართავდნენ. ასევე მრავლად იყვნენ ისეთნი, ვინც „წყვეტილ“ („ნახტომისებრ“) ევოლუციას ანიჭებდა უპირატესობას, დარვინისეული „უწყვეტი განვითარების“ საწინააღმდეგოდ. საუკუნის დასასრულისათვის, დარვინის თეორიამ ევოლუციის შესახებ, რომლის მიხედვითაც ევოლუცია უწყვეტი ბუნებრივი გადარჩევით წარიმართებოდა, მრავალი მოწინააღმდეგე შეიძინა, და მოწინააღმდეგეთა შორის ბევრი საქვეყნოდ ცნობილი ბიოლოგი იყო.
თუმცა უნდა აღინიშნოს, რომ ამ ოპონენტთა შორის სულ ცოტა ორი ჯგუფი არსებობდა. ერთნი უპირატესობას ანიჭებდნენ ალტერნატიულ „მექანისტურ“ თეორიებს, მაგალითად, მორიც ვაგნერის (Moritz Wagner, 1813-1887) მიგრაციის თეორიას ან ჰუგო დე ფრიზის (Hugo de Vries, 1848-1935, მენდელის კანონების ხელმეორედ აღმომჩენი) მუტაციის თეორიას. ასეთნი ოპოზიციის მცირე ნაწილს შეადგენდნენ, უმრავლესობა კი დარვინს აკრიტიკებდა და ევოლუციის სხვაგვარ მექანიზმს ეძებდა იმის იმედით, რომ მათი შემოთავაზებული ახსნა ევოლუციის მიზანსა და წინასწარ დაგეგმილ მიმართულებას წარმოაჩენდა. მათი წარმოდგენით, ბუნების გეგმაზომიერების იდეის უარყოფის გზა ადამიანის სიცოცხლის მნიშვნელობის უარყოფამდე მიდიოდა. ზოგადად, მეცხრამეტე საუკუნის დასასრულისათვის დიდძალი ლიტერატურაა დაგროვილი, რომელშიც აშკარად სჩანს ევოლუციის შერიგების მცდელობა ქრისტიანულ თეოლოგის (და ზოგადად რელიგიის) დოგმებთან. როგორც ზემოთქმული ცხადჰყოფს, დარვინმა, მთელი მისი გავლენის მიუხედავად, ვერ შეძლო ერთიანი აზრის შექმნა ევოლუციის მექანიზმების შესახებ.

ნახ. 1: ჰუგო დე ფრიზი (1848 - 1935).
მენდელისეული გენეტიკის აღზევება. 1930-1940 წლებში წარმოქმნილი „ევოლუციური სინთეზის“ (იხ. ქვემოთ) საფუძველზე შესაძლოა ვივარაუდოთ, რომ მეოცე საუკუნის დასაწყისისათვის დარვინიზმის ერთადერთი ნაკლი მემკვიდრეობითობის თეორიის არარსებობა გახლდათ. როდესაც ეს ნაკლი შეივსო მენდელის თეორიის ხელმეორედ აღმოჩენის გამო, შესაძლებელი გახდა მენდელიზმისა და დარვინიზმის სინთეზი. ამ ორი თეორიის შერწყმა საფუძვლად დაედო ნეო-დარვინიზმს, მაგრამ თვით სინთეზი არც ადვილი გახლდათ და არც უეცრად მომხდარა მენდელიზმის ხელმეორედ აღმოჩენის შემდეგ.
1900 წლისათვის დისკუსია უწყვეტი და წყვეტილი ევოლუციის მომხრეთა შორის უკიდურესად გამწვავდა. წყვეტილი ევოლუციის მომხრეთა შორის იყვნენ უილიამ ბაიტსონი (William Bateson, 1861-1926), ფრანსის გალტონი (Francis Galton, 1822-1911), ჰუგო დე ფრიზი. უწყვეტი ევოლუციის თეორიის მომხრეები ე.ბ. პოლტონი (E.B. Poulton, 1856-1943) და ი. რეი ლანკასტერი (E. Ray Lancaster, 1847-1929), ისევე როგორც ბიომეტრიის სპეციალისტები კარლ პირსონი (Karl Pearson, 1857-1936) და უ.ფ.რ. უელდონი (W.F.R. Weldon, 1860-1906). ადვილი მისახვედრია, რომ დე ფრიზი წყვეტილ ევოლუციას ემხრობოდა მუტაციების (უეცარი, „თვისობრივი“ ცვლილებების) საფუძველზე, ხოლო ბიომეტრიის დარგში მოღვაწე მეცნიერები თვლიდნენ, რომ რაოდენობრივი ნიშან-თვისებები (სიმაღლე, სიგრძე, წონა და ა.შ.), რომლებსაც ბიომეტრია შეისწავლის და რომლებიც „თანდათანობითი ცვლილების“ მაგალითია, სწორედ უწყვეტი ევოლუციის მაგალითი გახლდათ. ასევე ადვილი ასახსნელია, რომ მენდელისეული კანონები წყვეტილი ევოლუციის მხარდამჭერ არგუმენტად იქნა მიჩნეული: მენდელი თავის ექსპერიმენტში იყენებდა მკვეთრად განსხვავებულ ნიშან-თვისებებს, მაგ., მარცვლების მწვანე და ყვითელი ფერი, თეთრი და იასამნისფერი ყვავილები), და მათ შორის გარდამავალი ფორმები არ არსებობდა (მაგალითად, მომწვანო-მოყვითალო, ან ცისფერი ყვავილები. ამავე მიზეზით აიხსნება მენდელიზმის კრიტიკა უწყვეტი ევოლუციის მომხრეების მიერ. ზოგადად, დისკუსიის ამ პერიოდში (1900-1906) დარვინიზმის წარუმატებლობა გარკვეულწილად მენდელიზმის წარმატების შედეგი იყო.
ჰუგო დე ფრიზი წყვეტილი ევოლუციის ყველაზე გავლენიანი მომხრე გახლდათ. 1899 წელს დე ფრიზმა აღმოაჩინა წყვეტილი მუტაციების მრავალი მაგალითი საღამოს ფურისულაში (Oenothera). ამ მცენარეთაგან ზოგიერთი მშობლებთან შეჯვარებისას შთამომავლობას არ იძლეოდა, ამიტომ დე ფრიზმა დაასკვნა, რომ ეს მოვლენა ახალი სახეობის წარმოქმნის მიმანიშნებელი იყო. 1901-1903 წლებშ დე ფრიზმა გამოაქვეყნა ორტომეული „მუტაციების თეორია“ („Mutations theorie“), რომელშიც წყვეტილი ევოლუციის მრავალი მხარდამჭერი საბუთი ჩამოვთვალა. დე ფრიზის პოპულარობის ერთ-ერთი მიზეზი მეცხრამეტე საუკუნის დასასრულს ექსპერიმენტული ბიოლოგიის განვითარებაა; თანაც, რაც მეტად შთამბეჭდავი იყო, დარვინისაგან განსხვავებით, დე ფრიზმა თავისი თეორიის გარკვეული ექსპერიმენტული დასაბუთებაც წარადგინა.
1906 წლისათვის შეიქმნა შთაბეჭდილება, რომ მენდელიზმისა და მუტაციების თეორიის კავშირის გამარჯვება დარვინისეულ უწყვეტ ევოლუციაზე გარდაუვალი იყო. მაგრამ „არადარვინისეული“ თეორიების მთავარი პრობლემა ზუსტად იგივე იყო, რაც დარვინისეული ბუნებრივი გადარჩევისათვის: მისი მხარდამჭერი საბუთების ნაკლებობა. დარვინისეული იდეები ხელახლა გახდა მიმზიდველი, როდესაც გამოირკვა, რომ მუტაციების თეორიის საფუძველზე ყველაფერი ვერ აიხსნებოდა, და რომ პოპულაციების (ერთი ცალკეული სახეობის წარმომადგენლების ერთობლიობის) შეცვლისათვის უწყვეტი, ანუ თანდათანობითი გადარჩევა საკმაოდ ეფექტური აღმოჩნდა.
მენდელიზმისა და დარვინიზმის სინთეზი. მეოცე საუკუნის პირველ ათწლეულში გაბატონებული იყო შეხედულება, რომ გადარჩევა მოქმედებს მხოლოდ იმ ფენოტიპების (გარეგნული ნიშან-თვისებების) ფარგლებში, რომლებიც უკვე არსებობენ პოპულაციაში. ამ თეორიის მხარდასაჭერად, ფრანსის გალტონი ამტკიცებდა, რომ „რეგრესია“ რამდენიმე ხანში გააწონასწორებს გადარჩევას, თუ ამ პროცესში ახალი მუტაცია არ წარმოიქმნება. გენეტიკურად წმინდა „ჯიშებში“ გადარჩევა არაეფექტურია, ხოლო რეგრესია სრულყოფილი (ანუ დასრულებული).
ამ შეხედულების საფუძველზე წარმოიქმნა მეტად საინტერესო ტენდენცია, რომელიც ექსპერიმენტულ მეცნიერებაში არცთუ იშვიათად შეინიშნება: გენეტიკოსები, რომლების მიზანს წარმოადგენდა მუტაციების თეორიის მხარდასაჭერი (და დარვინიზმის საწინააღმდეგო) ექსპერიმენტული შედეგების მიღება, ამ შედეგების საფუძველზე მეტად განსაცვიფრებელი (უპირველეს ყოვლისა, საკუთარი თავისათვის) დასკვნები გამოიტანეს: მათ აღმოაჩინეს, რომ მენდელიზმი და დარვინიზმი არა მარტო არ ეწინააღმდეგება ერთმანეთს, არამედ სავსებით ურთიერთშეთანხმებადია. ამერიკელი გენეტიკოსი უილიამ კასლი (William Castle, 1867-1962) თავისი ექსპერიმენტებში იკვლევდა გადარჩევის გავლენას ვირთხების ზურგზე არსებული შეფერილი ზოლის ცვალებადობაზე. კასლმა აღმოაჩინა, რომ ზოლის შეფერილობაში ისეთი მრავალფეროვნება წარმოიქმნა, რომელიც ექსპერიმენტის დასაწყისში ამ პოპულაციის ფარგლებში არ შეიმჩნეოდა. ამრიგად, კასლის ცდებში გადარჩევა მცირე ცვალებადობების მიმართულებით წარმატებული აღმოჩნდა.

ნახ. 2: ვირთხების ზურგზე არსებული ზოლის ფორმისა და შეფერილობის რამდენიმე მაგალითი (უილიამ კასლის ექსპერიმენტებიდან).
ედუარდ ისტი (Edward M. East, 1879-1938) და ჰერმან ნილსონ-ელე (Herman Nilsson-Ehle, 183-1949) იმავდროულად ავითარებდნენ მრავალი ფაქტორის თეორიას (multiple-factor theory), რომელიც უწყვეტ ცვალებადობას მენდელისეული მემკვიდრეობის საფუძველზე განმარტავდა. 1918 წელს აღმოჩნდა, რომ კასლის ექსპერიმენტების შედეგები დროზოფილას შემთხვევაშიც მართლდებოდა. მეტიც, გარკვეული ნიშან-თვისებების განმსაზღვრელი ფაქტორების მემკვიდრეობით გადაცემა მენდელისეული კანონების თანახმად ხდებოდა. ამერიკელი გენეტიკოსი ალფრედ სტერტევანტი (Alfred H. Sturtevant, 1891-1970) თავის ექსპერიმენტებში, სადაც კვლევის ობიექტი დროზოფილა იყო კიდევ უფრო წინ წავიდა და ისიც აჩვენა, თუ რომელ ქრომოსომებზე იყვნენ განლაგებული ეს ფაქტორები. სტერტენავტს ასევე ეკუთვნის პირველი „ქრომოსომული რუკა“ - გენების განლაგების თანმიმდევრობის განსაზღვრა ქრომოსომაზე (1913). შემდგომ აღმოჩნდა, რომ გენეტიკური რეკომბინაციაც (ქრომოსომებს შორის ფრაგმენტებისა და, მაშასადამე, გენების მიმოცვლა) მენდელის კანონების თანახმად ხდება. რეკომბინაციის აღმოჩენის შედეგად კი ნათელი გახდა, რომ 10 სხვადასხვა გენიც კი, რომელთაგან თითოეული ორი განსხვავებული ვარიანტის (ალელის) სახით არსებობს, წარმოქმნის ~60 ათას სხვადასხვა კომბინაციას (გენოტიპს, ანუ გენების ერთობლიობას). გავითვალისწინოთ, რომ სინამდვილეში ორგანიზმები არა 10, არამედ 3 000-დან (ბაქტერია) 25 000-მდე (ადამიანი) გენს შეიცავენ. ასე რომ, ნებისმიერი პოპულაცია, რომელშიც თუნდაც მცირე ცვალებადობა არსებობს, მოცემულ დროში ამ ცვალებადობის ძალიან მცირე ნაწილს ავლენს. გადარჩევის შედეგად კი ნაკლები სიხშირით არსებული გენოტიპების სიხშირე იმატებს. ასე რომ, მენდელისეული მემკვიდრეობითობა და დარვინისეული გადარჩევა არათუ არ ეწინააღმდეგებიან, არამედ ავსებენ ერთმანეთს. 1918 წლისათვის, როდესაც ცნობილი გახდა, რომ ჰუგო დე ფრიზის მუტაციების უნაყოფობა მშობლებთან შეჯვარებისას სინამდვილეში „წმინდა ხაზების“ დაბალი სიცოცხლისუნარიანობით აიხსნებოდა, ექსპერიმენტულ გენეტიკაში მოღვაწე მეცნიერთაგან უმეტესობას სჯეროდა, რომ მენდელიზმისა და დარვინიზმის სინთეზი შესაძლებელია.
ნახ. 3: გენეტიკური რუკის ფრაგმენტი (დროზოფილის მაგალითზე). ნაჩვენებია გენების წრფივი განლაგება ქრომოსომაზე, მანძილი მათ შორის, და ნიშან-თვისებები, რომლებსაც ეს გენები განსაზღვრავენ (ფეხების სიგრძე, სხეულის ან თვალების ფერი, ფრთების ფორმა და ა.შ.).
მენდელისეული მემკვიდრეობითობა ნიშან-თვისებათა გადაცემის მწყობრი სისტემაა, და მის საფუძველზე შესაძლოა მემკვიდრეობითობის რაოდენობრივი შედეგების გამოთვლა. ასეთი გამოთვლებიდან უმარტივესია ჰარდი-ვაინბერგის კანონი, რომელიც ამტკიცებს, რომ პოპულაციაში ერთი გენის ორი ვარიანტის (ალელის) არსებობის შემთხვევაში, ამ ალელების სიხშირეების სტაბილური წონასწორობის მიღწევა შესაძლებელია ერთი თაობის განმავლობაში თავისუფალი შეჯვარების შედეგად. 1920 წლისათვის, პოპულაციებში ალელების სიხშირის გამოთვლის რამდენიმე რაოდენობრივი მოდელი იყო შემუშავებული. ყველა ეს მოდელი დაფუძნებული იყო იმ მოსაზრებაზე, რომ მენდელიზმი ავსებდა და სრულჰყოფდა დარვინისეულ თეორიას ბუნებრივი გადარჩევის გზით მიმავალი ევოლუციის შესახებ. ამ მოდელების არსს, ისევე როგორც დარვინის თეორიის შემდგომ განვითარებასა და ე.წ. „ევოლუციურ სინთეზს“, მომდევნო სტატიაში განვიხილავთ.
გელა თევზაძე.
24 საათი 18.09.11
|
|
17 ევოლუცია: დარვინის თეორია და მისი განვითარება. II. დარვინის იდეების გავლენა ევოლუციის კვლევაზე (ნაწილი III). |
▲ზევით დაბრუნება |
მენდელის დადგენილი (და ჰუგო დე ფრიზის მიერ ხელმეორედ აღმოჩენილი) კანონების საფუძველზე შესაძლებელია რომელიმე ნიშან-თვისების განმსაზღვრელი გენის სხვადასხვა ვარიანტების (ალელების) სიხშირეების გამოთვლა პოპულაციაში, ანუ ერთი სახეობის ინდივიდუუმთა ერთობლიობაში. ასეთი გამოთვლების საფუძველია ჰარდი-ვაინბერგის კანონი, რომლის თანახმად ერთი გენის ორი ალელის სიხშირეების განაწილება პოპულაციაში მარტივი მათემატიკური მოდელით (კვადრატული განტოლებით) გამოისახება. თავისუფალი შეჯვარების პირობებში, ანუ როდესაც არცერთ ალელს არა აქვს სელექციური უპირატესობა, მათი სიხშირეების წონასწორობა პირველივე თაობის განმავლობაში იქმნება. თანაც, ნაკლები სიხშირით არსებული ალელის შენარჩუნებას სწორედ ის ორგანიზმები უწყობენ ხელს, რომლების გენოტიპში ორივე ალელია „მოთავსებული“. მაგალითად, თუ ყვავილების ფერის განმსაზღვრელ გენს ორი ვარიანტი (ალელი) აქვს - თეთრი და იასამნისფერი, და ამათგან თეთრია დომინანტური, ორგანიზმში ორივე ალელის არსებობის შემთხვევაში ყვავილები თეთრი ფერის იქნება. ამრიგად პოპულაციაში არსებულ მცენარეთა უმრავლესობას თეთრი ყვავილები ექნება, მაგრამ „იასამნისფერი ალელი“ მაინც შენარჩუნდება (როგორც იასამნისფერ, ასევე იმ ყვავილებში, რომელნიც ორივე ალელს ატარებენ) და გადაეცემა მომდევნო თაობებს. ალელების სიხშირეების გამოსათვლელი მათემატიკური მოდელები არსებობს იმ შემთხვევებისთვისაც, როდესაც ერთ-ერთ ალელს სელექციური უპირატესობა აქვს ან როცა გარკვეული გენოტიპების ორგანიზმებს შორის შეჯვარება მეტი სიხშირით ხდება დანარჩენებთან შედარებით.
ამგვარმა რაოდენობრივმა მოდელებმა დიდი გავლენა მოახდინეს ევოლუციის შემსწავლელ მეცნიერებსა და მათი კვლევის მიმართულებების განსაზღვრაზე. ეს გავლენა რამდენიმენაირად გამოვლინდა: 1) ცხადი გახდა, რომ მენდელის კანონებით განსაზღვრული მემკვიდრეობითობა და ბუნებრივი გადარჩევა, პოპულაციებში მიმდინარე პროცესებთან ერთად, საკმარისია ბუნებაში არსებული ევოლუციის მექანიზმების ასახსნელად. ამ მოდელებმა დაადასტურეს დარვინის ვარაუდი იმის შესახებ, რომ გადარჩევას, თუნდაც ის დაბალი სიჩქარით მიმდინარეობდეს, შეუძლია პოპულაციის გენეტიკური შემადგენლობის შეცვლა. 2) ამავე მოდელების საფუძველზე გაირკვა, რომ თავისთავად მუტაციების წარმოქმნა ვერ იქნება ევოლუციის წამმართველი ძალა, და რომ გადარჩევა ამ თვალსაზრისით გაცილებით უფრო ეფექტურია, ვიდრე მხოლოდ მუტაციები. ეს მოსაზრებაც მეტად მნიშვნელოვანია, რადგან მან გააბათილა წარმოდგენა, რომ „მუტაციური წნეხი“ უმთავრესი ფაქტორი იყო ევოლუციური ცვლილებების წარმოქმნაში. ბუნებრივი გადარჩევის („სელექციური წნეხის“) ეს უპირატესობა მუტაციებთან შედარებით მათემატიკური მეთოდებით იქნა დადგენილი, მუტაციების წარმოქმნის ტემპის, სელექციური წნეხისა და გენების სიხშირეების ცვლილებების ურთიერთკავშირის გამოაშკარავების საფუძველზე. 3) მათემატიკური მოდელები გახდა ექსპერიმენტული შედეგების შემავსებელი და განმავრცობელი ნაწილი, რითაც თვით ექსპერიმენტული კვლევის მნიშვნელობა საგრძნობლად გაიზარდა. მაგალითად, 1924 წელს ჩრჩილის ერთ-ერთი სახეობის შეფერილობის კვლევისას მათემატიკური გამოთვლებით დადგინდა, რომ ინგლისის იმ რაიონებში,სადაც მრეწველობის განვითარების შედეგად ჭვარტლის რაოდენობა გაიზარდა, მუქი შეფერილობის მწერების გადარჩენის (ანუ გადარჩევისაც) ალბათობა ორჯერ მეტი იყო, ვიდრე მრეწველობის განვითარებამდე გავრცელებული ღია ფერის ორგანიზმებისა. ეს გამოთვლები ექსპერიმენტულადაც დადასტურდა, და ბუნებრივი გადარჩევის პირველი თვალსაჩინო (თანაც „ორგზის დასაბუთებული“) მაგალითი გახდა - დარვინის გარდაცვალებიდან 65 წლის შემდეგ... 4) და ბოლოს, მათემატიკურმა მოდელებმა ხელი შეუწყო ექსპერიმენტული კვლევების განვითარებას, რაც თვით ამ მოდელების შემოწმებისა და შემდგომი დახვეწისათვის იყო ხელსაყრელი. ამ გარემოების საუკეთესო მაგალითია თეოდოსიუს დობჟანსკის (Theodosius Dobzhansky, 1900-1975) სტატიების შთამბეჭდავი ერთობლიობა - 43 ნაშრომი დროზოფილის გენეტიკური კვლევის თემაზე, რომლებიც მისი გარდაცვალების შემდეგ, 1981 წელს, გამოქვეყნდა ერთობლივი სათაურით „ბუნებრივი პოპულაციების გენეტიკა“ („The Genetics of Natural Populations“). [დობჟანსკის ასევე ეკუთვნის სამეცნიერო-პოპულარული სტატია „ევოლუციური თვალთახედვის გარეშე, ბიოლოგიაში ყველაფერი უაზროა“, რომლის თარგმანს მომავალში შემოგთავაზებთ].

ნახ. 1:. ღია და მუქი შეფერილობის ჩრჩილები ჭვარტლიან („განვითარებული მრეწველობის“) ფონზე.
ევოლუციური სინთეზი. პოპულაციურ გენეტიკაში მოღვაწე მეცნიერთა მათემატიკურ ნაშრომები, მათი დიდი მნიშვნელობის მიუხედავად, საკმარისი არ აღმოჩნდა ბუნებრივი ევოლუციის „სინთეტური“ ახსნისათვის,რაც მხოლოდ 1930-1940-იან წლებში მოხდა. ბიოლოგიური კვლევის სხვადასხვა - მეცხრამეტე საუკუნის ბოლოსათვის ერთმანეთისაგან უკვე საკმაოდ დაშორებული მიმართულება - კვლავ გაერთიანდა ევოლუციური სინთეზის თეორიაში. ამ დროისათვის ბიოლოგებისათვის ნათელი გახდა, რომ გენეტიკა, სისტემატიკა, ციტოლოგია, პალეონტოლოგია და ემბრიოლოგია, კლასიკური ბოტანიკისა და ზოოლოგიის დამატებით, ერთიანი ევოლუციური პროცესის ურთიერთშემავსებელი ნაწილები არიან. ამგვარი „სინთეტური მიდგომის“ შესანიშნავი მაგალითებია უკვე ნახსენები „ბუნებრივი პოპულაციების გენეტიკა“ დობჟანსკის ავტორობით; ჯულიან ჰაქსლის „ახალი სისტემატიკა“ და „ევოლუცია: თანამედროვე სინთეზი“ (Julian Huxley, „The New Systematics“, 1940; „Evolution: The Modern Synthesis“, 1942), ერნსტ მეირის „სისტემატიკა და სახეობათა წარმოშობა“ (Ernst Mayr, „Systematics and the Origin of Species“, 1942), ჯორჯ სიმფსონის „ტემპი და მიმართულება ევოლუციაში“ (George G. Simpson, „Tempo and Mode in Evolution“, 1944), და ლედიარდ ს „ცვალებადობა და ევოლუცია მცენარეებში“ (Ledyard Stebbins, „Variation and Evolution in Plants“, 1950).

ნახ. 2: , სიმფსონი და დობჟანსკი 1959 წელს ჩიკაგოში ჩატარებულ კონფერენციაზე, რომელზეც ევოლუციური სინთეზის საკითხები განიხილებოდა.
ევოლუციური სინთეზის საფუძველზე, შესაძლებელი გახდა ნატურალისტური (ბუნებაზე დაკვირვების შედეგად მიღებული), ექსპერიმენტული (გენეტიკური ცდებიდან გამომდინარე დასკვნები) და პალეონტოლოგიური ცოდნის გაერთიანება. რაში მდგომარეობს ევოლუციური სინთეზის ძირითადი დებულებები?
ყველა ევოლუციური მოვლენის ახსნა შესაძლებელია არსებული გენეტიკური მექანიზმებისა და ნატურალისტური კვლევის საფუძველზე.
ევოლუცია თანდათანობით (უწყვეტად) მიმდინარეობს: მცირე გენეტიკური ცვლილებები და რეკომბინაცია იმართება ბუნებრივი გადარჩევით. სხვაობა („წყვეტილი განსხვავება“, discontinuities) სახეობებსა თუ სხვა ტაქსონომიურ ერთეულებს შორის აიხსნება თანდათანობითი („უწყვეტი“) ცვლილებების დაგროვებით, გეოგრაფიული განცალკევებისა და ზოგიერთი ფორმის გადაშენების გზით.
ბუნებრივი გადარჩევა ცვალებადობის უმთავრესი მექანიზმია, რადგან მცირე უპირატესობაც კი მეტად მნიშვნელოვანია დროის განმავლობაში. გადარჩევა მოქმედებს როგორც ფენოტიპზე (გარეგნულ ნიშან-თვისებებზე), ასევე მის გარემო პირობებზე.
გენეტიკური დრეიფის (ცალკეული გენების სიხშირის უეცარი ცვლილების) მნიშვნელობა დადგენილი არაა (დობჟანსკი ამ პროცესს ანგარიშგასაწევ ევოლუციურ მექანიზმად თვლიდა, მაგრამ მოგვიანებით ეკოლოგიურ გენეტიკაში ამ მოსაზრების საწინააღმდეგო შედეგებიც დაგროვდა).
მნიშვნელოვანია პოპულაციების, და არა ინდივიდუუმების ტერმინებით ოპერირება: ბუნებრივ პოპულაციაში არსებული გენეტიკური მრავალფეროვნება გადამწყვეტია ევოლუციისათვის. ბუნებრივ პირობებში გადარჩევა ძალიან ეფექტურია, დიდი როლი ენიჭებათ ეკოლოგიურ ფაქტორებსაც, მაგ., ეკოლოგიური ნიშებს, გენების გავრცელების დამაბრკოლებელ გეოგრაფიულ ბარიერებს და ა.შ.
პალეონტოლოგიური მონაცემების ახსნა შეიძლება მიკროევოლუციის (სახეობათა ფარგლებში არსებული ცვალებადობის) მექანიზმების ექსტრაპოლაციის გზით მაკროევოლუციის (სახეობათა და უფრო მაღალი სისტემატური ერთეულების წარმოქმნის) პროცესებზე. აქვე უნდა აღინიშნოს, რომ „თანდათანობითი“, „უწყვეტი“ ცვლილება არ ნიშნავს, რომ ამ ცვლილების ტემპი მუდმივია.
1950-იანი წლების დასაწყისისათვის, ევოლუციურმა სინთეზმა უაღრესად დახვეწილი თეორიის სახე მიიღო და უფრო დამაჯერებელ მოძღვრებად იყო მიჩნეული, ვიდრე ნებისმიერი წინანდელი თეორია ევოლუციის შესახებ. ცხადი გახდა, რომ ბუნებრივი ევოლუცია უპირატესად მიმდინარეობდა მცირე სელექციური წნეხის მექანიზმების საფუძველზე, და ამ პროცესში გენეტიკური მრავალფეროვნებას მნიშვნელოვანი როლი ჰქონდა. რაც არანაკლებ საყურადღებოა, ევოლუციის მექანიზმების ეს ნიშან-თვისებები დარვინის შეხედულებებს ემთხვეოდა, ამიტომაც 1959 წელს, როცა „სახეობათა წარმოშობას“ საუკუნე შეუსრულდა, ევოლუციური სინთეზის თეორია ხშირად (და სავსებით მართებულად) მოიხსენიებოდა, როგორც „მეოცე საუკუნის დარვინიზმი“ ან „ნეოდარვინიზმი“. ამ „ახალი“ დარვინიზმის ძირითადი განსხვავება „ძველისაგან“ იყო მენდელის კანონებისა და ექსპერიმენტული შედეგების დამატება, ისევე როგორც ბიოლოგების დიდი უმრავლესობის მკაფიოდ გამოხატული მხარდაჭერა, რაც დარვინიზმის კლასიკური თეორიის შევსებისა და განვითარების შედეგად მოხდა.

ნახ. 3: მოტოო კიმურა (1924-1994).
დარვინიზმი ორი იუბილეს შორის (1959-1982 - ასი წლისთავი, შესაბამისად, „სახეობათა წარმოშობის“ გამოქვეყნებიდან და დარვინის გარდაცვალებიდან): თითქმის მაშინვე, როგორც კი ნეოდარვინიზმი საყოველთაოდ აღიარეს დასრულებულ თეორიად, ახალი წინააღმდეგობები (და, მაშასადამე, განვითარების ახალი შესაძლებლობა) წარმოიქმნა. ბუნებრივ პოპულაციებში აღმოჩნდა გენეტიკური ცვალებადობის უდიდესი (და აქამდე შეუსწავლელი) რაოდენობა. არის თუ არა გენეტიკური ცვალებადობა „შემგუებლური“ (adaptive)? - ასეთი იყო მთავარი კითხვა, რომლის ერთ-ერთი საფუძველი დროზოფილას ფერმენტების მრავალფეროვნების გამოკვლევა იყო. მოტოო კიმურა (1924-1994) თვლიდა, რომ ცვალებადობის უმეტესობა სელექციის თვალსაზრისით ნეიტრალურია, და რომ გარკვეული ნიშან-თვისებების ერთობლიობა უფრო ალბათობის საფუძველზე წარმოიქმნება, ვიდრე შეგუებული ფორმების გადარჩევის (adaptive selection) შედეგად. სტივენ გოლდისა და ნაილს ელდრიჯის (Stephen Jay Gould, Niles Eldridge) თეორია წერტილოვანი წონასწორობის (punctuated equilibria) შესახებ უარყოფდა ნეო-დარვინისტულ მოსაზრებას, რომ მაკროევოლუცია (სახეობათა წარმოშობა) მიკროევოლუციის (სახეობათა შიგნით არსებული ცვალებადობის გადარჩევის) ბუნებრივი გაგრძელებაა. გოლდისა და ელდრიჯის აზრით, ნამარხი ფორმების „წყვეტილი“ ჯაჭვი აიხსნებოდა არა იმით, რომ აღმოჩენილი ფორმების შორის არსებული ყველა გარდამავალი ორგანიზმი ჯერ არ იყო აღმოჩენილი, არამედ იმ გარემოებით, რომ სახეობათა წარმოშობა მართლაც „წყვეტილად“, ნახტომისებურად ხდებოდა, და არა თანდათანობით, როგორც ნეოდარვინიზმის მიმდევრები ვარაუდობდნენ. ნეოდარვინისტების აზრით კი, ის სრულყოფილი გეოლოგიური მონაცემები, რომლებიც ზოგიერთი სახეობის შემთხვევაში მართლაც არსებობს, სწორედ „უწყვეტი“, თანდათანობით მიმავალი ევოლუციური პროცესის მიმანიშნებელი იყო.
წყვეტილი თუ უწყვეტი ევოლუციის მომხრეთა დაპირისპირება ნაწილობრივ აიხსნება ორივე ბანაკის წარმომადგენელთა სურვილით, ევოლუციაში არსებულ ყველა მოვლენას ერთნაირად მარტივი და არაორაზროვანი ახსნა მოუძებნონ. გოლდისა და ელდრიჯის „წერტილოვანი წონასწორობის“ თეორია უკეთ განმარტავს ნამარხი ფორმების არასრულ სიას, რადგან ორგანიზმების „წყვეტილი“ ჯაჭვის არსებობის საუკეთესო (და უმარტივესი) ახსნა სწორედ წყვეტილი ევოლუციაა. ნეო-დარვინისტები, თავის მხრივ, ამტკიცებენ, რომ ბუნებრივ პოპულაციებში მოქმედი „უწყვეტი“ ევოლუცია სახეობათა წარმოშობის დონეზეც გრძელდება, და რომ ევოლუციურ სინთეზს (ანუ ნეოდარვინიზმს) ამ პროცესების სრულყოფილად ახსნა ძალუძს. ამასთანავე, ორივე მხარისათვის აუცილებელი ხდება მრავალი დამატებითი ჰიპოთეზის შემოღება „მოწინააღმდეგეთა მომხრე მოვლენების“ ასახსნელად (ანუ „წერტილოვანი წონასწორობისათვის“ - ბუნებრივ პოპულაციებში მოქმედი უწყვეტი გადარჩევისა, ხოლო ნეოდარვინისტებისათვის - ნამარხი ფორმების წყვეტილი ჯაჭვის ახსნა). უნდა ვაღიაროთ, რომ აღმოჩენილი ნამარხი ფორმები (ე.წ. „გეოლოგიური საბუთები“), მთელი თავისი მრავალფეროვნების მიუხედავად, ვერ გამორიცხავს ვერც წყვეტილი და ვერც უწყვეტი ევოლუციური მექანიზმების არსებობას. ამ ორი თეორიიდან ერთ-ერთისათვის უეჭველი და საბოლოო უპირატესობის მინიჭება მხოლოდ მაშინ გახდება შესაძლებელი, როდესაც ბუნებრივი ევოლუციის პროცესების შესახებ ზუსტი და ყოვლისმომცველი ცოდნა დაგვიგროვდება. არაა გამორიცხული, რომ მწვავე კამათი უწყვეტი თუ წყვეტილი ევოლუციის შესახებ, რომელიც დარვინის დროს დაიწყო და აწმყოშიც მიმდინარეობს, კვლავაც გაგრძელდება დარვინის გარდაცვალების ორასი წლისთავისათვის, 2082 წელს.
და ბოლოს, თუმცა ევოლუციურ ბიოლოგიაში მოღვაწე მეცნიერთა შორის კვლავაც გრძელდება კამათი, თუ რა პროცესებია პირველადი თუ მეორეხარისხოვანი ბუნებრივი ევოლუციის მიმდინარეობისას, ყველა მათგანი ემხრობა დარვინის აზრს იმის შესახებ, რომ ბუნებაში არსებული მრავალფეროვნება მხოლოდ „მექანისტური“ (ანუ, სხვაგვარად, „არაზებუნებრივი“) პროცესებით აიხსნება. მრავალ ცნობილ ბიოლოგს გამოუთქვამს მოსაზრება, რომ თანამედროვე (მეოცე საუკუნის მეორე ნახევრიდან დაწყებული) მოლეკულური ბიოლოგიის მიღწევებმა გააქარწყლა წარმოდგენა, რომ რაიმე მიზანმიმართული ან ღვთიური წამმართავი ძალა ჩარეულია ევოლუციის მიმდინარეობაში. სხვაგვარად რომ ვთქვათ, ამგვარი ძალების არსებობის აღიარება არცერთი ევოლუციური პროცესის ასახსნელად არაა საჭირო. ამასთან ერთად, რაც არანაკლებ მნიშვნელოვანია, ბიოლოგიური პროცესები, რომლების საფუძველზე ადამიანის ევოლუციაც მიმდინარეობდა, არ ქმნიან ადამიანების საზოგადოებებში არსებული მორალური კანონების საფუძველს: ადამიანთა მოქმედების ეს მხარე ბუნებრივი ევოლუციის მიღმა რჩება (არაა „ჩადებული“ ადამიანის, როგორც სახეობის, ნიშან-თვისებებსა თუ „ბუნებაში“) და მხოლოდ ადამიანთა მოქმედებითაა განსაზღვრული.
როგორც ვნახეთ, დარვინის იდეების გაცნობიერებასა და აღიარებას როგორც ბიოლოგების, ასევე ფართო საზოგადოების მიერ თითქმის 80 წელი დასჭირდა. ევოლუციურ ბიოლოგიაში მიმდინარე კვლევების ყველა შედეგის გააზრება, ისევე როგორც მორალური კანონების ჭეშმარიტი საფუძვლების გამორკვევა, ალბათ კიდევ უფრო დიდხანს გასტანს. ეჭვსგარეშე მხოლოდ ის გარემოებაა, რომ დარვინის ნააზრევის გავლენა ევოლუციის შემსწავლელი მეცნიერების (და არა მარტო მეცნიერების ამ დარგის) განვითარების ნებისმიერ მიმართულებაზე მომავალშიც მეტად მნიშვნელოვანი იქნება.
გელა თევზაძე.
24 საათი 25.09.11
|
|
18 გენების ჰორიზონტალური გადატანა - უჩვეულო მექანიზმი ევოლუციაში. |
▲ზევით დაბრუნება |
როგორც ცნობილია, გენები (ორგანიზმების ნიშან-თვისებების განმსაზღვრელი ერთეულები) მშობლებისაგან მომავალ თაობას გადაეცემა, და ეს მემკვიდრეობითობის უმთავრესი მექანიზმია. ამ გზით ხორციელდება როგორც გარკვეული ნიშან-თვისებების შენარჩუნება მრავალი თაობის განმავლობაში, ასევე ახალი ნიშან-თვისებების (ცვალებადობის) წარმოქმნა, რადგან ორგანიზმები გენების ნახევარს დედისაგან იღებენ, ნახევარს კი - მამისაგან. გენების ამგვარ გადაცემას სხვაგვარად „ვერტიკალურს“ უწოდებენ, რადგან ერთი თაობა მეორეს, მის მომდევნოს, „ზემოდან ქვემოთ“, „შვეულად“ გადასცემს ნიშან-თვისებების განმსაზღვრელ ფაქტორებს. მაგრამ ასევე არსებობს გენების ე.წ. „ჰორიზონტალური“ გადაცემა, როდესაც გენები (დნ მჟავის ფრაგმენტების სახით) გადაეცემა ერთი „ზრდასრული“ ორგანიზმიდან მეორეს. ასეთ გადაცემას ასევე „ლატერალურს“, ანუ გვერდითს უწოდებენ, რადგან ამ პროცესში მონაწილე ორგანიზმები ერთსა და იმავე თაობას ეკუთვნიან, ერთსა და იმავე „დონეზე“ დგანან და გენეტიკური მასალის გადაცემა არა „ქვემოთ“, არამედ „გვერდითაა“ მიმართული. სხვაგვარად რომ ვთქვათ, გენების ჰორიზონტალური გადატანა (Horizontal Gene Transfer, HGT) ნიშნავს, რომ ერთი ორგანიზმი იღებს მეორე ორგანიზმის გენეტიკურ მასალას, და ამავე დროს მეორე ორგანიზმი პირველის მშობელი არაა. ამ სტატიაში აღწერილია HGT-ს სხვადასხვა მექანიზმი და განხილულია ამ პროცესის მნიშვნელობა ევოლუციაში.
ნახ. 1: გენების ჰორიზონტალური და ვერტიკალური გადატანა. მუქი წითელი წირით აღნიშნულია დნმ - ორგანიზმის გენეტიკური ინფორმაციის მატარებელი მოლეკულა.
ნახ. 2: კონიუგაციის პროცესი. ბაქტერიული უჯრედების შორის წარმოიქმნება დამაკავშირებელი მილი, რომლის საშუალებითაც დნმ ერთი ბაქტერიიდან მეორეში გადადის.
თავიდანვე უნდა აღვნიშნოთ, რომ HGT მეტად მნიშვნელოვანი პროცესია ბაქტერიებისათვის. ამ ორგანიზმებში HGT-ს მექანიზმების კვლევაც გაცილებით უფრო ადრე დაიწყო. ბაქტერიებში, ეუკარიოტებისაგან (ორგანიზმები, რომელთა გენეტიკური მასალა ძირითადად ბირთვშია მოთავსებული) განსხვავებით, გენეტიკური ინფორმაციის ცვალებადობის მრავალი შემთხვევა სწორედ HGT-ს საფუძველზე ხდება, როდესაც გენეტიკური მასალა მათ უჯრედებში სხვა ორგანიზმებიდან გადაიცემა. ამ სტატიის პირველ ნაწილში განვიხილავთ HGT-ს მაგალითებსა და მექანიზმებს ბაქტერიებში, შემდეგ კი - რამდენიმე შემთხვევას, როდესაც HGT ეუკარიოტული ორგანიზმების გენეტიკურ ინფორმაციას ცვლის.
ნახ. 3: HGT ევოლუციაში: ოთხი სახეობის ევოლუცია და HGT-ს ერთი შემთხვევა ამ პროცესში: a - ორი სხვადასხვა სახეობის გენომის განცალკევება საერთო წინაპრისაგან; b - HGT: დნმ-ის გადატანა ერთი სახეობიდან მეორეში; c-е: ახალი დნმ-ის შენარჩუნება, გამრავლება და სახეობის წარმომადგენელთა მნიშვნელოვან ნაწილში გავრცელება; f - სახეობის „გაუმჯობესება“ ახალი დნმ-ის გავლენით (მაგ., ანტიბიოტიკისადმი მდგრადობის განვითარება).
დავიწყოთ იმის განხილვით, თუ როგორ შეიძლება HGT-ს „გამოაშკარავება“, ანუ იმის დადგენა, არის თუ არა რომელიმე ახალწარმოქმნილი ნიშან-თვისება სწორედ „გვერდითი გადატანის“ მექანიზმით, და არა რაიმე სხვა მიზეზით (მაგალითად, ორგანიზმში უკვე არსებული გენების ცვლილებით მუტაციის შედეგად)? რასაკვირველია, საუკეთესო საბუთი იქნებოდა ამ ცვლილების წარმოქმნის უშუალოდ დანახვა, როცა გენეტიკური მასალა (დნმ) ერთი მიკროორგანიზმიდან მეორეს გადაეცემა, და ამ გადაცემის შედეგად პირველი ორგანიზმი რაიმე ახალ თვისებას იძენს. მაგრამ ასეთი დაკვირვება მხოლოდ ლაბორატორიაშია შესაძლებელი, HGT კი ბაქტერიების ევოლუციის პროცესში (ანუ ლაბორატორიის ფარგლებს გარეთ) უკვე მრავალჯერ მომხდარა. ამის გამო, დასკვნები HGT-ს შესახებ ხშირად არაპირდაპირი მტკიცებულებების საფუძველზე კეთდება.
გენების გვერდითი გადატანა ხშირად იწვევს უჩვეულოდ დიდ მსგავსებას ორ ორგანიზმს შორის, მაშინაც კი, როდესაც ეს ორგანიზმები ევოლუციურად საკმაოდ დაშირებულნი არიან. გენეტიკური მასალის გადამცემი (დონორი) ბაქტერიის თვისებები ჩნდება ამ მასალის მიმღებ ორგანიზმს (რეციპიენტს) და, რასაკვირველია, მის შთამომავლობასაც. მეტიც, HGT-ს დროს რეციპიენტი მრავალი ნიშან-თვისებით უფრო მეტად ჰგავს დონორს, ვიდრე რეციპიენტთან ევოლუციურად გაცილებით უფრო ახლო მდგომ სახეობებს. მაგრამ ამ შემთხვევაშიც შესაძლებელია, რომ ახალი თვისება შემთხვევით წარმოიქმნა მუტაციის შედეგად, ამიტომ HGT-ს დამაჯერებელი დასაბუთებისათვის მხოლოდ გარეგნულ ნიშან-თვისებებზე დაკვირვება საკმარისი არაა: საჭიროა დნმ-ის მოლეკულური ანალიზიც (მისი შემადგენელი ფუძეების თანმიმდევრობის - ე.წ. სიკვენსის) განსაზღვრა.
სიკვენსის საფუძველზე შესაძლებელი გახდა HGT-ს მრავალი შემთხვევის გამოაშკარავება, ისევე როგორც გენების „ევოლუციური ისტორიის“ დადგენა. HGT-ს შედეგად წარმოქმნილი გენების გარკვეული ფრაგმენტი მნიშვნელოვნად განსხვავდება იმ ფორმებისაგან, რომლებიც წინაპრებისაგან იყო მიღებული „ვერტიკალური გადაცემის“ შედეგად.
როგორ ხდება გენების გვერდითი გადაცემა, და რომელი გენების გადაცემა ხდება ყველაზე ხშირად? ახალი ნიშან-თვისებების წარმოქმნა HGT-ს გზით სამ მოთხოვნილებას უნდა აკმაყოფილებდეს: 1) უნდა არსებობდეს მექანიზმი, რომლის საშუალებითაც დონორის დნმ გადაიყვანება რეციპიენტის ორგანიზმში; 2) მიღებული გენები უნდა „ჩაინერგოს“ რეციპიენტის გენომში (გენების ერთობლიობაში); 3) ინფორმაციის ამოკითვა ახალი გენებიდან უნდა ხდებოდეს ისევე, როგორც რეციპიენტის სხვა („ძველი“) გენებიდან. პირველი ორი ეტაპი, თავის მხრივ, შესაძლოა სამი პროცესის საფუძველზე განხორციელდეს. ეს პროცესებია ტრანსფორმაცია, ტრანსდუქცია და კონიუგაცია.
ტრანსფორმაციის დროს ბაქტერიული უჯრედი გარემოდან შთანთქავს დნმ-ის მოლეკულებს. რადგან ამ შემთხვევაში დნმ-ის „წარმოშობას“ მნიშვნელობა არა აქვს, ტრანსფორმაციის გზით შესაძლებელია დნმ-ის გადაცემა ევოლუციურად მეტად დაშორებული ორგანიზმების შორის. ზოგიერთი სახეობის ბაქტერიებს დნმ-ის შთანთქმის (ე.წ. კომპეტენტურობის) უნარი მთელი სიცოცხლის განმავლობაში აქვთ, ზოგიერთნი კი მხოლოდ სიცოცხლის გარკვეულ პერიოდში იძენენ ამ თვისებას. თუმცა მაღალი კომპეტენცია სულაც არ ნიშნავს, რომ ასეთ ბაქტერიებში HGT დიდი სიხშირით ხდებოდა: არსებობს მაგალითები, როცა კომპეტენტური ბაქტერიების დნმ შეიცავს მხოლოდ ორ-სამ ფრაგმენტს, რომელსაც აშკარად „უცხო წარმოშობა“ აქვს.
ახალი გენეტიკური მასალის გადატანა უჯრედში შეიძლება ასევე მოხდეს ბაქტერიაში ვირუსის (ბაქტერიოფაგის) შეჭრის შედეგად. ამ პროცესს ტრანსდუქცია ეწოდება. ბაქტერიოფაგი მრავლდება მასპინძლის უჯრედში, ან „ჩაშენდება“ ბაქტერიის დნმ-ში. ამ გზით ბაქტერიის გენეტიკურ მასალას მნიშვნელოვანი დანამატი უჩნდება. ამასთანავე, ბაქტერიულ დნმ-ში „ჩაშენებით“ ვირუსის გენეტიკური მასალა თავს იცავს მასპინძლის იმ ფერმენტებისაგან, რომლებიც უცხო დნმ-ს ანაწევრებენ და ამგვარად უშლიან ხელს მასპინძლის უჯრედზე ვირუსების შეტევას. ისევე, როგორც ტრანსფორმაციის შემთხვევაში, აუცილებელი არაა, დონორისა და რეციპიენტის უჯრედები დნმ-ის გადატანისას ერთსა და იმავე ადგილას იმყოფებოდნენ: ადვილი წარმოსადგენია, რომ ერთ ბაქტერიულ უჯრედში ჩაშენებულ, ხოლო შემდგომ ამოჭრილ და გამრავლებულ ვირუსულ დნმ-ს შეიძლება „ნარჩენი“ ბაქტერიული დნმ თან „წაჰყვეს“. თუკი ეს ვირუსი სხვა სახეობის ბაქტერიაში შეიჭრება და აქაც მოხდება დნმ-ის ჩაშენება, ამ პროცესის შედეგად მოხდება ერთი სახეობის ბაქტერიის გენეტიკური მასალის მეორესთან შერწყმა, სადაც ვირუსს გადამტანის ფუნქცია აქვს.
გ) კონიუგაცია, წინა ორი მექანიზმისაგან განსხვავებით, საჭიროებს ორი ორგანიზმის უშუალო სიახლოვეს. ამ დროს ერთი ბაქტერიის დნმ მეორე ბაქტერიაში გადადის ამ ორ ორგანიზმს შორის წარმოქმნილი დამაკავშირებელი მილების საშუალებით. აქაც შესაძლებელია დონორი ბაქტერიის გენეტიკური მასალის გადატანა რეციპიენტში, და რეციპიენტის დნმ-ში ჩაშენება. კონიუგაცია ჩვეულებრივ ევოლუციურად ახლომონათესავე ბაქტერიული ორგანიზმების შორის ხდება.
მიუხედავად იმისა, რომ გენეტიკური ინფორმაციის მიმოცვლა ბაქტერიებს შორის მრავალი მექანიზმით შეიძლება მოხდეს, მხოლოდ ასეთი გადატანა არ უზრუნველყოფს „ახალშეძენილი“ დნმ-ის სტაბილურობას და, მითუმეტეს, მის მიერთებას ახალი მასპინძლის გენებთან. ამგვარი სტაბილურობის დასამყარებლად საჭიროა, რომ ახალი დნმ-ის დამოუკიდებლად შენარჩუნდეს უჯრედში, რაც შესაძლებელია, თუ 1) დნმ-ის ეს ფრაგმენტი თავისი გამრავლებისათვის საჭირო ინფორმაციის მატარებელ გენებს შეიცავს; 2) ახალი დნმ გაერთიანდება მასპინძლის გენებთან და მათთან ერთად გადაეცემა შემდეგ თაობებს.
რა თვისებები შეიძლება შეიძინოს ორგანიზმმა HGT-ს შედეგად? მათგან ყველაზე მნიშვნელოვანია ანტიბიოტიკებისადმი მდგრადობა. გენები, რომლებიც ანტიბიოტიკებისადმი გამძლეობას განსაზღვრავენ, ხელს უწყობს მათი მატარებელი მიკროორგანიზმის ფართოდ გავრცელებას, ისევე როგორც გამრავლებას იმისდა მიუხედავად, არის თუ არა ტოქსიკური ნაერთები ზრდის გარემოში. ევოლუციის თვალსაზრისით გამართლებულია, რომ ანტიბიოტიკებისადმი მდგრადობის განმსაზღვრელი გენები ბაქტერიის ქრომოსომაში (მისი გენეტიკური მასალის ძირითად ერთეულში) ჩაშენებულნი არ არიან, არამედ არსებობენ დამოუკიდებელი ერთეულების (ე.წ. პლაზმიდების) სახით. პლაზმიდის გადაცემა ერთი ორგანიზმიდან მეორეში გაცილებით უფრო მარტივი და ეფექტურია, ვიდრე ქრომოსომაში ჩაშენებული ფრაგმენტისა, რადგან ეს უკანასკნელი მექანიზმი ჯერ გენების ამოჭრას საჭიროებს, შემდეგ კი მის გადაცემას.
HGT-ით ბაქტერიებმა ასევე შესაძლოა განივითარონ ვირულენტობა, ანუ პათოგენურობა. ანტიბიოტიკისადმი მდგრადობის შეძენის პროცესისაგან განსხვავებით, პათოგენურობის შეძენა იწვევს მიკროორგანიზმის ეკოლოგიური მახასიათებლების მნიშვნელოვან ცვლილებას. პათოგენური ორგანიზმების არსებობა ბაქტერიების ყველა ჯგუფში (ანუ მათი შემთხვევითი განაწილება) იმის მიმანიშნებელია, რომ ვირულენტური თვისებები დამოკიდებულია გენების გარკვეულ ჯგუფზე, რომელიც არაპათოგენურმა ორგანიზმმა შეიძინა HGT-ს გზით უკვე არსებული პათოგენური მიკროორგანიზმისაგან. ცნობილია ლაბორატორიული ცდები, რომლებშიც ნაწლავის ჩხირის (Escherichia coli) უვნებელ ფორმებს შესძინეს ვირულენტობა პათოგენური ბაქტერიებიდან „ვირულენტობის გენების“ გადმოტანის შედეგად. კვლევების საფუძველზე აგრეთვე აღმოჩნდა, რომ ე.წ. „პათოგენურობის კუნძულები“ (pathogenicity islands) ბაქტერიების ქრომოსომებში, რომლებიც ამ ორგანიზმების ვირულენტობის მთავარი განსაზღვრელები არიან, სწორედ HGT-ს გზით იყვნენ შეძენილნი.
არსებობს თუ არა იგივე პროცესები ერთუჯრედიან და მრავალუჯრედიან ეუკარიოტებში? ამ ორგანიზმებში, ბაქტერიებისაგან განსხვავებით, გენები მოთავსებულია არა ერთი, არამედ რამდენიმე ქრომოსომის შემადგენლობაში. ქრომოსომები კი, თავის მხრივ, ბირთვითაა შემოსაზღვრული. ამ ორგანიზმებს მიეკუთვნებიან ერთუჯრედიანი და მრავალუჯრედიანი სოკოები, მცენარეები და ცხოველები. ეუკარიოტების გენომი (გენების ერთობლიობა) ე.წ. ქიმერული სტრუქტურისაა, რადგან იგი შეიცავს როგორც ეუკარიოტული, ასევე ბაქტერიული წარმოშობის გენებს. არსებობს ჰიპოთეზა, რომლის მიხედვითაც ეუკარიოტულ უჯრედში ბირთვის გარეთ არსებული ორგანელები (მაგ., მიტოქონდრიები, რომლებშიც უჯრედისათვის ენერგიის დაგროვება ხდება და რომლებიც საკუთარ დნმ-ს შეიცავენ) სწორედ HGT-ს გზით არიან წარმოქმნილნი. ამ ჰიპოთეზის თანახმად, ორგანელების არსებობა მიანიშნებს, რომ ეუკარიოტულ უჯრედში შეჭრილი იყო მიკროორგანიზმი, რომელმაც სიმბიოტური თანაარსებობის გზით სპეციალიზირებული ფუნქცია (მიტოქონდრიების შემთხვევაში - ენერგიის დაგროვება) შეიძინა, და საკუთარი დნმ-იც შეინარჩუნა. ასევე შესაძლებელია, რომ ამგვარი თანაარსებობის დროს ხდებოდა HGT - ორგანელიდან („ყოფილი ბაქტერიიდან“) ბირთვში დნმ-ის ფრაგმენტების და, მაშასადამე, გენების გადატანა. გამოთვლილია, რომ ზოგიერთი მცენარის გენების ~9%25 ბაქტერიული წარმოშობისაა. უნდა აღინიშნოს, რომ გენების ამგვარი გადატანის ევოლუციური მნიშვნელობა ბოლომდე არაა შეფასებული იმის გამო, რომ ჯერ-ჯერობით ბაქტერიებისა და ეუკარიოტების სიმბიოზური თანაარსებობის მრავალი შემთხვევა სათანადოდ შესწავლილი არაა.
HGT და ადამიანის გენომი: ადამიანის გენომის ანალიზის შედეგად აღმოჩნდა, რომ 40-მდე გენი (ადამიანის ~25 ათასი გენიდან) ბაქტერიული წარმოშობისაა და, შესაძლოა, ადამიანის რომელიმე წინაპრის (ხერხემლიანი ცხოველის) გენომში შეტანილი იქნა HGT-ს მექანიზმით. ასეთი ვარაუდი გულდასმით უნდა შემოწმდეს, რადგან საბოლოო დასკვნისათვის აუცილებელია როგორც დეტალური გენეალოგიური ანალიზის ჩატარება, აგრეთვე მრავალი სხვადასხვა წარმოშობის ადამიანის დნმ-ის ანალიზი.
დასკვნა: ჰიპოთეზები, რომლის თანახმადაც ეუკარიოტების წარმოშობა რამდენიმე პროკარიოტული ორგანიზმისა და ეუკარიოტული უჯრედის სიმბიოტური თანაარსებობის შედეგად მოხდა, დამაკმაყოფილებლად ვერ დასტურდება HGT-ს კვლევის შედეგად მიღებული დასკვნებით. მაგალითად, ორი ან სამი პროკარიოტული „წინაპრის“ გამოყოფა მხოლოდ ზოგიერთი ცალკეული გენისთვისაა მართებული, და მხოლოდ იმ შემთხვევებში, როდესაც ხდებოდა HGT მიტოქონდრიიდან ბირთვში. ისიც გაურკვეველია, რა იწვევს ამ შეუსაბამობას: შესწავლილი გენების რაოდენობის სიმცირე, ევოლუციის თვალსაზრისით გვიანდელი HGT-ს მოქმედება, რომლიც გამოც ადრინდელი პროცესების იდენტიფიკაცია რთულდება, თუ თვით ამ ჰიპოთეზის არამართებულობა.
ერთ-ერთი ჰიპოთეზა, რომელიც საბოლოოდ უარყოფილია HGT-ს კვლევის შედეგად, „გენეტიკური შერწყმის“ ჰიპოთეზაა. მის თანახმად, ბაქტერიული დნმ-ის გადატანა ეუკარიოტების ადრეულ წინაპრებში მოხდა, და მათგან ყველა გვიანდელ ფორმას გადაეცა. დაგროვებული შედეგები კი, პირიქით, მიანიშნებენ, რომ ბაქტერიების გენეტიკური მასალის გადატანა HGT-ს მექანიზმით ერთმანეთისაგან დამოუკიდებლად ხდებოდა როგორც წინაპარ ორგანიზმებში (მაგ., ერთუჯრედიან ცხოველებში), ასევე შედარებით გვიან წარმოქმნილ ფორმებში (მაგ., ხერხემლიანებში). ნებისმიერ შემთხვევაში, ნათელია, რომ ევოლუციის პროცესებზე მსჯელობისას უნდა უარვყოთ მოსაზრება, რომ გენების ვერტიკალური გადატანა (მშობლების ორგანიზმიდან შვილებისაკენ) ევოლუციის ერთადერთი მექანიზმი და ძირითადი კანონია, რომლისგანაც გამონაკლისები არ არსებობს.
გელა თევზაძე.
24 საათი 09.10.11
|
|
19 2011 წლის ნობელის პრემია ქიმიაში: „ოქროს კვეთის“ კრისტალები. |
▲ზევით დაბრუნება |
ნობელის პრემია ქიმიაში 2011 წელს მიენიჭა დენ შეხტმანს (Dan Shechtman, Technion - Israel Institute of Technology, Haifa, Israel), უჩვეულო სტრუქტურის კრისტალების (კვაზიკრისტალების) აღმოჩენისათვის. ქვემოთ გთავაზობთ ამ თემაზე გამოსული სამეცნიერო-პოპულარული მიმოხილვის თარგმანს:

ნახ.1: დენ შეხტმანი.
დენ შეხტმანის ლაბორატორიულ ჟურნალში იმ მონაცემებთან, რომლებმაც მას 2011 წლის ნობელის პრემია მოუტანა, სამი კითხვის ნიშანია დასმული. კრისტალში, რომელსაც შეხტმანი აკვირდებოდა, ატომები უჩვეულო - მეტიც, ბუნების კანონებით აკრძალულ - სტრუქტურას ქმნიდნენ. გარკვეულწილად, ეს ისეთივე შეუძლებელი ჩანდა, როგორც ბრტყელი ექვსკუთხედებისაგან შემდგარი სფერო. მას შემდეგ, მოზაიკური კონსტრუქციების, ოქროს კვეთის პრინციპისა და ხელოვნების სხვადასხვა დარგების დახმარებით შესაძლებელი გახდა ამ გასაოცარი მოვლენის ახსნა.
1982 წლის 8 აპრილს შეხტმანმა ელექტრონული მიკროსკოპის დახმარებით მიიღო ალუმინისა და მანგანუმის შენადნობის ატომური სტრუქტურის სურათები. უჩვეულო სტრუქტურა ყველა არსებულ წარმოდგენას ეწინააღმდეგებოდა: იგი შედგებოდა კონცენტრული წრეებისაგან, რომელთაგან ყოველი ერთმანეთისაგან თანაბრად დაშორებულ ათ კაშკაშა წერტილს შეიცავდა.
ამ შენადნობის მიღებისას, გალღობილი ლითონების სწრაფი გაცივება ხდებოდა, და ტემპერატურის ასეთ მკვეთრ ცვლილებას ატომების განლაგებაში სრული ქაოსი უნდა გამოეწვია. მაგრამ ატომები სავსებით მოწესრიგებულად იყვნენ განლაგებული, თუმცა ეს განლაგება ბუნების კანონებს ეწინააღმდეგებოდა. ოთხი ან ექვსი წერტილის არსებობა შესაძლებელი იქნებოდა, ათის კი სრულიად წარმოუდგენელი იყო. ამიტომაც ჩაწერა შეხტმანმა თავის ჟურნალში: „10-მაგი???“ („10-fold???“).
რატომ იყო 10 წერტილი ასე უჩვეულო და მნიშვნელოვნად განსხვავებული 4 - ან 6 წერტილიან („ჩვეულებრივ“) წრეებისაგან, ამის ახსნას ქვემოთ შევეცდებით.
„მწვერვალებისა“ და „ღრმულების“ ურთიერთქმედება. შეხტმანის ექსპერიმენტისა და მისი შედეგის გასაგებად, წარმოვიდგინოთ შემდეგი ცდა, რომლის ჩატარება საშუალო სკოლის ფიზიკის გაკვეთილზე შეიძლება: სინათლე მიმართულია ლითონის ფილაზე, რომელშიც ნახვრეტებია გაკეთებული. ადვილი წარმოსადგენია, რომ სხივები ნახვრეტებს შორის გაივლიან, შემდეგ კი გარკვეული კანონზომიერებით აირეკლებიან. სინათლის ტალღების ფილაში გავლა ან მისგან არეკვლა შეიძლება ისევე წარმოვიდგინოთ, როგორც ზღვის ტალღების მიერ რაიმე დაბრკოლების გადალახვა.
ფილის მეორე მხარეს, სინათლის ტალღები ნახევარწრის სახით ვრცელდებიან და ერთმანეთთან ურთიერთქმედებენ. ამის შედეგად, ფილის უკან (სინათლის წყაროს საწინააღმდეგო მხარეს) განლაგებულ ეკრანზე ნათელი და ბნელი ხაზების მონაცვლეობას დავინახავთ: ნათელი ზოლები წარმოიქმნება სინათლის ტალღების ინტენსივობის მწვერვალები (crests) გადაიკვეთება და ერთმანეთს გააძლიერებს, ხოლო ბნელი ზოლები მწვერვალებისა (crests) და ღრმულების (trough) ურთიერთქმედების შედეგია, რადგან ისინი ერთმანეთს ანეიტრალებენ. ნათელი და ბნელი ზოლების ამგვარი მონაცვლეობა დიფრაქციული გამოსახულებას, ანუ არეკვლის გამოსახულებას წარმოქმნის.
დენ შეხტმანმა 1982 წელს მსგავსი სურათი მიიღო. მისი ექსპერიმენტი აქ აღწერილისაგან სამი გარემოებით განსხვავდებოდა: 1) სინათლის ნაცვლად ელექტრონების ნაკადი იყო გამოყენებული; 2) „ფილა“ ამ შემთხვევაში შედგებოდა ლითონის ატომებისაგან, „ნახვრეტები“ კი მათ შორის არსებული მანძილი გახლდათ; 3) ექსპერიმენტი არა ორ, არამედ სამ განზომილებაში ჩატარდა.
დიფრაქციული სურათის თანახმად, ატომები მოწესრიგებული კრისტალის სახით იყო განლაგებული. ამაში, თავისთავად, არაფერია უჩვეულო: ყველა მყარი სხეული - ყინული თუ ოქრო - მოწესრიგებული კრისტალებისაგან შედგება. მაგრამ დიფრაქციული სურათი, სადაც ათი კაშკაშა წერტილი წრეს ქმნიდა, ჭეშმარიტად დაუჯერებელი იყო: შეხტმანს, კრისტალების ელექტრული მიკროსკოპით ანალიზის დიდი გამოცდილების მიუხედავად, ასეთი განლაგება არასდროს ენახა. მეტიც, ასეთი კრისტალი ნახსენებიც კი არ იყო კრისტალოგრაფიის საერთაშორისო ტაბულებში (International Tables for Crystallography) - გამოცემაში, რომელიც ამ დარგის ძირითადი ცნობარია. იმ დროისათვის, მეცნიერები მიიჩნევდნენ, რომ ათი წერტილისაგან შემდგარი წრიული ფორმა უბრალოდ შეუძლებელია, და ამ შეუძლებლობის დამტკიცება მარტივად და თვალსაჩინოდ ითვლებოდა.
განლაგება, რომელიც ლოგიკას ეწინააღმდეგება. კრისტალის შემადგენლობაში არსებული ატომების განლაგება განმეორებადია, და ამ განლაგებას კრისტალის ქიმიური შემადგენლობაზე დამოკიდებული სიმეტრია ახასიათებს. სამმაგი სიმეტრიის შემთხვევაში, ყოვლი ატომი სამი იდენტური ატომითაა გარშემორტყმული. ამ გამოსახულების შემოტრიალება 120 გრადუსით (360 გრადუსის, ანუ სრული წრის 1/3-ით) იგივე გამოსახულებას მოგვცემს. მსგავსი განლაგებაა ოთხმაგი და ექვსმაგი სიმეტრიის შემთხვევაშიც: გამოსახულება მეორდება, შესაბამისად, 90 (360/4) და 60 (360/6) გრადუსით შემოტრიალებისას.
მაგრამ ხუთმაგი სიმეტრია ამ კანონზომიერებას აღარ ემორჩილება, რადგან გამოსახულების 72 გრადუსზე (360/5) შემოტრიალებისას იგი აღარ მეორდება: ხუთმაგი სიმეტრიის მქონე კრისტალში ატომთა ზოგიერთი წყვილი ერთმანეთთან უფრო ახლოსაა განლაგებული, ვიდრე სხვები. ამის გამო, ხუთმაგი (ისევე როგორც შვიდმაგი და ა.შ.) სიმეტრიის კრისტალის მიღება შეუძლებლად მიაჩნდათ.
შეხტმანს კი შეეძლო, თავისი კრისტალი 36 გრადუსით (360/10) შემოეტრიალებინა და იგივე გამოსახულება მიეღო. ამრიგად, მან ათმაგი სიმეტრია მიიღო, რაც შეუძლებელი ჩანდა. ამიტომაც გასაკვირი არცაა კითხვის ნიშანთა სიმრავლე მისი ლაბორატორიული ჩანაწერის შემდეგ...
სახელძღვანელოების განმარტებით - არასწორი შედეგი: პირველი, რაც შეხტმანმა გადაამოწმა, კრისტალის სტრუქტურა იყო: არსებობდა ალბათობა, რომ მან ე.წ. „ტყუპი კრისტალი“ (twin crystals) მიიღო: უჩვეულო დიფრაქციულ სურათი იმ შემთხვევაშიც წარმოიქმნებოდა, თუ ორი კრისტალი ერთმანეთს „შეეზრდებოდა“. ამგვარი სტრუქტურის არსებობის შესაძლებლობა მაშინვე გამოირიცხა: შეზრდილი კრისტალებისათვის დამახასიათებელი ნიშნებიდან არცერთი არ გამოაშკარავდა. შემდეგ შეხტმანმა კრისტალი შემოატრიალა და კვლავ დაუკვირდა ელექტრონული მიკროსკოპის დახმარებით. ამ ექსპერიმენტის შედეგად ერთი მნიშვნელოვანი ფაქტი აღმოჩნდა: თვით კრისტალს არ ჰქონდა ათმაგი სიმეტრია, მაგრამ იგი ხუთმაგი (მაშინდელი წარმოდგენით, ასევე შეუძლებელი) სიმეტრიით იყო აგებული. აქედან მხოლოდ ერთი დასკვნა იყო შესაძლებელი - რომ მაშინდელი წარმოდგენა კრისტალების სტრუქტურის შესახებ სინამდვილეს არ შეესაბამებოდა.
როდესაც შეხტმანმა თავისი ექსპერიმენტების შედეგები კოლეგებს გააცნო, პასუხად უნდობლობისა და დაცინვის ნარევი მიიღო. ბევრნი დარწმუნებულნი იყვნენ, რომ მისი მონაცემები „ტყუპი კრისტალის“ დიფრაქციულ სურათს ასახავდა. ლაბორატორიის უფროსმა სარკასტულად ურჩია კიდეც, კრისტალოგრაფიის საფუძვლების სახელმძღვანელო წაეკითხა, ბოლოს კი მოითხოვა, რომ შეხტმანს მისი ლაბორატორია დაეტოვებინა. შეხტმანმა, რასაკვირველია, იცოდა, რა ეწერა სახელმძღვანელოში, მაგრამ საკუთარი შედეგების მაინც უფრო მეტად სჯეროდა. საბოლოოდ, მდგომარეობა მეტისმეტად უხერხული გახდა.
ბრძოლის შედეგად მოპოვებული აღიარება. 1983 შეხტმანის შედეგებით დაინტერესდა ილან ბლეხი (Ilan Blech). ამ ორი მეცნიერის თანამშრომლობის შედეგად - და შეხტმანის მიერ მიღებული დიფრაქციული გამოსახულების საფუძველზე - გაირკვა კრისტალის ატომური სტრუქტურა. ამ შედეგების ამსახველი სტატია, რომელიც „Journal of Applied Physics“-ში გააგზავნეს, რედაქტორის მიერ მიღებისთანავე იქნა უარყოფილი, რეცენზირებაზე გაგზავნის გარეშე. მხოლოდ მაშინ, როდესაც კიდევ ორმა ცნობილმა მეცნიერმა გამოაცხადა, რომ ეს შედეგები სანდო იყო, შესაძლებელი გახდა სტატიის გამოქვეყნება - „Physical Reviews Letters“-ში, 1984 წელს. ამ ნაშრომმა უდიდესი შთაბეჭდილება მოახდინა, რადგან მასში აღწერილმა შედეგებმა ეჭვქვეშ დააყენა დებულება, რომელიც ურყევ ჭეშმარიტებად იყო მიჩნეული: რომ ყველა კრისტალის აგებულება განმეორებადი, პერიოდული ერთეულებით განისაზღვრება.
სტატიის გამოქვეყნების შემდეგ, შეხტმანი კიდევ უფრო მეტი კრიტიკის ქარცეცხლში გაატარეს. მაგრამ ამასთანავე, ბევრმა ფიზიკოსმა და კრისტალოგრაფმა გაიხსენა საკუთარი - და ექსპერიმენტულ შეცდომად მიჩნეული - შედეგები, რომელნიც რვამაგი ან თორმეტმაგი სიმეტრიის არსებობაზე მიანიშნებდნენ. ამრიგად, უჩვეულო სიმეტრიის მქონე კრისტალების არსებობა უკვე დადგენილი იყო. მთავარი საკითხი (ამ კრისტალებში ატომების განლაგების მექანიზმების დადგენა) მაინც გადაუჭრელი რჩებოდა. პასუხი ყველაზე ნაკლებად მოსალოდნელი არედან წარმოიქმნა - მათემატიკური თამაშებიდან, რომლებიც მოზაიკის ელემენტებს იყენებდნენ.
მოზაიკა და „ოქროს კვეთი“. 1960-იან წლებში, ერთ-ერთი პოპულარული მათემატიკური ამოცანა იყო მოზაიკის შედგენა შეძლებისდაგვარად მცირე რაოდენობის ნაწილებისაგან იმგვარად, რომ ეს ანაწყობი არადროს განმეორებულიყო. სხვაგვარად რომ ვთქვათ, საჭირო იყო უმცირესი შესაძლებელი ზომის აპერიოდული მოზაიკის შედგენა. 1966 წელს პირველად მიიღეს ასეთი კონსტრუქცია, მაგრამ ის 20 ათასი დეტალისაგან შედგებოდა და, რასაკვირველია, ვერ აკმაყოფილებდა მათემატიკოსებისათვის დამახასიათებელ სწრაფვას სიმარტივისა და სიცხადისაკენ.
1970-იანი წლების შუაგულში, ბრიტანელი მათემატიკოსი, როჯერ პენროუზი (Roger Penrose) თავის გამოთვლებში სწორედ ამ სისტემის მაქსიმალურ გამარტივებას ცდილობდა და მიზანს მიაღწია კიდეც: პენროუზის შექმნილი ანაწყობი მხოლოდ ორი სხვადასხვა დეტალისაგან შედგებოდა, მაგრამ სრულიად აპერიოდული (არაგანმეორებადი) იყო. აღმოჩნდა, რომ შუა საუკუნეების ისლამურ არქიტექტურაში მსგავსი პრინციპი გამოიყენებოდა (მაგ., ალამბრას სასახლე ესპანეთში, დარბ-ი იმამის ტაძარი ირანში). ამ მოზაიკის აგებისას მხოლოდ ხუთი განსხვავებული დეტალია გამოყენებული.
კრისტალოგრაფი ალან მაკეი (Alan McKay) შეეცადა, კრისტალების სტრუქტურის კვლევისას დაედგინა, შეეძლოთ თუ არა ატომებს, მოზაიკის შემადგენელი დეტალების მსგავსად, აპერიოდული სტრუქტურის წარმოქმნა. მაკეის მიერ შემუშავებული და პენროუზის სისტემის ანალოგიური მოდელი, სადაც დეტალების გადაკვეთის წერტილებს ატომები წარმოადგენდნენ, დიფრაქციულ სურათში სწორედ ათმაგ სიმეტრიას იძლეოდა: წრეზე განლაგებულ ათ კაშკაშა წერტილს!
ფიზიკოსები პოლ სტაინჰარდტი და დოვ ლევაინი (Paul Steinhardt, Dov Levine) თავის სტატიაში, რომელიც 1984 წელს Physical Review Letters-ში გამოქვეყნდა, მაკეის მოდელისა და შეხტმანის დიფრაქციის ურთიერთკავშირს განიხილავდნენ. მათვე ეკუთვნის ამ სტრუქტურის სახელწოდებაც - „კვაზიკრისტალები“.
ასევე გამოირკვა, რომ როგორც პენროუზის მოზაიკაში, ასევე კვაზიკრისტალებში ხშირად მეორდება „ოქროს კვეთის“ შეფარდება: მთელი ნაწილი დიდ და მცირე ფრაგმენტებადაა გაყოფილი იმგვარად, რომ დიდი ფრაგმენტი შეეფარდება მთელ ნაწილს ისევე, როგორც მცირე ფრაგმენტი დიდს. ოქროს კვეთში ეს შეფარდება უდრის 1.61803398874989-ს. იგივე შეფარდება მიიღება ფიბონაჩის რიგშიც: 1, 1, 2, 3, 5, 8, 13, 21, 34, 55, 89, 144, 233... ამ რიგში ყოველი რიცხვი წინა ორი რიცხვის ჯამს წარმოადგენს. ფიბონაჩის რიგში, გარკვეული რიცხვიდან დაწყებული (21/13, 34/55, ... 233/144, 377/233...) ყოველი რიცხვის შეფარდება წინა რიცხვთან ოქროს კვეთთანაა მიახლოებული. კვაზიკრისტალებში ატომებს შორის მანძილთა შეფარდება სწორედ ამ რიცხვით განისაზღვრება, თვით მანძილთა სიდიდეები კი ფიბონაჩის რიგს მიესადაგება.
შეხტმანის აღმოჩენამ მნიშვნელოვნად შეცვალა თვით ტერმინ „კრისტალის“ განსაზღვრებაც. ტრადიციულად კრისტალი განისაზღვრებოდა, როგორც „ნივთიერება, რომელშიც ატომები მოწესრიგებულ და განმეორებად სტრუქტურას ქმნიან“. კვაზიკრისტალების აღმოჩენამ ამ განსაზღვრების მართებულობა ეჭვქვეშ დააყენა. 1992 წელს მიღებული განსაზღვრების თანახმად, „კრისტალი ყველა ის მყარი ნივთიერებაა, რომელსაც გამოკვეთილი დიფრაქციული გამოსახულება აქვს“. როგორც ვხედავთ, ამ განსაზღვრებაში „მოწესრიგებულობა და განმეორებადობა“ აღარაა ჩართული, ამიტომ იგი არა მარტო ჩვეულებრივ კრისტალებს, არამედ კვაზიკრისტალებსაც (და ჯერ კიდევ აღმოუჩენელ სახესხვაობებს) მოიცავს.
კვაზიკრისტალები ბუნებაში და წარმოებაში: შეხტმანის მიერ კვაზიკრისტალების აღმოჩენის შემდეგ, ლაბორატორიებში ასეულობით სხვადასხვა კვაზიოკრისტალის სინთეზი შესძლეს. მაგრამ ბუნებრივი კვაზიკრისტალი მხოლოდ 2009 წელს აღმოაჩინეს აღმოსავლეთ რუსეთში. ეს კვაზიკრისტალი, რომელსაც ათმაგი სიმეტრია ახასიათებს, ალუმინის, სპილენძისა და რკინის ატომებისაგან შედგება. მას იკოსაედრიტი ეწოდა, გეომეტრიული ფიგურის - იკოსაედრის - საფუძველზე. იკოსაედრს 20 წახნაგი აქვს, სადაც თითოეული წახნაგი ტოლგვერდა (სრულყოფილ) სამკუთხედს წარმოადგენს. იკოსაედრის პროპორციებიც ოქროს კვეთით აღიწერება.
კვაზიკრისტალები ასევე გამოიყენება უჩვეულოდ მყარი ფოლადის წარმოებაშიც. ასეთ ფოლადს ორი ურთიერთშერწყმული სტრუქტურა აქვს: მაგარი ფოლადის კვაზიკრისტალები ჩაშენებულია რბილ ფოლადში. ამ შენადნობიდან ქირურგიულ ინსტრუმენტებსაც (მაგ., თვალის ოპერაციისათვის საჭირო ნემსებს) ამზადებენ.
კვაზიკრისტალები, მათი სიმაგრის მიუხედავად, ძალიან ადვილად იმსხვრევა. ისინი ცუდად ატარებენ დენსა და სითბოს, მათი ზედაპირი არ იკრავს სხვადასხვა სითხეების ნარჩენებს. ამ თვისებების საფუძველზე კვაზიკრისტალები გამოიყენება თერმოელექტრულ მასალებში, რომლებიც სითბოს ელექტრობად გარდაქმნიან. ასევე შესაძლებელია კვაზიკრისტალების გამოყენება ტაფების ზედაპირის მასალის სახით, სითბოს იზოლაციისათვის მანქანის ძრავებში, და ა.შ.

ნახ. 2: ათმაგი სიმეტრიის დიფრაქცია: სურათი მეორდება გამოსახულების 36 გრადუსით შემოტრიალებისას.
ნახ. 3: სხვადასხვა სახის სიმეტრია კრისტალებში: ხუთმაგი სიმეტრიის მქონე კრისტალის სტრუქტურის განმეორება შეუძლებელია.

ნახ. 4: პენროუზის მოზაიკა: ასეთი რომბებისაგან შემდგარი სტრუქტურა აპერიოდულია (არაგანმეორებადი).
დასკვნა: დენ შეხტმანის ამბავი უნიკალური ნამდვილად არაა. მეცნიერების ისტორიაში მრავლადაა შემთხვევები, როდესაც მკვლევარებს უხდებოდათ დაკანონებული „ჭეშმარიტების“ წინააღმდეგ გალაშქრება. ეს „ჭეშმარიტება“ კი, როგორც მოგვიანებით ირკვეოდა, მხოლოდ და მხოლოდ შეუმოწმებელი ვარაუდი იყო. შეხტმანის ერთ-ერთი კრიტიკოსი ლაინუს პოლინგი (Linus Pauling) გახლდათ - ნობელის პრემიის ორგზის ლაურეატი. ამგვარად, დიდი მეცნიერებიც კი ზოგჯერ ვერ სცდებიან „საყოველთაო აზრით“ შემოსაზღვრულ ფარგლებს. საკუთარი შედეგების ნდობა და მიღებულ ჭეშმარიტებაში დაეჭვების უნარი მკვლევარისათვის მეტად მნიშვნელოვანი თვისებაა.
გელა თევზაძე.
24 საათი 23.10.11
|
|
20 2011 წლის ნობელის პრემია ფიზიოლოგიასა ან მედიცინაში: თანდაყოლილი და შეძენილი იმუნიტეტი. |
▲ზევით დაბრუნება |
ნობელის პრემია ფიზიოლოგიასა ან მედიცინაში 2011 წელს მიენიჭა სამ მეცნიერს. ლაურეატები გახდნენ ბრიუს ბიუტლერი (Bruce A. Beutler), ჟიულ ჰოფმანი (Jules A. Hoffman), და რალფ სტაინმანი (Ralph M. Steinman). ბიუტლერმა და ჰოფმანმა, მათი წვლილისათვის თანდაყოლილი იმუნიტეტის კვლევაში, პრემიის თანხის ნახევარი გაინაწილეს (სხვაგვარად რომ ვთქვათ, თითოეულმა პრემიის მეოთხედი მიიღო); სტაინმანს, რომელმაც აღმოაჩინა და შეისწავლა ადაპტური (ანუ შეძენილი) იმუნიტეტის განმსაზღვრელი მნიშვნელოვანი მექანიზმი, პრემიის მეორე ნახევარი მთლიანად ერგო. ასევე მნიშვნელოვანია, რომ სტაინმანმა პრემია ფაქტობრივად გარდაცვალების შემდეგ მიიღო: თუმცა ნობელის პრემიის მინიჭება სიკვდილის შემდეგ დაუშვებელია, ამ შემთხვევაში გამონაკლისი შესაძლებელი გახდა - ნობელის პრემიაზე წარდგენისას სტაინმანი ჯერ კიდევ ცოცხალი იყო, და პრემიის მინიჭებამდე მხოლოდ რამდენიმე დღით ადრე გარდაიცვალა.

ნახ. 1: ბრიუს ბიუტლერი (Bruce A. Beutler, University of Texas Southwestern Medical Center, Dallas, USA), ჟიულ ჰოფმანი (Jules A. Hoffman, Institut de Biologie Moléculaire et Cellulaire, Strasbourg, France), რალფ სტაინმანი (Ralph M. Steinman, Rockefeller University, New York, USA).
ნობელის კომიტეტის განცხადებაში ნათქვამია:
ნობელის პრემიის წლევანდელი ლაურეატების კვლევების შედეგად, ჩვენს წარმოდგენაში იმუნური სისტემის შესახებ მნიშვნელოვანი გარდატეხა მოხდა: მათ აღმოაჩინეს იმუნური სისტემის აქტივირების ძირეული კანონზომიერებები.
მეცნიერები დიდი ხნის განმავლობაში ეძებდნენ იმ მთავარ ფაქტორებს, რომლების მოქმედება განსაზღვრავს იმუნურ პასუხს - ადამიანისა და ცხოველების მთავარ თავდაცვით მექანიზმს ბაქტერიებისა და სხვა მიკროორგანიზმებისაგან. ბრიუს ბიუტლერმა და ჯულს ჰოფმანმა აღმოაჩინეს რეცეპტორები - ცილები, რომლებიც ამოიცნობენ ასეთ მიკროორგანიზმებს და ამოცნობის შედეგად იწვევენ თანდაყოლილი იმუნიტეტის გააქტიურებას. რალფ სტაინმანმა აღმოაჩინა დენდრიტული უჯრედები, ისევე როგორც მათი უნიკალური უნარი, გაააქტიურონ და არეგულირონ შეძენილი იმუნიტეტი, რომლის მოქმედების შედეგადაც მიკროორგანიზმები ადამიანის სხეულიდან განიდევნებიან.
ნობელის პრემიის ლაურეატების კვლევის შედეგად გამოაშკარავდა, როგორ მოქმედებს (და ენაცვლება ერთმანეთს) იმუნური პასუხის თანდაყოლილი და შეძენილი ფაზები, და როგორ ხდება მათი გააქტივება. ამის საფუძველზე, ავადმყოფობათა წარმოქმნის მექანიზმების დეტალური შესწავლაც გახდა შესაძლებელი. ამ შრომებმა ახალი პერსპექტივები შექმნა სხვადასხვაგვარი ინფექციის, ავთვისებიანი სიმსივნისა და მრავალი ანთებითი დაავადების როგორც პროფილაქტიკის, ასევე განკურნების მხრივ.
თავდაცვის ორი ხაზი იმუნურ სისტემაში. ადამიანს სახიფათო სამყაროში უხდება ცხოვრება: მრავალი მიკროორგანიზმი (ვირუსები, ბაქტერიები, სოკოები, ერთუჯრედიანი ცხოველები) გამუდმებით გვიქმნის საფრთხეს, მაგრამ ჩვენ მძლავრი დამცავი მექანიზმებით ვართ აღჭურვილი. თანდაყოლილ იმუნიტეტს, რომელიც დაცვის პირველი ხაზია, შეუძლია გაანადგუროს შემოჭრილი მიკროორგანიზმები და გამოიწვიოს ანთებითი პროცესი, რომელიც უცხო სხეულთა შემოტევას შეაჩერებს. თუ, ამის მიუხედავად, მიკროორგანიზმებმა მაინც მოახერხეს დაცვის ამ ხაზის გარღვევა, შეძენილი იმუნიტეტის მოქმედება იწყება. შეძენილ იმუნურ პასუხში, ანუ დაცვის მეორე ხაზში, მონაწილეობენ T- და B- უჯრედები, რომლებიც მიკროორგანიზმების წინააღმდეგ მოქმედ ანტისხეულებს წარმოქმნიან. იმის შემდეგაც კი, რაც ჩვენი იმუნური სისტემა მომხდურ სხეულებს წარმატებით მოიგერიებს, მას უნარჩუნდება „იმუნოლოგიური მეხსიერება“, რომლის საფუძველზეც მომავალში შესაძლებელი გახდება იგივე მიკროორგანიზმის შეტევის უფრო ეფექტური მოგერიება. როგორც ადვილი მისახვედრია, ამ მექანიზმზეა დამყარებული ავადმყოფობების წინააღმდეგ აცრის პრინციპი: შესუსტებული აგენტის შეყვანა ორგანიზმში იწვევს იმუნოლოგიურ რეაქციას, რომელიც, თავის მხრივ, ანტისხეულების წარმოქმნის წინაპირობაა. ეს ანტისხეულები (უფრო ზუსტად, ამ ანტისხეულების სინთეზისათვის საჭირო ინფორმაცია) ორგანიზმში დიდი ხნის განმავლობაში ინახება და ავადმყოფობის გამომწვევის („ნამდვილი აგენტის“) ორგანიზმში შეჭრის შემთხვევაში სწრაფ იმუნურ პასუხს უზრუნველყოფს.
ამრიგად, იმუნური სისტემის ეს ორი ნაწილი კარგად უზრუნველყოფს ორგანიზმის დაცვას ინფექციებისაგან. მაგრამ, მეორე მხრივ, ამ სისტემის არსებობა გარკვეულ რისკთანაა დაკავშირებული: თუ გააქტიურება ზედმეტად ადვილად ხდება, ან თუ ასეთი გააქტიურების გამოწვევა ენდოგენურ (ორგანიზმის შიგნით არსებულ) მოლეკულებს შეუძლია, ანთებითი დაავადებების განვითარებაა შესაძლებელი.
იმუნური სისტემის კომპონენტები პირველად მეოცე საუკუნის დასაწყისში აღმოაჩინეს. მრავალი აღმოჩენა ამ დარგში ნობელის პრემიით იყო აღნიშნული - მაგალითად, ანტისხეულების აგებულების გამოაშკარავება, ან T-უჯრედების მიერ უცხო სხეულების ამოცნობის მექანიზმი (დაწვრილებით იხ. ქვემოთ). ამ უეჭველად მნიშვნელოვანი კვლევების მიუხედავად, ნობელის პრემიის წლევანდელი ლაურეატების მიერ ჩატარებული კვლევის გარეშე არ გვეცოდინებოდა, რა მექანიზმებით ხდება თანდაყოლილი იმუნიტეტის გააქტიურება, ან როგორ ურთიერთქმედებს ერთმანეთთან თანდაყოლილი და ადაპტური იმუნური სისტემები.
თანდაყოლილი იმუნიტეტის სენსორების („ამომცნობი“ მოლეკულების) აღმოჩენა.
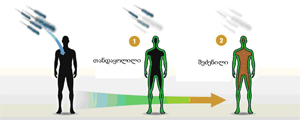
ნახ. 2: თანდაყოლილი და შეძენილი იმუნური სისტემების შედარება: 1 - თანდაყოლილი ხასიათდება სწრაფი რეაგირებით, იგი აჩერებს ინფექციას, და იმუნურ მეხსიერებას არ ტოვებს. 2 - შეძენილი - თანდაყოლილთან შედარებით ნელა მოქმედებს, საბოლოოს აძევებს ინფექციას ორგანიზმიდან, აქვს იმუნური მეხსიერების დატოვების უნარი.
ნახ. 3: თანდაყოლილი (1) და შეძენილი (2) იმუნიტეტის მოქმედების მექანიზმი: 1 - მიკროორგანიზმები ებმის Toll-რეცეპტორებს, რომლებიც უჯრედების ზედაპირზეა განლაგებული. ამის შედეგად აქტიურდება თანდაყოლილი იმუნიტეტი, რაც ანთებით პროცესებსა და მიკროორგანიზმების განადგურებას იწვევს. 2 - დენდრიტული უჯრედები ააქტიურებენ T-ლიმფოციტებს, რაც იწვევს შეძენილი იმუნიტეტის სისტემის ჩართვას. ამის შედეგად იწყება იმუნურ რეაქციათა რიგი, რომლის შედეგად წარმოიქმნება ანტისხეულები და უცხო სხეულებთან მებრძოლი „მკვლელი უჯრედები“.
ჟიულ ჰოფმანმა თავისი აღმოჩენა 1996 წელს გააკეთა, როდესაც მის ლაბორატორიაში ხილის ბუზების (დროზოფილების) ორგანიზმში ინფექციასთან ბრძოლის მექანიზმებს იკვლევდა. მისი ექსპერიმენტები ტარდებოდა ბუზებზე, რომლებიც შეიცავდნენ მუტაციებს Toll-გენში. რამდენიმე ხნით ადრე აღმოჩნდა, რომ ეს გენი დროზოფილის ჩანასახის ნორმალური განვითარებისათვის იყო საჭირო. მუტაციები, შესაბამისად, არღვევდნენ Toll-ის ნორმალურ ფუნქციას. როდესაც ჰოფმანმა ასეთი მუტანტების ორგანიზმი მიკროორგანიზმებით (ბაქტერიებით ან სოკოებით) დაასნებოვნა, აღმოჩნდა, რომ ბუზები სწრაფად იხოცებოდნენ. სიკვდილის მიზეზი კი ის იყო, რომ მუტანტებში, განვითარების ადრე აღმოჩენილი დეფექტის დამატებით, იმუნური დაცვის სისტემის ეფექტური ამოქმედება არ ხდებოდა. მაშასადამე, დაასკვნა ჰოფმანმა, Toll-გენი აუცილებელია პათოგენური მიკროორგანიზმების ამოსაცნობად, და იმუნური სისტემის გააქტიურება პათოგენების პასუხად ასევე Toll-გენის ნორმალურ მოქმედებაზე იყო დამოკიდებული.
ბრიუს ბიუტლერი თავის ექსპერიმენტებში ცდილობდა, ეპოვნა რეცეპტორი - მოლეკულა, რომელსაც ერთ-ერთ ბაქტერიულ პროდუქტთან (ლიპოპოლისაქარიდთან, LPS) შეეძლო ბმა. ამ პროდუქტის დაგროვება ადამიანის ორგანიზმში სეპტიკურ შოკს იწვევს. ეს მდგომარეობა სიცოცხლისათვის სახიფათოა, რადგან მისი განვითარებისას იმუნური სისტემის გადამეტებული სტიმულირებაც ხდება. 1998 წელს ბიუტლერმა დაადგინა, რომ თაგვებს, რომელნიც LPS-ისადმი მგრძნობიარენი არ იყვნენ, მუტაციებს სწორედ Toll-ის მსგავს გენში შეიცავდნენ. ეს Toll-ის მსგავსი რეცეპტორი (Toll-Like Receptor, TLR) ზუსტად LPS-ის რეცეპტორი აღმოჩნდა. როგორც კი LPS ამ რეცეპტორს ებმის, ანთების გამომწვევი სიგნალების წარმოქმნა ხდება. იმ შემთხვევაში კი, როცა LPS-ის რაოდენობა გადაჭარბებულია, სეპტიკური შოკის განვითარება ხდება. ამ ექსპერიმენტების საშუალებით დამტკიცდა, რომ პათოგენურ მიკროორგანიზმებთან ურთიერთობის შედეგად დროზოფილასა და ძუძუმწოვრებში წარმოქმნილი იმუნური პასუხი მეტად მსგავს მოლეკულებს ემყარება. ასე რომ, თანდაყოლილი იმუნიტეტის უნივერსალური სენსორები, როგორც იქნა, აღმოაჩინეს!
ჰოფმანისა და ბიუტლერის ექსპერიმენტებმა თანდაყოლილი იმუნიტეტის კვლევა მნიშვნელოვნად გააძლიერა. როგორც თაგვებში, ასევე ადამიანებში ათზე მეტი TLR-ის მსგავსი მოლეკულებია აღმოჩენილი. ყოველი მათგანი ამოიცნობს გარკვეული სახის მოლეკულებს, რომლებიც მიკროოგრანიზმებში გვხვდება. ადამიანები, რომელნიც TLR-ის მუტანტურ გენებს ატარებენ, სხვადასხვაგვარი ხიფათის წინაშე დგანან: ზოგიერთ შემთხვევაში, მიკროორგანიზმებით გამოწვეული ინფექციების რისკია მომატებული, სხვებს კი, იმუნური სისტემის „გადაჭარბებული მგრძნობიარობის“ გამო, ქრონიკული ანთებითი პროცესები უვითარდებათ.
უჯრედების ახალი ტიპი, რომელიც შეძენილ იმუნიტეტს აკონტროლებს. 1973 წელს რალფ სტაინმანმა აღმოაჩინა ახალი უჯრედი, რომელსაც დენდრიტული უჯრედი უწოდა. მან ივარაუდა, რომ დენდრიტული უჯრედები მნიშვნელოვანი იქნებოდა იმუნური სისტემის მოქმედებისათვის, და შეეცადა გამოერკვია, ძალუძთ თუ არა მათ T-უჯრედების გააქტიურება (გავიხსენოთ, რომ T-უჯრედები გადამწყვეტ როლს ასრულებენ იმუნური პასუხის განვითარებაში, და უზრუნველყოფენ იმუნური მეხსიერების წარმოქმნას მრავალი უცხო სხეულისა თუ ნაერთის წინააღმდეგ). სტაინმანმა დაადგინა, რომ დენდრიტული უჯრედები იწვევენ T-უჯრედების მნიშვნელოვან გააქტიურებას, ანუ ორგანიზმის იმუნურ პასუხს უცხო სხეულთა წინააღმდეგ. სტაინმანის კოლეგებმა პირველად ეს შედეგი უნდობლად მიიღეს, მაგრამ დამატებითმა ექსპერიმენტებმა უკვე ერთმნიშვნელოვნად აჩვენა, რომ დენდრიტულ უჯრედებს აქვთ T-უჯრედების გააქტივების უნიკალური თვისება.
სტაინმანისა და მისი კოლეგების შემდგომმა კვლევებმა ისიც ცხადყო, რა მექანიზმების მეშვეობით იღებს „გადაწყვეტილებას“ შეძენილი (ადაპტური) იმუნური სისტემა, საჭიროა თუ არა გააქტივება გარკვეული ნივთიერებისა თუ სხეულის ამოცნობის შედეგად. ასევე აღმოჩნდა, რომ T-უჯრედების მოქმედების კონტროლი ხორციელდება სიგნალებით, რომლებსაც შეძენილი იმუნური სისტემა წარმოქმნის და რომლებსაც დენდრიტული უჯრედები იღებენ. ამ მექანიზმების წყალობით, იმუნური სისტემა რეაგირებს სხეულში შემოჭრილ პათოგენებზე, მაგრამ არ უტევს საკუთარი ორგანიზმის მიერ წარმოქმნის მრავალფეროვან მოლეკულებს.
ფუნდამენტური კვლევიდან სამედიცინო გამოყენებამდე. აღმოჩენებმა, რომლებსაც 2011 წლის ნობელის პრემია მიენიჭა, მოგვცა ახალი ცოდნა იმის შესახებ, თუ როგორ ხდება ჩვენი იმუნური სისტემის აქტივირება და მისი მოქმედების რეგულაცია. ამ კვლევების საფუძველზე შესაძლებელი გახდა სხვადასხვა ავადმყოფობების როგორც თავიდან აცილება, ასევე განკურნებაც. შეიქმნა ახალი, გაუმჯობესებული ვაქცინები, მიმდინარეობს ექსპერიმენტები იმის გამოსარკვევად, შეიძლება თუ არა ავთვისებიანი სიმსივნის წინააღმდეგ ბრძოლა ორგანიზმის იმუნური სისტემის სტიმულირების გზით. ასევე გაირკვა მექანიზმები, რომლების დარღვევა იწვევს აუტოიმუნურ და ანთებით დაავადებებს, როცა იმუნური სისტემა საკუთარი ორგანიზმის მიერ წარმოქმნილი ნაერთების წინააღმდეგ იწყებს მოქმედებას.
სტატიის დასასრულს გთავაზობთ მოკლე ინფორმაციას იმუნური სისტემის კომპონენტების კვლევის მთავარი ეტაპებისა და იმ მკვლევარების შესახებ, რომელთა დამსახურება ნობელის პრემიით იყო აღნიშნული:
1901 წელი - ემილ ფონ ბერინგი; პასიურ-აგრესიული სისტემა. ფონ ბერინგმა აღმოაჩინა ფაქტორები, რომლებიც სისხლის შემადგენლობაში შედიან და რომლებსაც ძალუძთ გაშეშებისა ან დიფთერიის გამომწვევი ტოქსიკური ნაერთების გაუვნებლება. მან ასევე გაარკვია, როგორც შეიძლება ამ ფაქტორების გამოყენება დიფთერიის თავიდან ასაცილებლად.
1908 - ილია მეჩნიკოვი და პაულ ერლიხი; დაცვის მრავალი მექანიზმი. დამტკიცდა, რომ იმუნურ სისტემას მოქმედების რამდენიმე მექანიზმი აქვს; მეჩნიკოვმა აღმოაჩინა ფაგოციტური უჯრედები, რომლებიც გარს ერტყმიან მომხდურებს და ანადგურებენ მათ; ერლიხის თეორია იხილავდა, თუ როგორ ხდება ანტისხეულების გამოყოფა სისხლში და მათ მიერ უცხო სხეულებთან ურთიერთქმედება.
1913 - შარლ რიშე; შოკური პასუხი. რიშემ აღმოაჩინა ანაფილაქსია, ალერგიული რეაქცია ტოქსინების მიმართ. ამ აღმოჩენის საფუძველზე გაირკვა, რომ იმუნურ სისტემას, ავადმყოფობისაგან დაცვის გარდა, საკუთარი ორგანიზმის დაზიანებაც ძალუძს.
1919 - ჟიულ ბორდე; კომპლემენტური ძალები. სისხლის შრატში არსებულ ფაქტორებს შეუძლიათ, ანტისხეულებთან ერთად იმოქმედონ ბაქტერიების გასანადგურებლად. ბორდემ აღმოაჩინა ეს ფაქტორები, რომელთაც კომპლემენტური ცილები უწოდა, და რომელთა საფუძველზე შემუშავდა მრავალი დიაგნოსტიკური ტესტი სახიფათო ინფექციურ დაავადებებზე.
1930 - კარლ ლანდსტაინერი; სისხლის ჯგუფები. ლანდსტაინერმა აღმოაჩინა ადამიანის სისხლის ჯგუფები, რამაც შესაძლებელი გახადა სისხლის გადასხმა ორგანიზმის მძაფრი რეაქციის გარეშე, რომელიც სისხლის შეუთავსებლობით იქნებოდა გამოწვეული.
1960 - ფრანკ მაკფარლაინ ბარნეტი, პეტერ მედავარი; იმუნოლოგიური შემგუებლობა. ამ კონცეფციის საფუძველზე ნათელი გახდა, როგორ ახერხებს იმუნური სისტემა, არ იმოქმედოს საკუთარი ორგანიზმის კომპონენტების წინააღმდეგ და, მაშასადამე, არ გამოიწვიოს თავისივე განადგურება.
1972 - ჯერალდ ედელმანი, როდნი პორტერი; ანტისხეულების სტრუქტურა. ამ მეცნიერებმა ერთმანეთისაგან დამოუკიდებლად გაშიფრეს ანტისხეულების სტრუქტურა, რის შედეგადაც გაირკვა, თუ როგორც ხდება სხვა ანტისხეულებთან გარეგნულად მსგავსი ერთი ანტისხეულის მიერ მრავალი სხვადასხვა უცხო სხეულიდან (ანტიგენიდან) ერთ-ერთის სპეციფიურად ამოცნობა და განადგურება.
1980 - ბარუხ ბენაცერრაფი, ჟან დოსეტი და ჯორჯ სნელი; თავსებადობის მექანიზმები. ამ სამი მეცნიერის მიერ უჯრედის ზედაპირზე აღმოჩენილია ცილები, რომლებიც იმუნურ პასუხს არეგულირებენ. მათ შრომებში ის მექანიზმიცაა აღწერილი, თუ როგორ ხდება ამ ცილების მიერ იმუნური პასუხის რეგულირება.
1984 - ნილს ერნე, ჯორჯ კოლერი, ცეზარ მილშტაინი; სპეციფიური ანტისხეულების წარმოქმნა. ერნეს თეორიებში ჩამოყალიბდა იმუნური სისტემის მიერ ანტისხეულების მართვის პრინციპი. კოლერისა და მილშტაინის მიერ შექმნილმა სპეციფიური ანტისხეულების წარმოების მეთოდმა ხელი შეუწყო უკეთესი დიაგნოსტური ტესტების შექმნას.
1987 - სუსუმუ ტონეგავა; ანტისხეულების წარმოქმნის გენეტიკური მექანიზმები. ტონეგავამ შეისწავლა ანტისხეულების წარმოქმნის გენეტიკური მექანიზმები, რის შედეგადაც გაირკვა, თუ როგორაა შესაძლებელი მილიონობით სპეციფიური ანტისხეულის წარმოქმნა ბევრად უფრო ნაკლები რაოდენობის გენისაგან.
1996 - პიტერ დოერთი, როლფ ზინკერნაგელი; ვირუსების ამოცნობა იმუნური სისტემის მიერ. ვირუსით დაავადებული უჯრედების კვლევამ გამოააშკარავა, თუ როგორ ამოიცნობიან უცხო სხეულები იმუნური სისტემის მიერ, და როგორ განასხვავებს იმუნური სისტემა საკუთარსა და უცხო ნაერთებს.
გელა თევზაძე.
24 საათი 06.11.11
|
|
21 მიტოქონდრიული დნმ: არაქრომოსომული გენეტიკური ინფორმაცია, რომელიც ჩვენი წარმომავლობის ისტორიას შეიცავს. |
▲ზევით დაბრუნება |
ცნობილია, რომ ყოველი ცოცხალი ორგანიზმი უჯრედებისაგან შედგება. ყველა მრავალუჯრედიანი (და ზოგიერთი ერთუჯრედიანი) ორგანიზმის უჯრედები შეიცავს ბირთვსა და მის გარშემო არსებულ ციტოპლაზმას. ორგანიზმების გენეტიკური ინფორმაცია დეზოქსირიბონუკლეინის მჟავაში (დნ მჟავაში, ან დნმ-ში) არის კოდირებული, მისი უდიდესი ნაწილი უჯრედების ბირთვის შიგნით არის მოთავსებული და ქრომოსომებშია განლაგებული. მნიშვნელოვანი გამონაკლისია მიტოქონდრიული დნ მჟავა, რომელიც განლაგებულია ბირთვის გარეთ, ციტოპლაზმაში, უფრო ზუსტად კი მიტოქონდრიებში. მიტოქონდრიები ციტოპლაზმაში არსებული უჯრედსშიდა წარმონაქმნებია, რომელთა მთავარი ფუნქცია უჯრედისათვის აუცილებელი ენერგიის წარმოქმნა და დაგროვებაა.
რადგან მიტოქონდრიები - და შესაბამისად, მათში მოთავსებული დნმ - ბირთვის გარეთ არსებობს, მათი მემკვიდრეობით გადაცემის მექანიზმი მნიშვნელოვნად განსხვავდება ბირთვული დნმ-ის გადაცემის მექანიზმისაგან. როგორც ცნობილია, ყველა ორგანიზმი ბირთვული გენეტიკური მასალის ორ ტოლ ნახევარს იღებს: ერთს - დედისაგან, მეორეს კი - მამისაგან. ამიტომ, მაგალითად, ყველა ადამიანის 46 ქრომოსომიდან 23 დედისგანაა მიღებული, 23 კი - მამისაგან. ეს მოვლენა იმაზეა დაფუძნებული, რომ სასქესო უჯრედის წარმოქმნისას მასში ორგანიზმის ბირთვული გენეტიკური მასალის (ბირთვული დნმ-ის) რაოდენობის ნახევარი გადადის. განაყოფიერებისას, ანუ მდედრობითი და მამრობითი სასქესო უჯრედების შერწყმისას, ეს ორი ნახევარი (ადამიანის შემთხვევაში - 23-23 ქრომოსომა) ერთიანდება და იწყება ახალი ორგანიზმის განვითარება.
ნახ. 1: ბირთვული (მარცხნივ) და მიტოქონდრიული (მარჯვნივ) დნმ-ის მემკვიდრეობით გადაცემის მექანიზმების შედარება.
მიტოქონდრიული დნმ-ის შემთხვევაში მემკვიდრეობის მექანიზმი მნიშვნელოვნად განსხვავებულია. ეს განსხვავება იმითაა განპირობებული, რომ, თუმცა მდედრობითი და მამრობითი სასქესო უჯრედები ბირთვული დნმ-ის შემცველობის მხრივ ტოლფასნი არიან, ისინი მკვეთრად განირჩევიან ციტოპლაზმის (და, მაშასადამე, ციტოპლაზმაში არსებული მიტოქონდრიული დნმ-ის) შემცველობით: კვერცხუჯრედი (მდედრობითი სასქესო უჯრედი) ციტოპლაზმას დიდი რაოდენობით შეიცავს, სპერმატოზოიდი (მამრობითი სასქესო უჯრედი) კი ციტოპლაზმას თითქმის მთლიანადაა მოკლებული. შესაბამისად, ყოველ ორგანიზმში არსებული მიტოქონდრიული დნმ სწორედ დედის მხრიდანაა მიღებული.
ეს გარემოება განსაზღვრავს მიტოქონდრიული (და ზოგადად, ციტოპლაზმური) დნმ-ის მემკვიდრეობით გადაცემის თავისებურებას: მიტოქონდრიულ დნმ-ში კოდირებული ინფორმაცია დედისაგან გადაეცემა როგორც ბიჭებს, ასევე გოგონებს, მაგრამ ამ ინფორმაციის გადაცემა შემდეგი თაობისათვის მხოლოდ ქალებს შეუძლიათ. გავრცელებული მეტაფორით, ყველა ადამიანი თავის უჯრედებში „ევას მიტოქონდრიულ დნმ-ს“ ატარებს. იგივე მეტაფორას თუ განვავრცობთ, ადამის მიტოქონდრიული დნმ პირველივე თაობაში „დაიკარგა“.
ექსპერიმენტულ მეცნიერებაში კი ადამიანის ევოლუციის, მისი წარმომავლობისა და დედამიწაზე განსახლების ქრონოლოგიის შესწავლისას, მიტოქონდრიული დნმ-ის ანალიზი უკვე მიღებული და აღიარებული მეთოდია. თუ ბირთვული დნმ ყოველ თაობაში განიცდის ცვლილებას (ქრომოსომების ფრაგმენტების მიმოცვლის, ახალი ორგანიზმის წარმოქმნისას ახლებური კომბინაციების წარმოქმნის გამო), მიტოქონდრიული დნმ უცვლელად გადაეცემა შვილებს დედისაგან, და ქალიშვილებისაგან - შვილიშვილებს. მემკვიდრეობითი ცვლილებები მიტოქონდრიულ დნმ-ში მეტად იშვიათია, რადგან, ბირთვული დნმ-ისაგან განსხვავებით, იგი არც ფრაგმენტებს მიმოიცვლის და არც ორად იყოფა სქესობრივი უჯრედების წარმოქმნისას, რომ შემდეგ ახალ „ნახევარს“ შეერწყას. ის ცვლილებები კი, რომელიც მიტოქონდრიულ დნმ-ში მაინც ხდება, ძალიან ინფორმატულია და ადამიანთა დიდი ჯგუფების წარმომავლობის ერთგვაროვნებისა თუ განსხვავებულობის მიმანიშნებელია.
მიტოქონდრიული დნმ-ის გამოყენება ადამიანის წინაპრებისა და მისი წარმომავლობის შესასწავლად 1987 წელს დაიწყო, როდესაც რებეკა კანმა თანამშრომლებთან ერთად გამოაქვეყნდა სტატია „მიტოქონდრიული დნმ და ადამიანის ევოლუცია“ (Rebecca Cann, „Mitochondrial DNA and Human Evolution“). ამ სტატიაში გადმოცემული იყო 147 ადამიანის მიტოქონდრიული დნმ-ის ანალიზი. ამ ადამიანების წარმოშობა (უფრო ზუსტად, მათი მდედრობითი სქესის წინაპრების წარმომავლობა) ხუთ სხვადასხვა გეოგრაფიულ რეგიონს მიეკუთვნებოდა. სტატიის ორი ძირითადი დებულება, რამდენიმე მცირე უზუსტობის მიუხედავად, ახლაც ჭეშმარიტადაა მიჩნეული: 1) „მიტოქონდრიული დნმ-ის ხეს“ ფესვები აფრიკაში აქვს გადგმული, ანუ ხუთივე რეგიონის წარმომადგენლების წარმომავლობა აფრიკაში იწყებოდა; 2) ამ ხის ყველა „ტოტი“ შედარებით მოკლეა, რაც მიგვანიშნებს, რომ ხუთივე ჯგუფის საერთო წინაპარი შედარებით მცირე ხნის წინ ცხოვრობდა. ცხადია, რომ ამ საერთო წინაპრის დასახელებისათვის „ევა“ ყველაზე შესაფერი აღმოჩნდა...
ამ დებულებების საფუძველზე, ავტორებმა დაასკვნეს, რომ მათი შედეგები კარგად მიესადაგებოდა ადამიანის ევოლუციის ერთ-ერთ მოდელს. ამ მოდელის თანახმად, ანატომიურად თანამედროვე ადამიანის წარმოშობა პირველად აფრიკაში მოხდა თითქმის 150 ათასი წლის წინათ; ამას მოჰყვა ადამიანების განსახლება აფრიკიდან - დაახლოებით 100 ათასი წლის წინ. ამ განსახლების შედეგად, აფრიკიდან გამოსულმა ადამიანებმა სრულიად შეცვალეს (განდევნეს) ყველა სხვა ჯგუფი, რის შედეგადაც ყველა თანამედროვე ადამიანი ამ - აფრიკაში წარმოშობილი და შემდგომ განსახლებული - პოპულაციის (ანუ ინდივიდთა ჯგუფის) შთამომავალია. ეს ჰიპოთეზა, მიტოქონდრიული დნმ-ის კვლევის შედეგებისაგან დამოუკიდებლად, უკვე განიხილებოდა პალეოანთროპოლოგიურ ლიტერატურაში. მისი მთავარი ალტერნატივა იყო ე.წ. მულტირეგიონალური (მულტიცენტრული) ჰიპოთეზა, რომლის თანახმადაც როგორც აფრიკაში, ასევე ევრაზიაში მოსახლე „წინაპართა პოპულაციები“ მონაწილეობდნენ ანატომიურად თანამედროვე ადამიანის ფორმირებაში.

ნახ. 2: ადამიანის ევოლუციის „მულტირეგიონალური“ (მარცხნივ) და „აფრიკიდან ჩანაცვლების“ (მარჯვნივ) მოდელები.
1989 წელს უკვე გამოიცა პირველი სახელმძღვანელო ადამიანის ევოლუციაში, სადაც რებეკა კანის შედეგები იყო ნახსენები. სურათზე, რომელიც ამ სახელმძღვანელოდაა გადმობეჭდილი, ორივე ხსენებული ჰიპოთეზა გრაფიკულადაა მოცემული. როგორც ილუსტრაციიდან ჩანს, ორივე ჰიპოთეზის მიხედვით ადამიანის წარმოშობა აფრიკაში იღებს სათავეს, განსახლება კი 1.7-1.9 მილიონი წლის წინ დაიწყო. ადამიანის წინაპრებს, რომლებიც იმ დროში ცხოვრობდნენ, Homo erectus-ს („გამართულ ადამიანს“) უწოდებენ. პირველი (მულტირეგიონალური) მოდელის თანახმად, აფრიკიდან Homo erectus-ის სამი განსხვავებული პოპულაცია გამოვიდა და მათგან ადამიანის თანამედროვე აფრიკელები, აზიელები და ევროპელები ჩამოყალიბდნენ. მეორე მოდელის ვარაუდით, პირველი განსახლება მართლაც 1.7-1.9 მილიონი წლის წინ შეიძლება მომხდარიყო, მაგრამ ბევრად გვიან (დაახლოებით 100 ათასი წლის წინ) აფრიკიდან კიდევ ერთი ფორმა გამოვიდა, რომელმაც ევროპასა თუ აზიაში ყველა დანარჩენი პოპულაცია შეცვალა (გამოდევნა). ეს პროცესი მეორე მოდელის ამსახველ ნახაზზე (B) წყვეტილი მონაკვეთითაა აღნიშნული. ამ მოდელს, პირველისაგან ძირითადი განსხვავების აღსანიშნავად, „აფრიკიდან ჩანაცვლების“ (Out-of-Africa Replacement) მოდელს უწოდებენ.
ნახ. 3: ადამიანების განსახლების შესაძლო სქემა „აფრიკიდან ჩანაცვლების“ მოდელის თანახმად.
რებეკა კანის ნაშრომში აღნიშნულია, რომ მათი შედეგების საფუძველზე აგებული „მიტოქონდრიული ხის“ „ტოტები“ არააფრიკულ (ევროპულ, აზიურ) პოპულაციებს შორის მკვეთრად განსხვავებული წინაპრების არსებობას არ მიანიშნებდნენ. ასეთი წინაპრები კი, პირველი მოდელის თანახმად, ნამდვილად უნდა არსებობდნენ! ამგვარად, გენეტიკური კვლევის შედეგებმა უარჰყო მულტირეგიონალური ჰიპოთეზა, „აფრიკიდან ჩანაცვლების“ ჰიპოთეზას კი მხარს უჭერდა. სულ ცოტა ხანში, სასკოლო სახელმძღვანელოებისა და სამეცნიერო-პოპულარული ლიტერატურის თანახმად, „აფრიკიდან ჩანაცვლება“ ადამიანის ევოლუციის ერთადერთ ჭეშმარიტ მოდელად იქნა მიჩნეული.
მაგრამ აუცილებელია აღინიშნოს, რომ რებეკა კანის სტატიაში რამდენიმე უმართებულო მსჯელობა არსებობს: 1) მის მიერ აღწერილი პირველი მოდელი სრულყოფილად ვერ აკმაყოფილებს „მულტირეგიონალური“ ჰიპოთეზის განსაზღვრებას (დაწვრილებით იხ. ქვემოთ); 2) კანსა და მისი თანაავტორებს თავისი ჰიპოთეზა ექსპერიმენტულად (მაგ., სტატისტიკური ანალიზის გამოყენებით) არ შეუმოწმებიათ; 3) არსებობს ადამიანის ევოლუციის არა მხოლოდ ორი, არამედ რამდენიმე მოდელი, ასე რომ ორიდან ერთის უარყოფა სულაც არ ნიშნავს, რომ მეორე მოდელის ჭეშმარიტება ეჭვსგარეშეა. 4) ზოგადი პროცესის სტატისტიკური დასაბუთება არ უზრუნველჰყოფს ამ პროცესის შემადგენელი ნაწილების ამსახველი მოდელების ჭეშმარიტების დამტკიცებას. ახლა ეს ოთხი სუსტი წერტილი ცალკ-ცალკე განვიხილოთ:
1) რა არის მულტირეგიონალური ჰიპოთეზა? ადამიანის ევოლუციის მულტირეგიონალური მოდელი უშვებს, რომ შესაძლებელია ევოლუციის გასაოცარი პარალელიზმი ადამიანის წინაპრებიდან მის - სულ ცოტა, სამ - თანამედროვე ფორმებამდე. ეს მოდელი ჯერ კიდევ კანის ნაშრომის გამოქვეყნებამდე იყო დისკრედიტირებული, თუმდაც იმის საფუძველზე, რომ ასეთი „სამგზის პარალელური ევოლუცია“ თეორიულად ძნელი წარმოსადგენი იყო. მეტიც, კვლევებმა ცხადჰყო, რომ ევროპელ, აზიელ და აფრიკელ პოპულაციებს შორის არსებული გენეტიკური განსხვავებები მცირე იყო და არ იძლეოდა იმის საფუძველს, რომ ძველი, ერთმანეთისაგან მნიშვნელოვნად დაშორებული და დამოუკიდებელი წინაპრების არსებობა დაგვეჯერებინა. ამიტომ, კანის სტატიაში განხილული მოდელი იმ დროისათვისაც კი უკვე უარყოფილი იყო. მაგრამ უკვე არსებობდა სხვაგვარი მულტირეგიონალური მოდელი, რომლის თანახმადაც ადამიანთა სხვადასხვა პოპულაციები (ავსტრალიური, მონღოლური, სამხრეთაფრიკული, ევრაზიული) ევოლუციის დროს აქტიურად მიმოიცვლიდნენ გენებს (ე.წ. „გენების დრეიფის“ მექანიზმით). სხვაგვარად რომ ვთქვათ, ხდებოდა არა „სამგზის“ (ან „ოთხგზის“!) პარალელური ევოლუცია, არამედ ურთიერთდაკავშირებული განვითარება. თითოეული პოპულაცია, რასაკვირველია, დანარჩენებისაგან მეტ-ნაკლებად განსხვავდება, მაგრამ ერთმანეთისაგან სრულიად დამოუკიდებელ ევოლუციას ეს მოდელი არც კი ახსენებს. მეტიც, ამ მოდელში ოთხი „ევოლუციური ხაზი“ მუდმივ ურთიერთქმედებაში იმყოფება და ცალკეულ განშტოებებს არ წარმოქმნის. და ბოლოს, „მულტირეგიონალური მოდელის“ არა ერთი, არამედ რამდენიმე ვერსია არსებობს, რომლებიც განსხვავდებიან ევოლუციის პროცესში გენების დრეიფისა და გადარჩევის პირობების როლების სხვადასხვაგვარი განმარტებით. „აფრიკიდან განსახლება-გამოცვლის“ ჰიპოთეზა კი „მულტირეგიონალური მოდელებთან“ შედარებით უფრო მარტივად ხსნის ადამიანის ევოლუციაში მიმდინარე პროცესებს. სწორედ ამ მოსაზრებითაა აღიარებული „აფრიკიდან ჩანაცვლების“ მოდელი ადამიანის ევოლუციის აღწერისათვის ყველაზე უფრო შესაფერის საწყის ჰიპოთეზად.
ნახ. 4: ადამიანის წარმოშობის „მულტირეგიონალური“ მოდელი.
2. ჰიპოთეზასთან თავსებადობა თუ ჰიპოთეზის შემოწმება? ყველა სტუდენტისათვის მეტად მნიშვნელოვანია იმის გაცნობიერება, რომ მეცნიერების ერთ-ერთი უმთავრესი მიზანია ექსპერიმენტული შედეგების დაგროვება რაიმე ჰიპოთეზის შესამოწმებლად. სამწუხაროდ, მრავალ შემთხვევაში ადამიანის წარმომავლობასთან დაკავშირებით ჩატარებულ გენეტიკურ ექსპერიმენტებში მკვლევარები თავს არიდებენ ჰიპოთეზის შემოწმებას და მხოლოდ იმას აჩვენებენ მკითხველს, რომ მიღებული შედეგები თავსებადია რომელიმე (ხშირად ადრევე „შერჩეულ“) ჰიპოთეზასთან. როგორც აღვნიშნეთ, საწყისი შედეგები მიტოქონდრიული დნმ-ის გამოყენებით თავსებადი იყო როგორც „გამოცვლის“ მოდელთან, ასევე „მულტირეგიონალური მოდელის“ ახალ ვერსიასთან. ძირითადი სხვაობა ამ მოდელებს შორის იმაში მდგომარეობდა, მართლაც მოხდა თუ არა ევრაზიული პოპულაციების „ჩანაცვლება“. სხვა მხრივ - აფრიკული „ფესვებისა“ თუ შედარებით მცირე ხნის წინ არსებული საერთო წინაპრის თვალსაზრისით - ეს ორი მოდელი ერთმანეთისაგან თითქმის არ განსხვავდებოდა. ჩანაცვლების დასაბუთებისათვის კი აუცილებელია, რომ მიღებული შედეგები საკმარის ინფორმაციას გვაძლევდნენ პოპულაციების შესახებ როგორც ჩანაცვლების პერიოდში და მის შემდეგ, ასევე იმ დრომდეც, როცა ნავარაუდევი ჩანაცვლება მოხდა. მხოლოდ ამ შემთხვევაში შესაძლებელია საუბარი იმ ჰიპოთეზის შემოწმებაზე, რომლის თანახმადაც რაიმე ჩანაცვლება მართლაც მოხდა. მაშასადამე, ჭეშმარიტად ფასეულ ინფორმაციას გენეტიკური კვლევის ის შედეგები შეიცავენ, რომელთა საფუძველზე შესაძლოა ცვალებადობის ხარისხის დადგენა ჩანაცვლებამდე, ჩანაცვლებისას და ჩანაცვლების შემდეგ, და ამ მონაცემების შედარება. კანის კვლევაში კი, სამწუხაროდ, ასეთი შედეგები არ იყო მიღებული...
3. ბირთვული გენების „ხის ფესვები“. კანის ნაშრომში „ჩანაცვლების“ ჰიპოთეზის ერთ-ერთ უმთავრეს მხარდამჭერ საბუთად მიტოქონდრიული დნმ-ის „ხის“ „აფრიკული ფესვები“ ითვლება. მაგრამ „აფრიკული ფესვები“ ხომ მულტირეგიონალურ მოდელთანაცაა თავსებადი? თუ ამ უკანასკნელი მოდელის განხილვისას დავუშვებთ, რომ ადამიანთა წინაპრების ძირითადი პოპულაცია აფრიკაში ~2-1.5 მილიონი წლის წინაც ცხოვრობდა, „აფრიკული ფესვები“ აქაც სავსებით ბუნებრივი მოვლენაა...
რამდენიმე ხნის შემდეგ ჩატარდა ანალოგიური კვლევა Y-ქრომოსომაზე. ეს ქრომოსომა მხოლოდ მამაკაცებს აქვთ და, შესაბამისად, მამებისაგან მხოლოდ მამრობითი სქესის შვილებს გადაეცემათ. მიუხედავად იმისა, რომ Y-ქრომოსომა ბირთვში მდებარეობს, იგი, სხვა ქრომოსომებისაგან განსხვავებით, გამრავლებისა და ახალი ორგანიზმების წარმოქმნის დროს არ მიმოიცვლის ფრაგმენტებს და თითქმის უცვლელი ხდება თაობათა განმავლობაში. სხვაგვარად რომ ვთქვათ და ამ სტატიის დასაწყისში გამოყენებული მეტაფორა მოვიშველიოთ, ყველა მამაკაცს „ადამის Y-ქრომოსომა“ აქვს. ამიტომაც Y-ქრომოსომის ანალიზი მიტოქონდრიული დნმ-ის ტოლფასად შეიძლება ჩაითვალოს.
Y-ქრომოსომის ანალიზით მიღებული შედეგები ასევე თავსებადი იყო „აფრიკული ფესვების“ არსებობის იდეასთან. უნდა აღინიშნოს, რომ ამ შემთხვევაშიც სტატიის ავტორებმა სიფრთხილე გამოიჩინეს და ამ შედეგით დაკმაყოფილდნენ, ანუ თავსებადობა აღნიშნეს, მაგრამ ჰიპოთეზის სტატისტიკური შემოწმება აღარ დაუწყიათ.
სხვადასხვა ბირთვული გენების ანალიზის საფუძველზე აგებული „ხეები“ (მიტოქონდრიული დნმ-ის „ხის“ მსგავსი) მიანიშნებენ, რომ საერთო წინაპარი (ხის „ფესვები“) 100 ათას წელზე უფრო დიდი ხნის წინ არსებობდა. სხვაგვარად რომ ვთქვათ, „ფესვების“ ასაკი უფრო მეტია, ვიდრე „ჩანაცვლების“ ხანა. მაგალითად, აღმოჩენილია ჰემოგლობინის გენები, რომელთაც აშკარად „აზიური წარმოშობა“ აქვთ, მათი ასაკი კი 200 ათას წელს აღემატება. არის ისეთი გენიც, რომელიც ასევე „აზიური წარმოშობისაა“, მაგრამ მისი „ფესვების“ ასაკი 2 მილიონ წელს უახლოვდება. ამრიგად, არსებობს მონაცემები, რომლებიც „ჩანაცვლების“ ჰიპოთეზით (ყოველ შემთხვევაში, მისი თანამედროვე ფორმულირებით) უბრალოდ ვერ აიხსნება.
ცნობილია სტატისტიკური მეთოდები, რომელთა მეშვეობითაც შეიძლება ექსპერიმენტული მონაცემების შეფასება მათი თავსებადობის თვალსაზრისით ორ ალტერნატიულ მოდელთან. მაგალითად, ჩანაცვლების მოდელის თანახმად, 100 ათას წელზე უფრო „ხნიერი“ გენის „ფესვების“ აფრიკული წარმოშობის ალბათობა 1-ია (ანუ 100%25). მულტირეგიონალური მოდელის მიხედვით კი, თუმცა აფრიკული წარმოშობა ყველაზე უფრო სავარაუდოა, ასეთი ალბათობა მხოლოდ 0.5-ია (ანუ 50%25). სამწუხაროდ, ამ შემთხვევაში ვერც მიტოქონდრიულ დნმ-ისა და ვერც Y-ქრომოსომის ანალიზი დამაჯერებელ პასუხს ვერ იძლევა, რადგან ორივე მათგანი მხოლოდ „ერთი გენეტიკური ერთეულია“ (წრიული მიტოქონდრიული დნ მჟავის ერთი მოლეკულა, ერთი წრფივი Y-ქრომოსომა), და სტატისტიკისათვის სანდო მასალას ვერ მოგვცემს.
ადამიანის წარმომავლობის მოდელების განხილვის გაგრძელებასა და ამ მიმართულებით კვლევის განვითარების ანალიზს შემდეგ სტატიაში შემოგთავაზებთ.
გელა თევზაძე.
24 საათი 20.11.11